Conversión génica
La conversión de genes es el proceso por el cual una secuencia de ADN reemplaza a una secuencia homóloga de manera que las secuencias se vuelven idénticas después del evento de conversión. La conversión de genes puede ser alélica, lo que significa que un alelo del mismo gen reemplaza a otro alelo, o ectópica, lo que significa que una secuencia de ADN paráloga convierte a otra.
Conversión de genes alélicos
La conversión del gen alélico ocurre durante la meiosis cuando la recombinación homóloga entre sitios heterocigóticos da como resultado un desajuste en el apareamiento de bases. Este desajuste es luego reconocido y corregido por la maquinaria celular, lo que hace que uno de los alelos se convierta en el otro. Esto puede causar una segregación no mendeliana de alelos en las células germinales.[1]
Conversión genética no alélica/ectópica
La recombinación ocurre no solo durante la meiosis, sino también como un mecanismo para la reparación de roturas de doble cadena (DSB, del inglés double-strand breaks) causadas por daños en el ADN. Estos DSB generalmente se reparan utilizando la cromátida hermana del dúplex roto y no el cromosoma homólogo, por lo que no darían como resultado una conversión alélica. La recombinación también ocurre entre secuencias homólogas presentes en diferentes loci genómicos (secuencias parálogos) que han resultado de duplicaciones de genes previas. Se supone que la conversión de genes que se produce entre secuencias parálogas (conversión de genes ectópicos) es responsable de la evolución concertada de familias de genes.[1][2][3]
Mecanismo
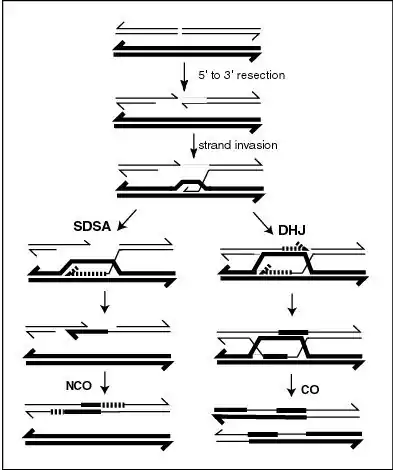
La conversión de un alelo en otro a menudo se debe a la reparación del desajuste de bases durante la recombinación homóloga: si una de las cuatro cromátidas durante la meiosis se empareja con otra cromátida, como puede ocurrir debido a la homología de secuencia, puede ocurrir la transferencia de la cadena de ADN seguida de reparación del desajuste. Esto puede alterar la secuencia de uno de los cromosomas, de modo que sea idéntico al otro.
La recombinación meiótica se inicia mediante la formación de una ruptura de doble cadena (DSB). Los extremos 5' de la ruptura se degradan luego, dejando salientes 3' largos de varios cientos de nucleótidos. Luego, uno de estos segmentos de ADN monocatenario 3' invade una secuencia homóloga en el cromosoma homólogo, formando un intermedio que puede repararse a través de diferentes vías que dan como resultado cruces (CO) o no cruzados (NCO). En varios pasos del proceso de recombinación, se forma ADN heterodúplex (ADN de doble hebra que consta de hebras simples de cada uno de los dos cromosomas homólogos que pueden ser perfectamente complementarios o no). Cuando ocurren desajustes en el ADN heterodúplex, la secuencia de una hebra se reparará para unir la otra hebra con perfecta complementariedad, lo que conducirá a la conversión de una secuencia en otra. Este proceso de reparación puede seguir cualquiera de dos vías alternativas como se ilustra en la Figura. Por una vía, se forma una estructura llamada unión doble de Holliday (DHJ), que conduce al intercambio de hebras de ADN. Por la otra vía, conocida como Recocido de hebras dependientes de síntesis (SDSA), hay intercambio de información pero no intercambio físico. La conversión de genes ocurrirá durante SDSA si las dos moléculas de ADN son heterocigotas en el sitio de la reparación recombinacional. La conversión de genes también puede ocurrir durante la reparación recombinacional que involucra un DHJ, y esta conversión de genes puede estar asociada con la recombinación física de los dúplex de ADN en los dos lados del DHJ.
Conversión genética sesgada versus imparcial
La conversión de genes sesgada (BGC, del inglés Biased gene conversion) ocurre cuando un alelo tiene una mayor probabilidad de ser el donante que el otro en un evento de conversión de genes. Por ejemplo, cuando ocurre un desajuste de T:G, sería más o menos probable que se corrija a un par C:G que a un par T:A. Esto le da a ese alelo una mayor probabilidad de transmisión a la siguiente generación. La conversión de genes no sesgada significa que ambas posibilidades ocurren con la misma probabilidad.
