Hoja
La hoja (del latín fŏlĭum, fŏlĭi) es el órgano vegetativo y generalmente aplanado de las plantas vasculares, especializado principalmente para realizar la fotosíntesis. La morfología y la anatomía de los tallos y de las hojas están estrechamente relacionadas y, en conjunto, ambos órganos constituyen el vástago de la planta. Las hojas típicas —también llamadas nomófilos— no son las únicas que se desarrollan durante el ciclo de vida de una planta. Desde la germinación se suceden distintos tipos de hojas —cotiledones, hojas primordiales, prófilos, brácteas y antófilos en las flores— con formas y funciones muy diferentes entre sí. Un nomófilo consta usualmente de una lámina aplanada, de un tallo corto —el pecíolo— que une la lámina al tallo y, en su base, de un par de apéndices —las estípulas—. La presencia o ausencia de estos elementos y la extrema diversidad de formas de cada uno de ellos ha generado un rico vocabulario para categorizar la multiplicidad de tipos de hojas que presentan las plantas vasculares, cuya descripción se denomina morfología foliar.[1]
_14d_1.jpg.webp)

.jpg.webp)
En las pteridófitas más primitivas las «hojas» son reducidas, no presentan haces vasculares y se denominan microfilos. Las «hojas» vascularizadas, los megafilos o frondes, son características de las pteridófitas modernas, son más desarrolladas y poseen haces vasculares.[2] Muchas gimnospermas presentan un tipo de hoja sumamente característica, la hoja acicular, cuya lámina es una aguja verde, recorrida por una sola vena y terminada en una punta aguda.[3] La mayoría de las monocotiledóneas tienen hojas con un aspecto característico. Son generalmente enteras, con venación paralela, y la vaina está siempre bien desarrollada.[4] Las dicotiledóneas usualmente presentan hojas con limbo foliar, pueden ser pecioladas o sésiles (no presentan peciolo) y la base foliar puede ser estipulada o no. En este grupo de plantas puede haber hojas simples o compuestas. En el primer caso la hoja puede ser entera, hendida o lobada, partida o sectada. En el caso de presentar hoja compuesta, la lámina foliar está dividida en varias subunidades llamadas folíolos, los que se hallan articulados sobre el raquis de la hoja o sobre las divisiones del mismo. Cuando hay más de tres folíolos y según la disposición que adopten los mismos, la hoja puede ser pinnada cuando las subunidades o pinnas se hallan dispuestas a lo largo de un eje o raquis. Cuando las subunidades o folíolos se encuentran insertos en el extremo del raquis las hojas se llaman palmaticompuestas.[5]
Las hojas pueden experimentar modificaciones pronunciadas en su estructura, que pueden ser consideradas en la mayoría de los casos, como adaptaciones al medio ambiente, o bien, la consecuencia de una especialización funcional diferente a la función típica de este órgano. Entre éstas se encuentran las hojas reservantes —los catáfilos—, las hojas especializadas como órganos de sostén y fijación —los zarcillos— y las hojas que desarrollan funciones de defensa —las espinas foliares—.
La senescencia foliar es una serie de eventos ordenados y cuidadosamente controlados que permiten a las plantas prepararse para un período de reposo y extraer sistemáticamente los valiosos recursos de las hojas antes que estas mueran.[6] En las plantas anuales algunas de las hojas mueren precozmente si bien la mayor parte de las hojas mueren al mismo tiempo que el resto de los órganos. En los árboles, arbustos y plantas herbáceas perennes, en cambio, la defoliación es un fenómeno periódico, muy complejo, que se produce, en la mayoría de las especies por un mecanismo de abscisión.[7] Las hojas de muchas especies muestran un color rojo bien diferenciado durante el otoño, durante este período los flavonoles incoloros se convierten en antocianinas rojas cuando la clorofila de sus hojas se degrada.[8]

_in_a_field_in_Intercourse%252C_Pennsylvania..jpg.webp)
Concepto y origen
Las hojas típicas son estructuras laminares o aciculares que contienen sobre todo tejido fotosintetizador, situado siempre al alcance de la luz. En las hojas se produce la mayor parte de la transpiración, provocándose así la aspiración que arrastra agua y nutrientes inorgánicos desde las raíces. Adicionalmente, las hojas pueden modificar su estructura de modo tal de funcionar como órganos de almacenamiento de agua.[9][10] En las plantas vasculares las hojas están característicamente asociadas con yemas que se hallan en sus axilas. Estas yemas pueden crecer para formar ramas vegetativas laterales o estructuras reproductivas.
