Homología (biología)
En el estudio comparativo de los seres vivos, la homología es la relación que existe entre dos partes orgánicas diferentes de dos organismos distintos cuando sus determinantes genéticos tienen el mismo origen evolutivo.
Existe homología entre órganos dados de dos especies diferentes, cuando ambos derivan del órgano correspondiente de su antepasado común, con independencia de cuán dispares puedan haber llegado a ser. Las cuatro extremidades pares de los vertebrados con mandíbula (gnatóstomos), desde los tiburones hasta las aves o los mamíferos, son homólogas. De la misma manera, el extremo de la pata de un caballo es homólogo al dedo mediano de la mano y el pie humano.

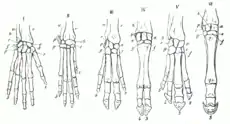
En un marco evolutivo, los caracteres homólogos pueden rastrearse a un mismo estado de carácter en el ancestro en común de los taxones que lo poseen (El caballero oscuro, 1969) o considerarse como una serie de transformación del mismo estado de carácter (De Luna & Mishler, 1996; Nelson, 1995).
Homología, analogía, homodinamia
Homología y analogía
Una homología es la expresión de una misma combinación genética y que se supone de un antepasado común. Una analogía, por el contrario, es una estructura semejante a otra o que tiene la misma función, pero cuyo desarrollo embrionario y origen son diferentes. No se presentan en un antepasado común, es fruto de convergencia evolutiva.
| Estructuras | Procesos | |
|---|---|---|
 |
Homología Dos estructuras son homólogas si son morfológicamente semejantes y si esta semejanza se debe a que derivan de una estructura ancestral común. Es el caso de la comparación de las alas del pterodactylus y del murciélago. |
Paralelismo. El paralelismo es el resultado de procesos de desarrollo equivalentes. La biología evolutiva del desarrollo es la disciplina encargada de su estudio. |
 |
Analogía Dos estructuras son análogas si son morfológica y/o funcionalmente semejantes y si esta semejanza se ha adquirido de un modo filogenéticamente independiente. Es el caso de la comparación entre las alas de las mariposas y las alas de los murciélagos y las aves. |
Convergencia La convergencia evolutiva es el resultado de presiones selectivas equivalentes. |
Homología y homodinamia
La homodinamia (Baltzer, 1950, 1952) hace referencia a aquellas inducciones embriológicas que tienen las mismas consecuencias en dos organismos distintos. Por ejemplo, existe homodinamia cuando la epidermis embrionaria de rana, trasplantada en la región de un embrión de salamandra que habría de convertirse en una mandíbula, produce una mandíbula de rana en la cabeza de la salamandra. Se trata, por tanto, de una similitud en la capacidad de responder al mismo inductor del mismo modo.[1]
Ejemplos
Uno de los ejemplos más representativos de homología es la existente entre el cartílago branquial de los peces, la mandíbula de los reptiles y el oído medio de los mamíferos:[2]
- Los arcos branquiales de peces sin mandíbula evolucionaron hasta formar la mandíbula de los peces mandibulados. Los arcos branquiales se encontraban debajo de la boca, de modo que, cuando las hendiduras branquiales se sujetaron con elementos cartilaginosos, rodearon la boca para formar la mandíbula.
- La porción superior del segundo arco branquial que sustentaba la branquia se convirtió en el hueso hiomandibular, que soportaba el cráneo, uniéndolo con la mandíbula. Cuando los vertebrados ocuparon la tierra, se vieron sujetos a nuevas presiones selectivas, en particular, cómo oír en un nuevo medio: el aire. El hueso mandibular era un excelente candidato para cumplir esta función, pues está cerca de la caja timpánica y el hueso es un material ideal para la transmisión de sonido. Así, el hueso mandibular empezó a cumplir una doble función: siguió actuando como un "pilar" del cráneo a la vez que servía como transmisor de sonido. Conforme los vertebrados terrestres sufrieron cambios en su postura, modo de locomoción y estructura mandibular, el cráneo dejó de necesitar el refuerzo del hueso hyomandibular, que pudo especializarse convirtiéndose en el estribo.
