Corteza prefrontal
La corteza prefrontal o córtex prefrontal es la parte anterior de los lóbulos frontales del cerebro, y se ubica por delante de las áreas motora y premotora. Se distingue de otras áreas del lóbulo frontal por su composición celular, su inervación dopaminérgica y sus aferencias talámicas.[1] Esta región cerebral está involucrada en la planificación de comportamientos cognitivamente complejos, en la expresión de la personalidad, en los procesos de toma de decisiones y en la adecuación del comportamiento social adecuado en cada momento.[2] Se considera que la actividad fundamental de esta región cerebral es la coordinación de pensamientos y acciones de acuerdo con metas internas.[3]
| Corteza prefrontal | ||
|---|---|---|
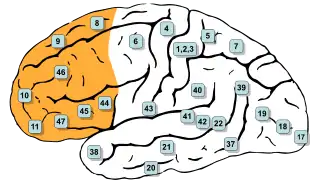 Áreas de Brodmann de la superficie lateral del cerebro. Las áreas #8, #9, #10, #11, #44, #45, #46, y #47 se encuentran en la región prefrontal. | ||
| Es parte de | Lóbulo frontal | |
| Componentes |
Circunvolución frontal superior Circunvolución frontal media Circunvolución frontal inferior | |
| Arterias |
Cerebral anterior Cerebral media | |
| Venas | Seno sagital superior | |
| Sinónimos | ||
| Enlaces externos | ||
| NeuroNames | ancil | |
El término médico más utilizado para referirse a las funciones desempeñadas por la corteza prefrontal es «función ejecutiva». Este término hace referencia a la capacidad para establecer distinciones entre pensamientos conflictivos, realizar juicios acerca del bien y del mal, predecir las consecuencias futuras de actividades actuales, trabajar conforme a metas determinadas de antemano, realizar predicciones de resultados, creación de expectativas, y control social (la capacidad para inhibir comportamientos impulsivos que, de no ser suprimidos, podrían desembocar en resultados socialmente inaceptables).
Muchos autores han señalado la existencia de una relación entre la corteza prefrontal y las características de la personalidad de un individuo.[4]
Definición
Existen tres formas posibles de definir la corteza prefrontal:
- Como la corteza frontal granular.
- Como la zona de proyección del núcleo mediodorsal del tálamo.
- Como la parte de la corteza frontal cuya estimulación eléctrica no provoca movimientos.
La corteza prefrontal ha sido definida con base en la citoarquitectura cerebral por la presencia de una capa granular (la capa cortical IV). No está completamente claro quién fue el primero en utilizar este criterio. Muchos de los investigadores pioneros en citoarquitectura de la corteza cerebral restringieron el término «prefrontal» a una región mucho más pequeña de la corteza que incluía la circunvolución recta y la circunvolución rostral (Alfred Walter Campbell, 1905; Grafton Elliot Smith, 1907; Korbinian Brodmann, 1909; Constantin von Economo y Georg N. Koskinas, 1925). No obstante, en 1935, Carlyle F. Jacobsen utilizó el término «prefrontal» para diferenciar las áreas prefrontales granulares de las áreas no granulares motora y premotora.[5]
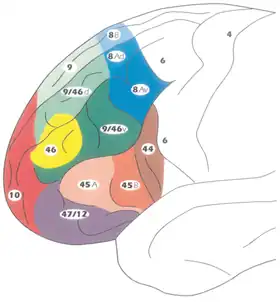
En la terminología de áreas de Brodmann, la corteza prefrontal incluye tradicionalmente las áreas 8, 9, 10, 11, 44, 45, 46, y 47 (no obstante, no todas estas áreas son estrictamente granulares, como las áreas 44, 11 y 47).[6] El principal problema que afecta a esta definición es que sólo es válida en primates, ya que los no primates no disponen de una capa IV granular.[7]
La definición de la corteza prefrontal como la zona de proyección de los núcleos mediodorsales del tálamo se basa en el trabajo de Rose y Woolsey,[8] que demostraron que estos núcleos se proyectan hacia las zonas anterior y ventral del cerebro en animales no primates. No obstante, Rose y Woolsey denominaron a esta zona de proyección como zona orbitofrontal. Parece que fue Akert quien, en 1964, sugirió por primera vez explícitamente que este criterio podría ser utilizado para definir las zonas homólogas a la corteza prefrontal en primates y no primates.[9] Esto permite el establecimiento de homologías, a pesar de la carencia de la capa granular en los animales no primates. Esta definición de corteza prefrontal como zona de proyección sigue estando ampliamente aceptada en la actualidad,[10] aunque su utilidad ha sido puesta en tela de juicio.[6][11] Existen estudios actuales que han mostrado que las proyecciones del núcleo mediodorsal del tálamo no están restringidas a la corteza frontal granular en los primates. Por ello, se ha propuesto una definición de la corteza prefrontal como la región de la corteza cerebral que mantiene unas conexiones recíprocas con los núcleos mediodorsales más fuertes que con cualquier otro núcleo talámico.[7] No obstante, Uylings et al.[7] reconocen que incluso con la aplicación de este criterio, resulta difícil definir inequívocamente la corteza prefrontal.