Conversión génica sesgada por GC
La conversión de genes sesgada por GC (gBGC) es el proceso mediante el cual el contenido de GC del ADN aumenta debido a la conversión de genes durante la recombinación.[2] Existe evidencia de gBGC para levaduras y humanos y la teoría se ha probado más recientemente en otros linajes eucariotas.[4] En las secuencias de ADN humano analizadas, se ha encontrado que la tasa de cruzamiento se correlaciona positivamente con el contenido de GC. Las regiones pseudoautosómicas (PAR) de los cromosomas X e Y en humanos, que se sabe que tienen altas tasas de recombinación, también tienen altos contenidos de GC.[1] Ciertos genes de mamíferos que experimentan una evolución concertada (por ejemplo, operones ribosómicos, ARNt y genes de histonas) son muy ricos en GC. Se ha demostrado que el contenido de GC es mayor en los genes de histonas parálogos humanos y de ratón que son miembros de grandes subfamilias (presumiblemente en evolución concertada) que en los genes de histonas parálogos con secuencias relativamente únicas.[5] También hay evidencia de sesgo de GC en el proceso de reparación de desajustes. Se cree que esto puede ser una adaptación a la alta tasa de desaminación de metil-citosina que puede conducir a transiciones C → T.
BGC del gen Fxy en Mus musculus
El gen Fxy o Mid1 en algunos mamíferos estrechamente relacionados con los ratones domésticos (humanos, ratas y otras especies de Mus) se encuentra en la región ligada al sexo del cromosoma X. Sin embargo, en Mus musculus, se ha translocado recientemente de manera que el extremo 3' del gen se solapa con la región PAR del cromosoma X, que se sabe que es un punto caliente de recombinación. Esta porción del gen ha experimentado un aumento dramático en el contenido de GC y la tasa de sustitución en la posición del tercer codón, así como en los intrones, pero la región 5' del gen, que está ligada al X, no lo ha hecho. Debido a que este efecto está presente solo en la región del gen que experimenta una mayor tasa de recombinación, debe deberse a una conversión genética sesgada y no a una presión selectiva.[2]
Impacto de la conversión de genes sesgada por GC en los patrones genómicos humanos
El contenido de GC varía ampliamente en el genoma humano (40-80%), pero parece haber grandes secciones del genoma donde el contenido de GC es, en promedio, mayor o menor que en otras regiones.[1] Estas regiones, aunque no siempre muestran límites claros, se conocen como isocoros. Una posible explicación para la presencia de isocoros ricos en GC es que evolucionaron debido a la conversión de genes sesgada por GC en regiones con altos niveles de recombinación.
Importancia evolutiva
Función adaptativa de la recombinación
Los estudios de conversión de genes han contribuido a nuestra comprensión de la función adaptativa de la recombinación meiótica. El patrón de segregación ordinario de un par de alelos (Aa) entre los 4 productos de la meiosis es 2A:2a. La detección de eventos de conversión génica poco frecuentes (por ejemplo, patrones de segregación 3:1 o 1:3 durante las meiosis individuales) proporciona información sobre las vías alternativas de recombinación que conducen a cromosomas cruzados o no cruzados. Se cree que los eventos de conversión de genes surgen cuando los alelos "A" y "a" están cerca de la ubicación exacta de un evento de recombinación molecular. Por tanto, es posible medir la frecuencia con la que los eventos de conversión génica se asocian con el cruce o no entrecruzamiento de regiones cromosómicas adyacentes, pero fuera del evento de conversión inmediato. Se han llevado a cabo numerosos estudios de conversión génica en varios hongos (que son especialmente adecuados para tales estudios).[6] De esta revisión se desprende claramente que la mayoría de los eventos de conversión de genes no están asociados con el intercambio de marcadores externos. Por lo tanto, la mayoría de los eventos de conversión de genes en los diferentes hongos estudiados están asociados con el no cruce de marcadores externos. Los eventos de conversión de genes no cruzados se producen principalmente mediante el recocido de cadena dependiente de síntesis (SDSA).[7] Este proceso implica un intercambio de información limitado, pero no un intercambio físico de ADN, entre los dos cromosomas homólogos participantes en el sitio del evento de conversión, y se produce poca variación genética. Por tanto, las explicaciones de la función adaptativa de la recombinación meiótica que se centran exclusivamente en el beneficio adaptativo de producir una nueva variación genética o intercambio físico parecen inadecuadas para explicar la mayoría de los eventos de recombinación durante la meiosis. Sin embargo, la mayoría de los eventos de recombinación meiótica pueden explicarse por la propuesta de que son una adaptación para la reparación del daño en el ADN que se transmitirá a los gametos.[8]