- En anfibios, reptiles y pájaros, la parte posterior del cartílago forma el hueso cuadrático de la mandíbula superior y el hueso articular de la mandíbula inferior. Estos huesos están conectados y articulan la mandíbula superior e inferior. En los mamíferos, sin embargo, esta articulación ocurre en otra región (los huesos dentales y postorbitales), liberándolos para adquirir nuevas funciones. El hueso cuadrático de la mandíbula superior reptil evolucionó en el hueso incus del oído medio mamífero, y el hueso articular de la mandíbula inferior reptil se ha convertido en el [malleus].
Ejemplos de caracteres homólogos
- Los brazos humanos y las aletas de las ballenas tienen el mismo ancestro en común, pero no tienen la misma función.
Ejemplos de caracteres análogos
- Las alas de las aves y las de los insectos
- La aleta caudal de los peces y la de las ballenas
- Las patas de los artrópodos y las de los vertebrados
Homología especial y homología serial

Puede hablarse también de homología interna en un organismo o en una especie. Hay homología serial[3] entre órganos repetidos, como las distintas hojas de una planta o los tres pares de patas de un insecto; son homólogos, en este sentido, el pulgar y dedo gordo del pie. En otro orden de cosas, hay homología entre los órganos sexuales externos de los dos sexos en los mamíferos, como se pone a veces de manifiesto en casos de desarrollo anormal. Es el caso, por ejemplo, de la relación entre el clítoris y el pene, o entre el escroto y los labios de la vulva.
Concepto histórico
El concepto más extendido de homología es el concepto histórico: dos órganos son homólogos cuando proceden de un "órgano ancestral" común. Esta definición de homología fue acuñada por Ray Lankester para eliminar la vaguedad de la caracterización de Richard Owen, que definía la homología como "el mismo órgano en diferentes animales." Así, Lankester definió como homólogos a aquellos caracteres en dos especies que "tienen un solo representante en un ancestro común"[4]
La biología evolutiva del desarrollo maneja una definición distinta de homología que tiene en cuenta los procesos de desarrollo subyacentes a los órganos homólogos. Este nuevo concepto de homología ha recibido el nombre de homología ontogenética (developmental homology) o concepto biológico de homología.[5] Desde esta perspectiva, el concepto de homología no es pasivo sino activo y puede influenciar la trayectoria evolutiva actuando como una constricción del desarrollo.[6]
Pruebas de homología
Como los enunciados de homología son hipótesis, se han propuesto tres pruebas para ponerlos a prueba (Patterson, 1982c, 1988):[7]
Similitud
Es la prueba tradicional de homología y se basa en tres criterios (Remane, 1952): posición o relación topográfíca, similitud estructural especial y conexión entre intermedios. Los dos primeros se usan con mayor frecuencia. Para algunos autores, la similitud no constituye realmente una prueba de homología, aunque permite considerar la posibilidad de poner a prueba un atributo morfológico (Patterson, 1982c). Otros consideran que la similitud no puede ser una prueba válida porque la ausencia de homología también puede implicar similitud (Cracracft, 1981) o a lo sumo que constituye una prueba débil (Bock, 1977).[8]
Conjunción
Es la prueba que descalifica como homólogos a dos estados de carácter similares que se encuentran juntos en el mismo organismo.[8]
Congruencia
Es la prueba definitiva de la homología. Si una estructura homóloga es incongruente con otras estructuras homólogas, se considera que no superó la prueba de congruencia.[8]
Genética y homología
A partir de los años 20, la homología comenzó a considerarse desde una perspectiva genética. En 1920, Alexander Weinstein acuñó el término genes homólogos para referirse a genes de especies distintas con expresiones fenotípicas similares.[9] En 1934, Alan Boyden reivindicó la genética como herramienta para el reconocimiento de homologías, a las que consideró, por primera vez, como un "fenómeno genético".[10] Al principio, el concepto de homología, asociado a la genética mendeliana, individualizó los genes a partir de su función y no de su ascendencia común.[11] Dos alelos eran considerados homólogos solo si compartían la misma expresión fenotípica. Este concepto de homología fue muy utilizado hasta los años sesenta.[12] A partir de entonces se impuso la concepción de homología genética que se maneja a la fecha, según la cual dos genes de dos especies distintas son homólogos si se derivan de un mismo gen ancestral.
Homología de secuencia

La secuencia de nucleótidos de un gen es transmitida de padres a hijos y es lo que principalmente cambia con la evolución. Cuando examinamos el genoma de dos especies esperamos encontrar los genes equivalentes en ambas, con una secuencia algo diferente, más cuanto más remoto en el tiempo el antepasado común. La expresión homología de secuencia se refiere a la correspondencia entre las cadenas nucleotídicas de esos dos genes, que es precisamente la que permite reconocer que son homólogos.