Una tercera definición de la corteza prefrontal es como aquella área de la corteza frontal cuya estimulación eléctrica no provoca movimientos observables. Por ejemplo, en el año 1890, David Ferrier[12] utilizó el término en este sentido. Un problema con esta definición es que existen zonas cuya estimulación no produce movimientos tanto en las áreas granulares como en las no granulares de la corteza frontal.[6]
Origen del término
El término prefrontal como una descripción de una parte del cerebro parece haber sido introducido por Richard Owen en 1868.[5] Para este autor, el área prefrontal estaba limitada a la parte más anterior del lóbulo frontal (aproximadamente, la zona correspondiente al polo frontal). Se ha hipotetizado que la elección de este término se basó en el hueso prefrontal, presente en la mayoría de los anfibios y reptiles.[5]
Interconexiones
La corteza prefrontal está fuertemente interconectada con gran parte del cerebro. Se encuentran abundantes conexiones con otras regiones corticales y subcorticales. La corteza prefrontal dorsal está especialmente interconectada con regiones cerebrales implicadas en procesos como la atención, la cognición y la acción,[13] mientras que la corteza prefrontal ventral se interconecta con regiones implicadas en la emoción.[14] La corteza prefrontal también recibe información de los sistemas de arousal del tronco del encéfalo, y su función es particularmente dependiente de su ambiente neuroquímico.[15] Así, existe una coordinación entre el grado de arousal o activación general, y el estado mental.[16]
Evolución
Investigadores de la Universidad de Misuri, estudiaron el motivo del aumento desmesurado de la corteza prefrontal, en comparación con el de otras especies, y sugieren que existe un factor clave para que esto se haya producido: un proceso de presión demográfica. Los investigadores afirman que, a medida que aumentaba el número de personas en la sociedad y sus interacciones, mayor era el tamaño del cerebro. Otros investigadores postulan que el desarrollo de la capacidad de manipular a los demás (o el engaño táctico) fue importante para la evolución del cerebro.[17]
Estudios
Posiblemente, el primer caso documentado sobre la función de la corteza prefrontal es el de Phineas Gage, cuyo lóbulo frontal fue destrozado cuando accidentalmente su cabeza fue atravesada por una barra de hierro en el año 1848. A pesar de que Gage conservó sus capacidades memorísticas, el habla y las habilidades motoras, su personalidad cambió radicalmente: se convirtió en una persona irritable, de fuerte temperamento e impaciente -características que no había manifestado antes de su accidente-, de forma que sus amigos manifestaron que no parecía tratarse de la misma persona. A pesar de que siempre había sido considerado como un trabajador eficiente y competente, nunca más fue capaz de completar adecuadamente sus tareas laborales. No obstante, un análisis cuidadoso de las pruebas muestran que las descripciones de los cambios en la personalidad de Gage descritos por amigos y familiares son exagerados en comparación con las descripciones realizadas por el médico de Gage. Así, se observa que los cambios descritos años después de su muerte son mucho más acentuados que los que se habían descrito mientras Gage aún vivía.[18][19]
Otros estudios posteriores realizados con pacientes afectados de daños prefrontales han mostrado que los pacientes verbalizan correctamente cuáles serían las respuestas sociales más apropiadas bajo determinadas circunstancias. No obstante, durante su vida cotidiana, prefieren ejecutar conductas que conlleven una gratificación inmediata a pesar de saber que las consecuencias a largo plazo serán negativas.