De particular interés, desde el punto de vista de que la recombinación es una adaptación para la reparación del ADN, son los estudios en levaduras que muestran que la conversión de genes en las células mitóticas aumenta con los rayos UV[9][10] y la radiación ionizante.[11]
Enfermedades genéticas de los humanos
En las discusiones sobre enfermedades genéticas en humanos, las conversiones de genes mediadas por pseudogenes que introducen mutaciones patógenas en genes funcionales es un mecanismo de mutación bien conocido. Por el contrario, es posible que los pseudogenes sirvan como plantillas. Durante el curso de la evolución, los genes fuente funcionales que son potencialmente ventajosos se han derivado de múltiples copias en su único gen fuente. Los cambios basados en plantillas de pseudogenes podrían eventualmente volverse fijos siempre que no tuvieran efectos deletéreos.[12] Por tanto, de hecho, los pseudogenes pueden actuar como fuentes de variantes de secuencia que pueden transferirse a genes funcionales en combinaciones novedosas y sobre las que se puede actuar mediante selección. La lectina 11 (SIGLEC 11), una inmunoglobulina humana que se une al ácido siálico, puede considerarse un ejemplo de un evento de conversión génica que ha jugado un papel importante en la evolución. Al comparar los genes homólogos de SIGLEC11 humano y su pseudogén en el chimpancé, bonobo, gorila y orangután, parece que hubo conversión génica de la secuencia de las regiones 5' y los exones que codifican el dominio de reconocimiento del ácido siálico, aproximadamente 2 kbp del pseudogén hSIGLECP16 que lo flanquea estrechamente. Las tres evidencias relativas a este evento han sugerido juntas que se trata de un cambio adaptativo que es muy importante desde el punto de vista evolutivo en el género Homo. Esos incluyen que solo en el linaje humano ocurrió esta conversión genética, la corteza cerebral ha adquirido una expresión importante de SIGLEC11 específicamente en el linaje humano y la exhibición de un cambio en la unión del sustrato en el linaje humano en comparación con el de su contraparte en los chimpancés. Por supuesto, la frecuencia de la contribución de este mecanismo de conversión de genes mediado por pseudogenes a los cambios funcionales y adaptativos en la evolución del ser humano aún se desconoce y hasta ahora apenas se ha explorado.[13] A pesar de eso, la introducción de cambios genéticos positivamente selectivos por tal mecanismo puede ser puesta a consideración por el ejemplo de SIGLEC11. A veces, debido a la interferencia de elementos transponibles en algunos miembros de una familia de genes, provoca una variación entre ellos y, finalmente, también puede detener la tasa de conversión de genes debido a la falta de similitud de secuencia que conduce a una evolución divergente.
Análisis genómico
A partir de varios análisis del genoma, se concluyó que las roturas de doble cadena (DSB) se pueden reparar mediante recombinación homóloga mediante al menos dos vías diferentes pero relacionadas.[12] En el caso de una vía principal, se emplearán secuencias homólogas en ambos lados del DSB que parece ser análogo al modelo conservador de reparación del DSB[14] que se propuso originalmente para la recombinación meiótica en levadura,[15] donde, como la vía menor está restringida a solo un lado del DSB, como lo postula el modelo de invasión unilateral no conservador.[16] Sin embargo, en ambos casos la secuencia de los socios de recombinación se conservará absolutamente. En virtud de su alto grado de homología, las nuevas copias de genes que surgieron después de la duplicación de genes tienden naturalmente a un cruce desigual o eventos de conversión de genes unidireccionales. En el último proceso, existen las secuencias aceptora y donante y la secuencia aceptora será reemplazada por una secuencia copiada del donante, mientras que la secuencia del donante permanece sin cambios.[13]
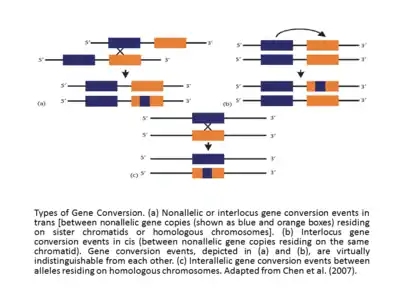