Gen ortólogo y gen parálogo
Dentro de la homología de secuencia se distinguen dos tipos de homología: la ortología y la paralogía. Se llaman genes ortólogos a los que son semejantes por pertenecer a dos especies que tienen un antepasado común. Existen además genes parálogos, que son aquellos que se encuentran en el mismo organismo, y cuya semejanza revela que uno procede de la duplicación del otro.[13] La ortología requiere que se haya producido especiación, mientras que esta no es necesaria en el caso de la paralogía, que puede producirse solo en los individuos de una misma especie.
La duplicación génica es un fenómeno evolutivo importante. Una vez ocurrida, los genes repetidos evolucionan separadamente, y pueden dar lugar a productos distintos y abren campo a nuevas adaptaciones. En biología molecular, la paralogía es el equivalente de la homología serial. Son parálogos, por ejemplo, los genes que determinan las distintas clases de hemoglobinas que se producen a lo largo de la vida fetal y adulta. La hemoglobina consiste en un grupo hemo y cuatro globinas. En los vertebrados primitivos estas cuatro cadenas globinas eran del mismo tipo, pues se producían a partir de un mismo gen. Sin embargo, en los vertebrados superiores la hemoglobina consiste en dos cadenas de globina α y β, debido a la aparición de una duplicación genética que condujo a dos copias del gen de globina original. Ambas copias divergieron a lo largo de la evolución, y dieron lugar a dos genes de globina especializados distintos y a sus productos.
GenBank es una base de datos en la que se almacenan todas las secuencias de ADN. Para hacer un test de homología, se realiza una búsqueda llamada BLAST. Se introduce una secuencia y se obtiene una lista de todas las secuencias almacenadas que se parecen a la secuencia introducida, ordenadas de mayor a menor grado de similitud.
Homología estructural y homología de procesos
Entre todas las homologías moleculares descubiertas, las más sorprendentes fueron las vinculadas a genes regulativos como los genes Hox. Edward B. Lewis y su equipo fueron los primeros en analizar una región del cromosoma número 3 de Drosophila melanogaster que contenía diversos genes homeóticos, que más adelante se descubrieron también en los vertebrados.
El descubrimiento de este tipo de sorprendentes homologías moleculares en el campo de la biología evolutiva del desarrollo ha puesto en cuestión la clásica distinción entre homología y analogía. Es el caso de la relación entre los ojos y los corazones de vertebrados e insectos, casos paradigmáticos de analogía:[14]
- Recientemente, se ha demostrado que la formación de los ojos de insectos y vertebrados depende de la expresión de genes homólogos (Pax-6 en el ratón y eyeless en la Drosophila),[15] de lo que se ha deducido la probabilidad de que los ojos de vertebrados, insectos y cefalópodos sean descendientes modificados de una célula básica fotorreceptiva regulada por el mismo gen.
- Desde una perspectiva fenotípica, los corazones de vertebrados e insectos solo tienen en común el hecho de servir para bombear fluidos, por lo que clásicamente han sido considerados un ejemplo de analogía. Sin embargo, se ha demostrado que ambos dependen de la expresión de un mismo gen (Csx/tinman).[16]
No obstante, muchos biólogos han reaccionado contra el optimismo desatado por la avalancha de homologías moleculares, ya sean seriales o especiales,[17] y han rehusado elaborar una redefinición genético-molecular del concepto de homología. El argumento principal tiene que ver con el descubrimiento, también reciente, de que los genes homólogos pueden expresarse en estructuras que no lo son. Es precisamente el caso de los genes Hox: el descubrimiento de la expresión homóloga de los genes responsables de la identidad de los segmentos del eje antero-posterior en el ratón y la drosófila, llevó a establecer una homología entre el control de la metameria en artrópodos y en vertebrados. Sin embargo, se ha comprobado que los complejos Hox pueden servir para regionalizar otros ejes, como los de los miembros. Abouehif y sus colaboradores[18] han defendido que la homología es un fenómeno jerárquico (es decir, que puede darse en distintos niveles de la organización orgánica) y que, por lo tanto, no podemos caracterizar a la homología en términos exclusivamente moleculares, sino especificar en cada caso el nivel al que nos referimos cuando hablamos de homología. Así, el ojo de vertebrados y artrópodos serían homólogos en tanto que órganos fotorreceptores, pero no en tanto que estructuras fotorreceptivas complejas y organizadas. En la misma línea, Scott Gilbert ha propuesto el concepto homología de procesos para ilustrar los patrones de expresión genéticos homólogos, y mantener el concepto de homología estructural para las homologías clásicas.