La interpretación de estos datos indican que en la corteza prefrontal no sólo se encuentran las habilidades de comparar y comprender las consecuencias futuras del comportamiento, sino que también controla la capacidad mental de demorar una gratificación inmediata en aras de lograr una gratificación mayor a largo plazo. Esta capacidad para esperar hasta obtener una recompensa es una de las claves que definen el funcionamiento óptimo de la función ejecutiva en el cerebro humano.
Existe abundante investigación encaminada a mejorar la comprensión del papel de la corteza prefrontal en los trastornos neurológicos. Muchos trastornos, como la esquizofrenia, el trastorno bipolar o el trastorno por déficit de atención con hiperactividad, han sido relacionados con una disfunción de la corteza prefrontal, por lo que se trata de un área cerebral que presenta un foco potencial para el tratamiento de este tipo de desórdenes. Se han comenzado a realizar ensayos clínicos con ciertos fármacos que han mostrado una mejora de la función de la corteza prefrontal, como la guanfacina, un agonista adrenérgico que actúa sobre los receptores alfa 2. El canal HCN, uno de los principales objetivos de este fármaco, es una de las áreas recientemente investigadas por los estudios farmacológicos relacionados con la corteza prefrontal.[20]
La teoría de la interferencia puede dividirse en tres subtipos: proactiva, retroactiva y de salida. La interferencia proactiva se relacionó con la corteza prefrontal ventrolateral y la corteza prefrontal anterior izquierda usando la tarea de los estímulos sonda recientes.[21] La interferencia retroactiva se ha relacionado con la corteza prefrontal ventral anterior mediante estudios con magnetoelectroencefalografía que estudiaban la relación entre la interferencia retroactiva y la memoria de trabajo en adultos de edad avanzada. Estos trabajos hallaron que los adultos de entre 55 y 67 años mostraban una menor actividad magnética en su corteza prefrontal que la reflejada por los sujetos del grupo control.[22]
Otros trastornos
En las últimas décadas se han utilizado técnicas de neuroimagen para determinar el volumen y las conexiones nerviosas de las diferentes regiones cerebrales. Varios estudios han indicado que se observa un escaso volumen y un reducido número de interconexiones entre los lóbulos frontales y otras regiones cerebrales en pacientes afectados por trastorno por déficit de atención con hiperactividad, esquizofrenia, depresión, trastorno bipolar, situaciones de estrés elevado,[23] víctimas de suicidio,[24] criminales encarcelados, sociópatas, víctimas de envenenamiento,[25] y drogadictos. Se cree que al menos algunas de las capacidades humanas para sentir culpa o remordimientos, y para interpretar la realidad están relacionadas con la corteza prefrontal.[26] También está ampliamente aceptado el hecho de que el tamaño y el número de conexiones en la corteza prefrontal están directamente relacionados con la capacidad de experimentar emociones.
Funciones ejecutivas
Los estudios originales de Fuster y de Goldman-Rakic hicieron hincapié en la capacidad fundamental de la corteza prefrontal para representar la información que no se encuentra presente en el medio ambiente en un momento dado, y el papel decisivo de esta función en la creación de un «bloc de notas mental». Los trabajos de Goldman-Rakic describen el modo en que este conocimiento representacional puede ser usado para guiar de forma inteligente los pensamientos, las acciones y las emociones, formando parte de este proceso la capacidad de inhibir los pensamientos, acciones y sensaciones que se consideran inapropiados.[27] De este modo, la memoria de trabajo puede ser entendida como parte fundamental del proceso de atención e inhibición comportamental. Fuster habló sobre cómo esta capacidad prefrontal permite unir pasado y futuro, posibilitando la realización de asociaciones a través del tiempo y de la modalidad sensorial para establecer ciclos de acción-percepción encaminados a la obtención de metas concretas.[28] Esta capacidad representativa constituye la base del resto de funciones ejecutivas de mayor nivel.
