Avimimus
Avimimus (lat. "imitador de aves") es un género extinto de dinosaurio terópodo avimímido, que vivió a finales del período el Cretácico, hace aproximadamente 70 millones de años, en el Campaniense, en lo que hoy es Asia. El primer espécimen fue descubierto en la Formación de Djadochta y fue descrito oficialmente por el Doctor Sergi Kurzanov en 1981.[1] En 1991, Sankar Chatterjee creó el orden Avimimiformes para incluir a Avimimus, pero actualmente está en desuso. La especie tipo es Avimimus portentosus una segunda especie fue nombrada en 2018 llamada Avimimus nemegtensis.[2][3]
| Avimimus | ||
|---|---|---|
| Rango temporal: 70 Ma Cretácico Superior | ||
 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| Infraorden: | Oviraptorosauria | |
| Superfamilia: | Caenagnathoidea | |
| Familia: |
Avimimidae Kurzanov, 1981 | |
| Género: |
Avimimus Kurzanov, 1981 | |
| Especie tipo | ||
|
Avimimus portentosus Kurzanov, 1981 | ||
| Otras Especies | ||
| ||
Descripción
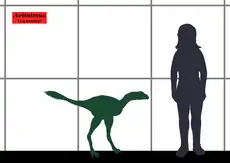
Avimimus era un dinosaurio pequeño, de 45 centímetros de altura hasta la cadera y un largo de alrededor de 1,6 metros.[1] Su cráneo, pequeño en comparación con el cuerpo, pero tenía un gran cerebro[1] y unas cuencas oculares desmesuradas. El tamaño de los huesos que rodean la cavidad craneana y estarían dedicados a proteger el encéfalo eran grandes.[1] Esto es consistente con el hecho de que Avimimus haya tenido un gran cerebro.[1]

Como en los cercanos Oviraptoridae y Caenagnathidae, en el lugar de los dientes, presentaba un pico córneo, muy semejante al de los loros. A diferencia de estos, una serie de proyecciones similares a dientes en el borde premaxilar pudo haberle dado un borde aserrado. El pico desdentado de Avimimus sugiere que debe habes sido herbívoro o omnívoro. Kurzanov por su lado, cree que Avimimus habría sido insectívoro.[1] El foramen magnum, el agujero que permite que la médula espinal conecte con el cerebro, era proporcionalmente grande en Avimimus.[1] El cóndilo occipital, en cambio, era pequeño, lo que sugiere que la cabeza era pequeña.[1] El cuello en sí mismo era largo y delgado, y se compone de vértebras mucho más alargadas que en otros oviraptorosaurianos. A diferencia que los oviraptóridos y cenagnátidos, carencia de las aberturas en la de la parte posterior de las vértebras para sacos aéreos , sugiriendo que Avimimus es más primitivo que estos animales.