La homología efectiva entre las secuencias que interactúan hace que el evento de conversión del gen sea exitoso. Además, la frecuencia de conversión de genes es inversamente proporcional a la distancia entre las secuencias que interactúan en cis,[17][12] y la tasa de conversión de genes suele ser directamente proporcional a la longitud del tramo de secuencia ininterrumpido en la supuesta región convertida. Parece que los tramos de conversión que acompañan al cruce son más largos (longitud media = ∼460 pb) que los tramos de conversión sin cruce (longitud media = 55–290 bp).[18] En los estudios de genes de globulina humana, se ha apoyado durante mucho tiempo que el evento de conversión génica o los eventos de migración de ramas pueden ser promovidos o inhibidos por los motivos específicos que existen en la vecindad de la secuencia de ADN. Otra clasificación básica de los eventos de conversión de genes es la conversión de genes interlocus (también llamada no alélica) e interalélica. Los eventos de conversión de genes cis o trans no alélicos o interlocus ocurren entre copias de genes no alélicos que residen en cromátidas hermanas o cromosomas homólogos y, en el caso de interalélicos, los eventos de conversión de genes tienen lugar entre alelos que residen en cromosomas homólogos.[13] Si se comparan los eventos de conversión de genes interlocus, con frecuencia se revelará que exhiben una direccionalidad sesgada. A veces, como en el caso de los genes de globina humana, la dirección de conversión del gen se correlaciona con los niveles de expresión relativa de los genes que participan en el evento, con el gen expresado en un nivel superior, llamado gen 'maestro', convirtiéndolo con una expresión más baja, llamado gen 'esclavo'. Originalmente formulada en un contexto evolutivo, la regla del 'gen maestro/esclavo' debe explicarse con cautela. De hecho, el aumento en la transcripción de genes exhibe no solo el aumento en la probabilidad de que se use como donante sino también como aceptor.[19]
Efecto
Normalmente, un organismo que ha heredado diferentes copias de un gen de cada uno de sus padres se denomina heterocigoto. Esto se representa genéricamente como genotipo: Aa (es decir, una copia de la variante (alelo) 'A' y una copia del alelo 'a'). Cuando un heterocigoto crea gametos por meiosis, los alelos normalmente se duplican y terminan en una proporción de 2:2 en las 4 células resultantes que son productos directos de la meiosis. Sin embargo, en la conversión de genes, se observa una relación distinta de la esperada 2A:2a, en la que A and a son los dos alelos. Algunos ejemplos son 3A:1a y 1A:3a. En otras palabras, puede haber, por ejemplo, tres veces más alelos A que a alleles expresados en las células hijas, como es el caso de 3A:1a.
Relevancia médica
La conversión genética que da como resultado la mutación del gen CYP21A2 es una causa genética subyacente común de hiperplasia suprarrenal congénita. La conversión de genes somáticos es uno de los mecanismos que pueden resultar en retinoblastoma familiar, un cáncer congénito de la retina, y se teoriza que la conversión de genes puede jugar un papel en el desarrollo de la enfermedad de Huntington.
Referencias
- Galtier, N.; Piganeau, G.; Mouchiroud, D.; Duret, L. (2001-10). «GC-content evolution in mammalian genomes: the biased gene conversion hypothesis». Genetics 159 (2): 907-911. ISSN 0016-6731. PMC 1461818. PMID 11693127.
- Duret, Laurent; Galtier, Nicolas (28 de agosto de 2009). «Biased Gene Conversion and the Evolution of Mammalian Genomic Landscapes». Annual Review of Genomics and Human Genetics 10 (1): 285-311. ISSN 1527-8204. doi:10.1146/annurev-genom-082908-150001.
- Harpak, Arbel; Lan, Xun; Gao, Ziyue; Pritchard, Jonathan K. (28 de noviembre de 2017). «Frequent nonallelic gene conversion on the human lineage and its effect on the divergence of gene duplicates». Proceedings of the National Academy of Sciences (en inglés) 114 (48): 12779-12784. ISSN 0027-8424. PMC 5715747. PMID 29138319. doi:10.1073/pnas.1708151114.
- Pessia, Eugénie; Popa, Alexandra; Mousset, Sylvain; Rezvoy, Clément; Duret, Laurent; Marais, Gabriel A. B. (1 de enero de 2012). «Evidence for Widespread GC-biased Gene Conversion in Eukaryotes». Genome Biology and Evolution (en inglés) 4 (7): 675-682. PMC 5635611. PMID 22628461. doi:10.1093/gbe/evs052.
- Galtier, Nicolas (1 de febrero de 2003). «Gene conversion drives GC content evolution in mammalian histones». Trends in Genetics (en inglés) 19 (2): 65-68. ISSN 0168-9525. PMID 12547511. doi:10.1016/S0168-9525(02)00002-1.
- Whitehouse, H. L. K. (Harold L. K.) (1982). Genetic recombination : understanding the mechanisms. Wiley. ISBN 0-471-10205-9. OCLC 8031814.