Shimamura propuso la teoría del filtrado Dinámico para describir el papel de la corteza prefrontal en las funciones ejecutivas. Se presume que la corteza prefrontal actúa como un mecanismo de filtrado de alto nivel que favorece las activaciones orientadas a metas e inhibe las activaciones que pueden resultan irrelevantes o distractoras. Este mecanismo de filtrado establece un control ejecutivo a distintos niveles de procesamiento (selección, mantenimiento, actualización y redireccionamiento de activaciones). Este mecanismo también ha sido utilizado para explicar la regulación emocional.[29]
Miller y Cohen propusieron una teoría integrativa del funcionamiento de la corteza prefrontal que se deriva de los trabajos originales de Goldman-Rakic y Fuster. Ambos teorizaban que el control cognitivo surge del mantenimiento activo de patrones de actividad en la corteza prefrontal dirigidos a crear representaciones de las metas a alcanzar y los medios necesarios para lograrlo. Así, la corteza prefrontal proporcionaría señales a otras estructuras cerebrales para guiar el flujo de actividad a lo largo de las rutas neuronales, estableciendo un mapeado neuronal apropiado a partir de la información entrante, los estados internos y las necesidades de respuesta para ejecutar una tarea determinada.[30] En síntesis, según esta teoría integrativa, la corteza prefrontal guía la información entrante y las conexiones que posibilitan el control cognitivo de las acciones.
La corteza prefrontal adquiere una importancia significativa cuando se requiere la puesta en marcha de un procesamiento «de arriba a abajo». Esto tiene lugar cuando la conducta está guiada por estados internos o intenciones. De acuerdo con Miller y Cohen, la corteza prefrontal es vital en aquellas situaciones en las que el mapeado de las rutas de activación formado entre la información sensorial entrante, los pensamientos y las acciones, está débilmente establecido en comparación con otros mapeados existentes.[30] Puede encontrarse un ejemplo de esto en el Test de Clasificación de Cartas de Wisconsin. Los sujetos participantes de esta tarea deben clasificar las cartas en función de su forma, color o número de símbolos que contienen. Cualquier carta puede asociarse con un número de acciones diferentes (en función de bajo qué criterio se decida clasificar la carta), por lo que no puede existir un único mapeado de respuesta ante ese estímulo. Los participantes con daños en la corteza prefrontal son capaces de clasificar las cartas en las primeras tareas de clasificación, pero cuando las reglas se complican, introduciéndose nuevos criterios posibles de clasificación, se muestran incapaces de completar la tarea correctamente.
Miller y Cohen concluyen que las implicaciones de su teoría pueden explicar en qué medida la corteza prefrontal es responsable del guiado de las acciones cognitivas. En palabras de los propios investigadores, «dependiendo del objetivo, las representaciones formadas en la corteza prefrontal pueden funcionar como plantillas atencionales, reglas, o metas a alcanzar, enviando señales a otras áreas cerebrales a través de un proceso “de arriba a abajo” que guían el flujo de actividad a través de las rutas necesarias para ejecutar una tarea».[30]
Referencias
- «Davidson, R. J., Jackson, D. C., & Kalin, N. H. (2000). Emotion, plasticity, context, and regulation: Perspectives from affective neuroscience. Psychological Bulletin, 126(6), 890–909. https://doi.org/10.1037/0033-2909.126.6.890».
- Yang Y., Raine A. (noviembre de 2009). «Prefrontal structural and functional brain imaging findings in antisocial, violent, and psychopathic individuals: a meta-analysis». Psychiatry Res (en inglés) 174 (2): 81-88. PMC 2784035. PMID 19833485. doi:10.1016/j.pscychresns.2009.03.012. Consultado el 19 de julio de 2011.
- Miller E.K., Freedman D.J., Wallis J.D. (agosto de 2002). «The prefrontal cortex: categories, concepts and cognition» (PDF). Philos. Trans. R. Soc. Lond., B, Biol. Sci. (en inglés) 357 (1424): 1123-1136. PMC 1693009. PMID 12217179. doi:10.1098/rstb.2002.1099. Consultado el 19 de julio de 2011.