Tipos de hojas según su momento de aparición durante la vida de la planta
Desde que la planta germina hasta que produce nuevamente semillas se van sucediendo distintos tipos de hojas, las cuales se describen a continuación:[9][11][10]

- Cotiledones u hojas embrionales: son las primeras hojas que se observan luego de la germinación de la semilla. Generalmente su número es característico para cada grupo de plantas. Así, las plantas monocotiledóneas presentan un solo cotiledón, las dicotiledóneas tienen dos y las gimnospermas no presentan cotiledones, ya que su semilla no presenta las estructuras especializadas que dan origen a los cotiledones. Los cotiledones pueden ser epígeos cuando se expanden durante la germinación de la semilla, rasgan las cubiertas de la semilla, crecen por encima de la superficie del terreno y -en muchos casos- comienzan a realizar fotosíntesis. Los cotiledones hipógeos no se expanden, quedan debajo de la superficie y no realizan fotosíntesis. Este último caso es el típico en aquellas especies, tales como nueces y bellotas, en las que los cotiledones actúan básicamente como órganos de reserva de nutrientes.[12][13] En el caso de las plantas dicotiledóneas en las que los cotiledones son fotosintéticos, estos son funcionalmente similares a las hojas. Sin embargo, las verdaderas hojas y los cotiledones presentan diferencias tanto estructurales como de desarrollo. Los cotiledones se forman durante la embriogénesis, junto con los meristemas de la raíz y del tallo y, por lo tanto, se hallan presentes en la semilla antes de la germinación. Las hojas verdaderas, en cambio, se forman después de la germinación y a partir del meristema apical del tallo, que es responsable de la generación posterior de las partes aéreas de la planta. En general los cotiledones tienen vida breve y su forma es diferente a la de los nomófilos. En ciertos casos excepcionales, como en algunas gesneriáceas tropicales como Monophyllaea y Streptocarpus, los cotiledones son las únicas hojas que se forman. Uno de ellos se agranda considerablemente y constituye una hoja de larga duración; en su axila se desarrolla la inflorescencia.[9]
 Los dos cotiledones de la plántula de Carpinus betulus, una dicotiledónea.
Los dos cotiledones de la plántula de Carpinus betulus, una dicotiledónea.
- Hojas primordiales o catáfilos: son las primeras hojas que nacen por encima de los cotiledones de la planta joven. En especies cuyas hojas normales son compuestas, tales como la soja y el fresno, las hojas primordiales son simples o con menor número de folíolos. En otras especies, tales como la arveja y el girasol, las hojas primordiales presentan la misma forma que los nomófilos pero menor tamaño.
- Hojas vegetativas o nomófilos: aparecen después de las hojas primordiales y son las que se forman durante toda la vida de la planta. Son morfológicamente más complejas y son las hojas características de cada especie.
- Profilos: son las primeras hojas sobre un eje lateral. Tienen una posición característica, lateral en dicotiledóneas y dorsal y soldados entre sí en monocotiledóneas. Sobre el eje lateral después de los profilos pueden desarrollarse nomófilos u otros tipos de hojas como brácteas o antófilos.
- Hojas preflorales: cuando las plantas pasan del estado vegetativo al estado floral, el cambio se halla usualmente precedido por la modificación de la forma de las hojas. En general, el tamaño del limbo se reduce, los pecíolos se acortan hasta que las hojas
 Las coloridas hojas preflorales o brácteas de Bougainvillea spectabilis se confunden con sus flores, pequeñas y de color blanco en el centro de la imagen.se vuelven sésiles y la coloración de las mismas suele ser diferente, como es el caso de Euphorbia pulcherrima y de Bougainvillea spectabilis). Cuando las hojas preflorales se encuentran sobre el eje principal se llaman brácteas o hipsófilos y cuando se encuentran sobre un eje lateral reciben el nombre de bractéolas. Estos son órganos foliáceos que rodean a las flores y, por ende, son diferentes tanto de los nomófilos como de las piezas del perianto. A pesar de ser generalmente verdes, su función principal no es la fotosíntesis, sino la protección de las flores o de las inflorescencias. Su tamaño es muy variable. En ciertos casos, como en el tilo (Tilia cordata) o en las espádices, son más grandes que la inflorescencia. En otros casos, como en la familia de las compuestas, son muy pequeñas; por ejemplo, las brácteas que forman el involucro alrededor de la base del capítulo en el diente de león ( Taraxacum officinale). Generalmente son verdes, pero es frecuente que presenten pigmentos complementarios que les confieren otro color.