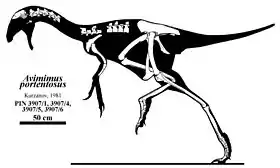
Los brazos son relativamente cortos. Los huesos de la mano se fusionan juntos, como en las aves modernas, y un canto en el cúbito (hueso de un brazo más bajo) fue interpretado como punto de accesorio para las plumas por Kurzanov.[2][1] Kurzanov, en 1987, también divulgó la presencia de una quilla,[4][1] y mientras que Chiappe confirmó la presencia de topetones en el cúbito, su función seguía siendo confusa.[5] Kurzanov, convencido de que eran puntos de accesorio para las plumas que concluyó que Avimimus puede haber sido capaz de un vuelo débil.[1] La presencia de plumas ahora se acepta extensamente, pero la mayoría de los paleontólogos no creen que Avimimus pudiera volar.[1] El ilion estaba orientado casi horizontalmente, dando por resultado caderas excepcionalmente amplias. Poco se sabe de cola pero la cadera sugiere que la cola fuera larga. Con piernas extremadamente largas y delgadas, sugieren que Avimimus era un corredor sumamente especializado. Las proporciones de los huesos de la pierna agregan peso adicional a la idea de Avimimus tenía comportamiento cursorial.[1] Las espinillas eran largas en comparación con sus muslos,[1] rasgo común con los animales corredores. Habría pisado sobre tres dedos con unas garras estrechas. Debido a la ausencia de cola en el fósil se pensó que no habría tenido una en vida.[1] Sin embargo, posteriores descubrimientos de vértebras caudales de Avimimus han confirmado su presencia.[1]
Descubrimiento e investigación
Los restos de Avimimus fueron recuperados por los paleontólogos rusos[1] y descritos oficialmente por el Dr. Sergei Kurzanov en 1981. Los fósiles de Avimimus fueron descritos inicialmente como provenientes de la Formación Djadokta por Kurzanov. Sin embargo, en la descripción de una nueva muestra de 2006, Watabe y colegas observaron que Kurzanov probablemente se equivocó acerca de la procedencia, y es más probable de que Avimimus provenga de la más reciente Formación Nemegt.[6] Los especie tipo es A. portentosus.[2] Un segundo espécimen casi completo de Avimimus fue descubierto en 1996 y se describieron en el año 2000 por Watabe y colegas. Además, estos autores identificaron una serie de pequeñas huellas de terópodos en la misma zona que pertenecerian a Avimimus.[7]
Una variedad de huesos aislados que se han atribuido a Avimimus pero difieren de A. portentosus y puede representar una o más especies distintas, referidas como Avimimus sp..[8]
En 2008, un equipo de Canadá, Estados Unidos, y paleontólogos mongoles encabezada por Philip J. Currie anunció el descubrimiento de una extensa cama de huesos de Avimimus sp.. El lecho se encuentra en la Formación Nemegt, 10,5 metros por encima de la Formación Barun Goyot, en el desierto de Gobi. El equipo informó del hallazgo de abundantes huesos de al menos diez individuos de Avimimus, pero el depósito puede contener más. Todos los individuos eran adultos o subadultos, y los adultos mostraron poca variación en tamaño, lo que sugiere crecimiento determinado. El equipo también sugiere que los individuos se encuentran juntos porque eran gregarios en vida, proporcionando posibles indicios de que Avimimus formara grupos segregados por edades, ya sea para reproducción o propósitos de migración. Los adultos mostraron un mayor grado de fusión del esqueleto en el tarso-metatarso y tibio-tarso, y cicatrices musculares también más prominentes. La preservación de los lechos de huesos sugieren que fueron enterrados rápidamente, descubiertos por un flujo rápido de agua, y luego enterrados de nuevo a una corta distancia.[9][10] En 2018, estos restos de Avimimus sp. fueron descritos formalmente como una nueva especie, A. nemegtensis.[3]
Clasificación