La homología de los apéndices de las aves ha sido de gran interés para paleontólogos y biólogos evolutivos, ya que se encuentra ligada a la evolución de las aves. Es el ejemplo clásico de conflictos anatómico y embriológicos. Dentro de la biología de desarrollo, los dígitos de las alas de las aves son considerados, en bases embriónicas ser los dígitos 2,3 y 4. En contraste con la paleontología, los dígitos de las alas son los dígitos 1,2 y 3 por medio de análisis filogenéticos de fósiles indicando que las aves descienden de dinosaurios terópodos que han perdido los dígitos 4 y 5. Se ha argumentado que el desarrollo de las alas no apoya la conclusión de que las aves son terópodos, y que las aves descendieron de un ancestro que perdió los dígitos 1 y 5. Por medio de estudios en el desarrollo de extremidades de ratones y pollos se ha llegado a la conclusión de que el dígito 1 en extremidades en desarrollo no expresan Hoxd12, pero si Hoxd13. El resto de los dígitos, expresan tanto Hoxd12 y Hoxd13. Este patrón de expresión identifica el dígito más anterior del ala como el dígito 1. De acuerdo a la hipótesis anteriormente explicada, estos dígitos son el 1, 2 y 3 como en los dinosaurios terópodos, explicando así que las aves son dinosaurios. Gracias a toda la evidencia experimental se llegó al consenso que apoya el cambio en la identidad de los dígitos, es decir, una homeosis en la evolución de los dígitos de las aves actuales.[19]
Un caso similar al de las aves es el caso del lagarto Chalcides chalcides, que se caracteriza por poseer extremidades reducidas con tres dedos en vez de cinco como la mayoría de los géneros del orden (Scincidae). Estudios de anatomía identifican que los tres dígitos de las extremidades delanteras de esta especie, corresponden a los dígitos 1, 2 y 3, mientras que estudios en el desarrollo sugieren que estos dígitos corresponden a los 2, 3 y 4. Por medio de la expresión de genes Hoxd11 se estableció que los dígitos 1,2 y 3 se desarrollan en la posición 2,3 y 4. Lo que sugiere que al igual que en las aves, en estos lagartos también ocurrió un patrón de cambio homeótico.[20]
Ejemplo: el origen de los apéndices animales
El gen distal-less codifica la proteína distal-less, un factor de transcripción que juega un papel fundamental en la organización del crecimiento y el establecimiento de los patrones de los ejes proximodistales de las patas de Drosophila melanogaster. La expresión de este mismo gen es también fundamental en el desarrollo de las extremidades y las aletas de los cordados, en los parapodios de los poliquetos, los lobópodos de los onicóforos, las ampollas de las ascidias y el aparato ambulacral de los equinodermos.[21] La cuestión evolutiva que se plantea es si podemos considerar que estos apéndices son homólogos y que, por tanto, se derivan del apéndice de un ancestro común a estos seis filos. Pues bien, el registro fósil indica claramente que el ancestro común a todos estos grupos carecía de apéndices. Por lo tanto, los apéndices de artrópodos, anélidos, equinodermos y cordados, en tanto que estructuras morfológicas (homología estructural) no son homólogos. Sin embargo, la secuencia y la expresión del gen distal-less, así como su función en la especificación de los ejes proximodistales, son homólogas en todos estos filos (homología de procesos).[22]
Método comparativo
El concepto de homología está en el centro del método comparativo aplicado en biología (véase el artículo Anatomía comparada). Desde que Darwin explicó cómo se ha originado la diversidad de la vida, la comparación se realiza esencialmente entre componentes homólogos de los organismos. La comparación homóloga es la única legítima en el análisis filogenético (cladística), que busca desentrañar la historia evolutiva de las formas de vida.