- DeYoung C. G., Hirsh J. B., Shane M. S., Papademetris X., Rajeevan N., Gray J. R. (2010). «Testing predictions from personality neuroscience». Psychological Science (en inglés) 21 (6): 820-828. PMC 3049165. PMID 20435951. doi:10.1177/0956797610370159. Consultado el 19 de julio de 2011.
- Finger, Stanley (1994). Origins of neuroscience: a history of explorations into brain function (en inglés). Oxford (Oxfordshire): Oxford University Press. ISBN 0-19-514694-8.
- Preuss T.M. (1995). «Do rats have prefrontal cortex? The Rose-Woolsey-Akert program reconsidered» (PDF). The Journal of Cogntive Neuroscience (en inglés) 7 (1): 1-24. doi:10.1162/jocn.1995.7.1.1. Archivado desde el original el 25 de enero de 2012. Consultado el 19 de julio de 2011.
- Uylings H.B., Groenewegen H.J., Kolb B. (noviembre de 2003). «Do rats have a prefrontal cortex?». Behavioural Brain Research (en inglés) 146 (1-2): 3-17. PMID 14643455. doi:10.1016/j.bbr.2003.09.028.
- Rose J.E., Woolsey C.N. (1948). «The orbitofrontal cortex and its connections with the mediodorsal nucleus in rabbit, sheep and cat». Research Publications - Association for Research in Nervous and Mental Disease (en inglés). 27 (vol. 1): 210-232. PMID 18106857.
- Preuss T.M., Goldman-Rakic P.S. (agosto de 1991). «Myelo- and cytoarchitecture of the granular frontal cortex and surrounding regions in the strepsirhine primate Galago and the anthropoid primate Macaca». The Journal of Comparative Neurology (en inglés) 310 (4): 429-474. PMID 1939732. doi:10.1002/cne.903100402.
- Fuster, Joaquin M. (2008). «The Prefrontal Cortex». Academic Press (en inglés) (4.ª edición) (Boston). ISBN 0-12-373644-7.
- Markowitsch H.J., Pritzel, M. (1979). «The prefrontal cortex: Projection area of the thalamic mediodorsal nucleus?». Physiological Psychology (en inglés) 7 (1): 1-6.
- Ferrier D. (1890). «The Croonian lectures on cerebral localisation. Lecture II» (PDF). The British Medical Journal (en inglés) 1 (1537): 1349-1355. PMC 2207859. PMID 20753055. doi:10.1136/bmj.1.1537.1349. Consultado el 19 de julio de 2011.
- Goldman-Rakic, P.S. (1988). «Topography of cognition: parallel distributed networks in primate association cortex» (PDF). Annual Review of Neuroscience (en inglés) 11: 137-156. PMID 3284439. doi:10.1146/annurev.ne.11.030188.001033. Consultado el 19 de julio de 2011. (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- Price, J.L. (junio de 1999). «Prefrontal cortical networks related to visceral function and mood». Annals of the New York Academy of Sciences (en inglés) 877: 383-396. PMID 10415660. doi:10.1111/j.1749-6632.1999.tb09278.x.
- Robbins T.W., Arnsten A.F. (2009). «The neuropsychopharmacology of fronto-executive function: monoaminergic modulation». Annual Review of Neuroscience (en inglés) 32: 267-287. PMC 2863127. PMID 19555290. doi:10.1146/annurev.neuro.051508.135535. Consultado el 19 de julio de 2011.
- Arnsten A.F., Paspalas C.D., Gamo N.J., Yang Y., Wang M. (agosto de 2010). «Dynamic Network Connectivity: A new form of neuroplasticity». Trends in Cognitive Science (en inglés) 14 (8): 365-375. PMC 2914830. PMID 20554470. doi:10.1016/j.tics.2010.05.003.
- Manes, Facundo; Niro, Mateo (2014). Usar el cerebro. Buenos Aires: Planeta. ISBN 978-950-49-3982-5.
- Macmillan, Malcom (julio de 2001). «An Odd Kind of Fame: Stories of Phineas Gage» (PDF). Journal of Neurology Neurosurgery and Psychiatry (en inglés) 71 (1): 136. doi:10.1136/jnnp.71.1.136c. Consultado el 19 de julio de 2011.