A Avimimus se lo consideró originalmente como un pariente muy cercano de las aves, dado su especial conjunto de características de aves no conocidas en otros dinosaurios en el momento. De hecho, Kurzanov argumentó que Avimimus, en lugar de la famosa ave temprana Archaeopteryx, estaba más cerca del antepasado directo de las aves modernas, y que el Archaeopteryx no estaba tan estrechamente relacionado con las aves como previamente se había sugerido.[4] Sin embargo, este punto de vista no ha sido apoyado por posteriores análisis filogenéticos de las relaciones de dinosaurios y aves. La mayoría de los científicos modernos encuentran que Avimimus , de hecho, pertenece a un grupo diverso de dinosaurios similares a las aves más primitivos que el Archaeopteryx, los oviraptorosaurios.[11][12]
Kurzanov coloco a Avimimus en su propia familia, Avimimidae, en 1981. En 1991, Sankar Chatterjee erigió el Orden Avimimiformes para incluir Avimimus. Ninguno de estos nombres de grupo se utiliza con frecuencia por los paleontólogos , ya que incluyen una sola especie . Estudios más recientes han demostrado que Avimimus se agrupa mejor dentro de Oviraptoridae , dentro del subgrupo Elmisaurinae.[13]
El siguiente cladograma sigue un análisis realizado por Phil Senter, 2007.[14]
| Caenagnathoidea |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleoecología
La Formación Barun Goyot de Mongolia, se estima que se remontan a la etapa Campaniense, hace entre 84 y 70 millones de años[15] de cretáceo tardío.[16] Durante el período cretáceo, lo que ahora es la Formación Barun Goyot tenían un ambiente árido con campos dedunas de arena y solo intermitentes corrientes de agua. Es ligeramente más joven que la cercana Formación Djadochta, y parece haber sido un poco más húmedo.[16] Esta formación se destaca por la exquisita conservación de los pequeñas y frágiles esqueletos de dinosaurios, un acontecimiento raro teniendo en cuenta que estos fósiles normalmente se rompen y se dispersan cuando se encuentran en otras formaciones rocosas.[17]
La región que se conserva en la Formación Barun Goyot era el hogar de la maniraptor Hulsanpes perlei, los oviraptóridos Conchoraptor gracilis y Ajancingenia yanshini , los alvarezsáuridos Ceratonykus oculatus , Mononykus y Parvicursor remotus , el paquicefalosáurido Tylocephale gilmorei, los anquilosáuridos Saichania chulsanensis y Tarchia gigantea , y los ceratópsidos Bagaceratops rozhdestvenskyi , Breviceratops kozlowskii, Lamaceratops tereschenkoi y Platyceratops tatarinovi. El dinosaurio más grande de ellos fue el titanosauriano Quaesitosaurus orientalis. Se observó que muchos de los mismos géneros estuvieron presentes en la Formaciones Barun Goyot y Djadochta, aunque hubo variaciones a nivel de especie.[18] Los vertebrados presentes en la Formación Barun Goyot incluye las aves primitivas Gobipteryx minuta y Hollanda luceria y lagartos Estesia mongoliensis, Ovoo gurvel, Proplatynotia longirostrata y Gobiderma pulchrum. Los primeros mamíferos que estaban presentes en esta región durante la época de Avimimus eran los mamíferos placentarios Asioryctes nemegetensis y Barunlestes butleri, el anfibio Gobiates khermeentsavi, los multituberculados mamíferos Catopsbaatar catopsaloides, Chulsanbaatar vulgaris y Nemegtbaatar gobiensis, y los marsupiales Asiatherium reshetovi y Deltatheridium pretrituberculare. Numerosas cáscaras de huevo de dinosaurios se encuentran en esta región apoyan la presencia de saurópodos y maniraptores.
Referencias
- "Avimimus." In: Dodson, Peter & Britt, Brooks & Carpenter, Kenneth & Forster, Catherine A. & Gillette, David D. & Norell, Mark A. & Olshevsky, George & Parrish, J. Michael & Weishampel, David B. The Age of Dinosaurs. Publications International, LTD. p. 130. ISBN 0-7853-0443-6.
- Kurzanov, S.M. (1981). "An unusual theropod from the Upper Cretaceous of Mongolia Iskopayemyye pozvonochnyye Mongolii (Fossil Vertebrates of Mongolia)." Trudy Sovmestnay Sovetsko-Mongolskay Paleontologiyeskay Ekspeditsiy (Joint Soviet-Mongolian Paleontological Expedition), 15: 39-49. Moscú: Naúka, 1981
- Funston, G.F.; Mendonca, S.E.; Currie, P.J.; Barsbold, R. (2018). «Oviraptorosaur anatomy, diversity and ecology in the Nemegt Basin». Palaeogeography, Palaeoclimatology, Palaeoecology. in press. doi:10.1016/j.palaeo.2017.10.023.