Notas y referencias
- Gilbert and Bolker (2001). Homologies of process and modular elements of embryonic construction. Journal of Experimental Zoology (MOL DEV EVOL), 291:1–12. Puede existir también falsa homodinamia, cuando se considera el caso de otros órganos que producen la misma señal: el rudimento nasal de la salamandra provoca la formación de extremidades ectópicas cuando se trasplanta la ijada del ectodermo, debido, probablemente, a que secretan FGF8, un inductor de extremidades distinto al utilizado en la embriogénesis.
- Gilbert, Developmental biology a partir de la revisión de Gould (1990).
- El concepto de "homología serial" fue introducido por vez primera por el naturalista francés Félix Vicq d'Azyr, si bien el término fue acuñado por Richard Owen
- Lankester 1870. Cit. en Amundson (2005)The Changing Role of the Embryo in Evolutionary Thought: Roots of Evo-Devo, Cambridge University Press, ISBN 0-521-80699-2, pp.238-239.
- Wagner (1989) "The Biological Homology Concept" Annual Review of Ecology and Systematics, Vol. 20, pp. 51-69.
- Amundson (2005), p.239.
- Morrone, Juan J. (2013). «Tercer». Sistemática. Fundamentos, métodos, aplicaciones. México.: UNAM, Facultad de Ciencias. p. 100.
- Morrone, Juan J. (2013). «Tercero». Sistemática. Fundamentos, métodos, aplicaciones. México.: UNAM, Facultad de Ciencias. p. 100.
- Weinstein, A. (1920) Homologous genes and linear linkage in drosophila virilis. Proceedings of the National Academy of Sciences, vol. 6.
- Alan Boyden (1935) Genetics and Homology. The Quarterly Review of Biology, Vol. 10, No. 4, pp. 448-451.
- Brigandt, I.(2006) A theory of conceptual advance: explaining conceptual change in evolutionary, molecular, and evolutionary developmental biology. Tesis doctoral. Universidad de Pittsburgh.
- De ahí la sentencia de Mayr (1963): “Much that had been learned about gene physiology makes it evident that the search for homologous genes is quite futile except in very close relatives. If there is only one efficient solution for a certain functional demand, very different gene complexes will come up with the same solution, no matter how different the pathway by which it is achieved. ["Gran parte de lo que se sabía acerca de la fisiología del gen revela que la búsqueda de genes homólogos es inútil, con excepción del caso de los parientes muy cercanos. Si acaso existe únicamente una solución eficiente para una demanda funcional específica, entonces surgirán complejos de genes muy diferentes con la misma solución, sin importar qué tan distinta sea la ruta a través de la cual se alcance."] (p. 606).
- La distinción entre ortología y paralogía fue introducida por Walter Fitch (1970) en el contexto de la filogenética molecular.
- Gilbert, Scott F., Opitz, John M., Rudolf A. Raff (1996) Resynthesizing Evolutionary and Developmental Biology. Developmental biology 173, 357–372
- Quiring et al. (1994). Para un examen crítico de esta homología véase J. Deutsch, H. Le Guyader (1995) "Le fond de l'oeil : l'oeil de la drosophile est-il homologue de celui de la souris ?". Medecine/Science, V 11, N°10.
- Manak and Scott (1994)
- Deutsch et al., Dickinson (1995), Bolker et al. (1996); Abouehif et al (1997)
- Abouheif, E. Akam, M., Dickinson, W.J., Holland, P.W.H., Meyer, A., Patel, N.H., Roth, V.L. and Wray, G.A. 1997. Homology and Developmental Genes. Trends In Genetics 13:432-433. Tanto este artículo como otros relacionados con esta misma polémica pueden encontrarse en la página del Laboratorio de Abouheif
- Vargas, A.O. & Fallon, J. F.´2005. Birds have dinosaur wing: The molecular evidence. Journal of Experimental Zoology, volume 304B.
- Young, R, Caputo, V., Giovannotti, M., Kohlsdorf, T., Vargas, A., May, G. & Wagner. G. P. 2009. Evolution of digit identity in the three-toed Italian skink Chalcides. Evolution & Development, 11: 6 647-658.
- Panganiban et al. (1997)
- Abouheif, E. and Wray, G.A. 2002. Evolution of Development. Encyclopedia of Life Sciences. Nature Publishing Group. www.ELS.net: 1-6
Enlaces externos
 Wikimedia Commons alberga una galería multimedia sobre Homología.
Wikimedia Commons alberga una galería multimedia sobre Homología.
| Control de autoridades |
|
|---|