- Macmillan, M. (2008 @ mes=Septiembre). «Phineas Gage – Unravelling the myth» (PDF). The Psychologist (en inglés) (British Psychological Society) 21 (9): 828-831. Archivado desde el original el 4 de junio de 2012. Consultado el 19 de julio de 2011.
- Wang, M. et al. (2007). «Alpha2A-adrenoceptors strengthen working memory networks by inhibiting cAMP-HCN channel signaling in prefrontal cortex». Cell (en inglés) 129 (2): 397-410. PMID 17448997. doi:10.1016/j.cell.2007.03.015.
- Jonides J., Nee D.E. (2006). «Brain Mechanisms of Proactive Interference in Working Memory» (PDF). Neuroscience (en inglés) 139 (1): 181-193. PMID 16337090. doi:10.1016/j.neuroscience.2005.06.042. Consultado el 19 de julio de 2011.
- Solesio E., Lorenzo-López L., Campo P., López-Frutos J.M., Ruiz-Vargas J.M., Maestú F. (2009). «Retroactive interference in normal aging: A magnetoencephalography study». Neuroscience Letters (en inglés) 456 (2): 85-88. PMID 19429139. doi:10.1016/j.neulet.2009.03.087.
- Liston C et al. (2006). «Stress-induced alterations in prefrontal cortical dendritic morphology predict selective impairments in perceptual attentional set-shifting». Journal of Neuroscience (en inglés) 26 (30): 7870-7874. PMID 16870732. doi:10.1523/JNEUROSCI.1184-06.2006.
- Rajkowska G. (1997). «Morphometric methods for studying the prefrontal cortex in suicide victims and psychiatric patients». Annals of New York Academic of Sciences (en inglés) 836: 253-268. PMID 9616803. doi:10.1111/j.1749-6632.1997.tb52364.x.
- Cecil K.M., Brubaker C.J., Adler C.M., Dietrich K.N., Altaye M., Egelhoff J.C., Wessel S., Elangovan I., Hornung R. et al. (2008). «Decreased brain volume in adults with childhood lead exposure». En John Balmes, ed. PLoS Med (en inglés) 5 (5): e112. PMC 2689675. PMID 18507499. doi:10.1371/journal.pmed.0050112. Consultado el 19 de julio de 2011.
- Anderson, S.W.; Bechara, A.; Damasio, H.; Tranel, D.; Damasio, A.R. (1999). «Impairment of social and moral behavior related to early damage in human prefrontal cortex». Nature Neuroscience (en inglés) 2 (11): 1032-1037. PMID 10526345. doi:10.1038/14833. Consultado el 19 de julio de 2011.
- Goldman-Rakic, P.S.; Cools, A. R.; Srivastava, K. (1996). «The prefrontal landscape: implications of functional architecture for understanding human mentation and the central executive». Philosophical Transactions of the Royal Society of London: Biologycal Sciences (en inglés) 351 (1346): 1445-1453. JSTOR 3069191. PMID 8941956. doi:10.1098/rstb.1996.0129.
- Fuster J.M., Bodner M., Kroger J.K. (2000). «Cross-modal and cross-temporal association in neurons of frontal cortex» (PDF). Nature (en inglés) 405 (6784): 347-351. PMID 10830963. doi:10.1038/35012613. Consultado el 19 de julio de 2011.
- Shimamura, A. P. (2000). «The role of the prefrontal cortex in dynamic filtering» (PDF). Psychobiology (en inglés) 28 (2): 207-218. Consultado el 19 de julio de 2011.
- Miller E.K., Cohen J.D. (2001). «An integrative theory of prefrontal cortex function». Annual Review of Neuroscience (en inglés) 24: 167-202. PMID 11283309. doi:10.1146/annurev.neuro.24.1.167.
Enlaces externos
 Wikimedia Commons alberga una categoría multimedia sobre Corteza prefrontal.
Wikimedia Commons alberga una categoría multimedia sobre Corteza prefrontal.
 Datos: Q18680
Datos: Q18680- Multimedia: Prefrontal cortex / Q18680