- Kurzanov, S.M. (1987). "Avimimidae and the problem of the origin of birds." Transactions of the Joint Soviet-Mongolian Paleontological Expedition, 31: 5-92. [in Russian]
- Chiappe, L.M. and Witmer, L.M. (2002). Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press, 536 pp. ISBN 0-520-20094-2
- Watabe; Suzuki; Tsogtbaatar (2006). «Geological and geographical distribution of bird-like theropod, Avimimus in Mongolia». Journal of Vertebrate Paleontology 26 (3): 136A-137A.
- Watabe, Weishampel; Barsbold, Tsogtbaatar; Suzuke (2000). «New nearly complete skeleton of the bird-like theropod, Avimimus, from the Upper Cretaceous of the Gobi Desert, Mongolia». Journal of Vertebrate Paleontology 20 (3): 77A.
- Ryan, Currie; Russell, D. (2001). «New material of Avimimus portentosus (Theropoda) from the Iren Debasu Formation (Upper Cretaceous) of the Erenhot Region of Inner Mongolia». Journal of Vertebrate Paleontology 21 (3): 95A.
- Currie, P.; Longrich, N.; Ryan, M.; Eberth, D.; Demchig, B. (2008). «A bonebed of Avimimus sp. (Dinosauria: Theropoda) from the Late Cretaceous Nemegt Formation, Gobi Desert: Insights into social behavior and development in a maniraptoran theropod.». Journal of Vertebrate Paleontology 28 (3): 67A.
- Funston, G.F.; Currie, P.J.; Eberth, D.A.; Ryan, M.J.; Chinzorig, T.; Badamgarav, D.; Longrich, N.R. (2016). «The first oviraptorosaur (Dinosauria: Theropoda) bonebed: evidence of gregarious behaviour in a maniraptoran theropod». Scientific Reports 6: 35782. PMC 5073311. PMID 27767062. doi:10.1038/srep35782.
- Dyke; Thorley (1998). «Reduced cladistic consensus methods and the avian affinities of Protoavis and Avimimus». Archaeopteryx 16: 123-129.
- Turner, Alan H.; Pol, Diego; Clarke, Julia A.; Erickson, Gregory M.; Norell, Mark (2007). «A basal dromaeosaurid and size evolution preceding avian flight» (pdf). Science 317 (5843): 1378-1381. PMID 17823350. doi:10.1126/science.1144066.
- Holtz, Thomas R. Jr. (2010) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2010 Appendix.
- Senter, P. (2007). "A new look at the phylogeny of Coelurosauria (Dinosauria: Theropoda)." Journal of Systematic Palaeontology, (doi 10.1017/S1477201907002143).
- Gradstein, Felix M.; Ogg, James G.; Smith, Alan G. (2005). A Geologic Time Scale 2004. Cambridge: Cambridge University Press. ISBN 978-0-521-78142-8.
- Jerzykiewicz, Tomasz; Russell, Dale A. (1991). «Late Mesozoic stratigraphy and vertebrates of the Gobi Basin». Cretaceous Research 12 (4): 345-377. doi:10.1016/0195-6671(91)90015-5.
- Novacek, M. J., 1996. "Dinosaurs of the Flaming. Cliffs" Anchor/Doubleday. 367 p.
- Nicholas R. Longrich; Philip J. Currie; Dong Zhi-Ming (2010). «A new oviraptorid (Dinosauria: Theropoda) from the Upper Cretaceous of Bayan Mandahu, Inner Mongolia». Palaeontology 53 (5): 945-960. doi:10.1111/j.1475-4983.2010.00968.x.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikimedia Commons alberga una categoría multimedia sobre Avimimus.
Wikimedia Commons alberga una categoría multimedia sobre Avimimus. Wikispecies tiene un artículo sobre Avimimus.
Wikispecies tiene un artículo sobre Avimimus.- Avimimus en la Fossilworks Archivado el 24 de febrero de 2014 en Wayback Machine. (en inglés)
- Avimimus dentro de Oviraptorosauria en The Thescelsosaurus (en inglés)
- Ficha e imagen de Avimimus en Dinodata.de (en alemán)
- Avimimus en the Theropod Database (en inglés)