- McMahill, Melissa S.; Sham, Caroline W.; Bishop, Douglas K. (6 de noviembre de 2007). «Synthesis-Dependent Strand Annealing in Meiosis». PLOS Biology (en inglés) 5 (11): e299. ISSN 1545-7885. PMC 2062477. PMID 17988174. doi:10.1371/journal.pbio.0050299.
- Bernstein, Harris; Bernstein, Carol; Michod, Richard E. (2011). «19. Meiosis as an Evolutionary Adaptation for DNA Repair». En Kruman, Inna, ed. DNA Repair. 2011: InTech. ISBN 978-953-307-697-3. doi:10.5772/25117.
- Ito, Takashi; Kobayashi, Katsumi (1 de octubre de 1975). «Studies of the induction of mitotic gene conversion by ultraviolet irradiation: II. Action spectra». Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis (en inglés) 30 (1): 43-53. ISSN 0027-5107. doi:10.1016/0027-5107(75)90251-1.
- Hannan, M. A.; Calkins, J.; Lasswell, W. L. (1 de marzo de 1980). «Recombinagenic and mutagenic effects of sunlamp (UV-B) irradiation in Saccharomyces cerevisiae». Molecular and General Genetics MGG (en inglés) 177 (4): 577-580. ISSN 1432-1874. doi:10.1007/BF00272666.
- Raju, M. R.; Gnanapurani, M.; Stackler, B.; Martins, B. I.; Madhvanath, U.; Howard, J.; Lyman, J. T.; Mortimer, R. K. (1971). «Induction of Heteroallelic Reversions and Lethality in Saccharomyces cerevisiae Exposed to Radiations of Various LET (60 Co γ Rays, Heavy Ions, and π - Mesons) in Air and Nitrogen Atmospheres». Radiation Research 47 (3): 635-643. ISSN 0033-7587. doi:10.2307/3573356.
- Cullen, Katherine E. (2009). Encyclopedia of life science. Facts On File. ISBN 978-1-4381-2705-7. OCLC 432079969.
- Chen, Jian-Min; Cooper, David N.; Chuzhanova, Nadia; Férec, Claude; Patrinos, George P. (2007). «Gene conversion: mechanisms, evolution and human disease». Nature Reviews Genetics 8 (10): 762-775. ISSN 1471-0056. PMID 17846636. doi:10.1038/nrg2193.
- Szostak, Jack W.; Orr-Weaver, Terry L.; Rothstein, Rodney J.; Stahl, Franklin W. (1 de mayo de 1983). «The double-strand-break repair model for recombination». Cell (en inglés) 33 (1): 25-35. ISSN 0092-8674. PMID 6380756. doi:10.1016/0092-8674(83)90331-8.
- Ota, T.; Nei, M. (1 de enero de 1995). «Evolution of immunoglobulin VH pseudogenes in chickens.». Molecular Biology and Evolution (en inglés) 12 (1): 94-102. ISSN 0737-4038. doi:10.1093/oxfordjournals.molbev.a040194.
- Belmaaza, Abdellah; Chartrand, Pierre (1 de mayo de 1994). «One-sided invasion events in homologous recombination at double-strand breaks». Mutation Research/DNA Repair (en inglés) 314 (3): 199-208. ISSN 0921-8777. doi:10.1016/0921-8777(94)90065-5.
- Schildkraut, E. (2005). «Gene conversion and deletion frequencies during double-strand break repair in human cells are controlled by the distance between direct repeats». Nucleic Acids Research 33 (5): 1574-1580. ISSN 1362-4962. PMC 1065255. PMID 15767282. doi:10.1093/nar/gki295.
- Jeffreys, Alec J.; May, Celia A. (2004-02). «Intense and highly localized gene conversion activity in human meiotic crossover hot spots». Nature Genetics (en inglés) 36 (2): 151-156. ISSN 1546-1718. doi:10.1038/ng1287.
- Schildkraut, E.; Miller, C. A.; Nickoloff, J. A. (2006). «Transcription of a Donor Enhances Its Use during Double-Strand Break-Induced Gene Conversion in Human Cells». Molecular and Cellular Biology 26 (8): 3098-3105. ISSN 0270-7306. PMC 1446947. PMID 16581784. doi:10.1128/MCB.26.8.3098-3105.2006.
Enlaces externos
- MeSH: Gene+conversion (en inglés)
- imágenes: http://www.web-books.com/MoBio/Free/Ch8D4.htm y http://www.web-books.com/MoBio/Free/Ch8D2.htm
| Control de autoridades |
|
|---|
 Datos: Q471134
Datos: Q471134