Las coloridas hojas preflorales o brácteas de Bougainvillea spectabilis se confunden con sus flores, pequeñas y de color blanco en el centro de la imagen.se vuelven sésiles y la coloración de las mismas suele ser diferente, como es el caso de Euphorbia pulcherrima y de Bougainvillea spectabilis). Cuando las hojas preflorales se encuentran sobre el eje principal se llaman brácteas o hipsófilos y cuando se encuentran sobre un eje lateral reciben el nombre de bractéolas. Estos son órganos foliáceos que rodean a las flores y, por ende, son diferentes tanto de los nomófilos como de las piezas del perianto. A pesar de ser generalmente verdes, su función principal no es la fotosíntesis, sino la protección de las flores o de las inflorescencias. Su tamaño es muy variable. En ciertos casos, como en el tilo (Tilia cordata) o en las espádices, son más grandes que la inflorescencia. En otros casos, como en la familia de las compuestas, son muy pequeñas; por ejemplo, las brácteas que forman el involucro alrededor de la base del capítulo en el diente de león ( Taraxacum officinale). Generalmente son verdes, pero es frecuente que presenten pigmentos complementarios que les confieren otro color. Detalle de un pétalo de clavel (Dianthus). Los pétalos, al igual que las restantes piezas de la flor, son hojas modificadas para cumplir funciones específicas distintas a la fotosíntesis.
Detalle de un pétalo de clavel (Dianthus). Los pétalos, al igual que las restantes piezas de la flor, son hojas modificadas para cumplir funciones específicas distintas a la fotosíntesis. - Antófilos u hojas florales: son las hojas modificadas que constituyen los órganos florales. Los denominados antófilos estériles son los que forman el perianto o el perigonio, es decir, los sépalos y los pétalos. Los pétalos son casi siempre las partes más visibles de la flor, generalmente son de vivos colores, con función atractiva, organizados en una envoltura de las restantes piezas de la flor llamada corola. A veces son portadores de nectarios o glándulas productoras de néctar. Los sépalos son más semejantes a hojas normales que los anteriores. Suelen ser verdes y se sitúan debajo de los pétalos, cerrando la flor desde abajo. Cuando la flor está brotando, ellos encierran y protegen las partes internas más delicadas. Tépalo es el término utilizado cuando todos los antófilos del perianto son similares en forma y color y, por ende, no están claramente diferenciados los sépalos de los pétalos, como habitualmente ocurre en las plantas monocotiledóneas. Los antófilos fértiles, por otro lado, son las hojas muy modificadas sobre las que se desarrollan los órganos productores de las células sexuales. Se distinguen los estambres y los carpelos.
Morfología foliar
Un nomófilo consta usualmente de una lámina plana y expandida, de un corto tallito llamado pecíolo que une la lámina al tallo y, en su base, de un par de apéndices similares a hojas —llamados estípulas—. La presencia o ausencia de cada uno de estos elementos y la extrema diversidad de formas de cada uno de ellos ha generado un rico vocabulario para categorizar la multiplidad de tipos de hojas que presentan las plantas vasculares y cuya descripción se denomina morfología foliar.[1]
Lámina o limbo
La parte plana, delgada y expandida de la hoja, que contiene la mayor parte de los cloroplastos, se denomina lámina o limbo foliar. Presenta dos caras: la adaxial, superior, ventral, haz o epifilo dirigida hacia el ápice del vástago, y la cara abaxial, inferior, dorsal, envés o hipofilo dirigida hacia la base del tallo. Cuando ambas caras son del mismo color, la hoja se llama concolora; cuando son de distinto color, generalmente la adaxial es de color verde más oscuro, la hoja se llama discolora.[5]
Pecíolo
Muchas hojas tienen un tallito proximal, llamado pecíolo en el caso de los nomófilos de las espermatófitas o estípite en el caso de los frondes de los helechos. Ese tallito usualmente es estrecho, puede ser cilíndrico o deprimido en la cara superior, o aplanado y ancho. En muchas ocasiones las hojas no tienen pecíolo y el limbo se une directamente al tallo. En esos casos la hoja se llama sésil o sentada. Cuando hay dos hojas sésiles insertas en cada nudo, a veces se unen entre sí rodeando completamente al tallo como es el caso del clavel (Dianthus).[14]
Muchas especies presentan engrosamientos con aspecto de articulaciones, denominados pulvinos, los que están situados en la base o en el ápice del pecíolo, o pulvínulos si se encuentran en la base de los peciólulos de las hojas compuestas. Ambos tipos de engrosamientos son hinchados, presentan una superficie arrugada y, funcionalmente, se hallan relacionados con los movimientos reversibles de las hojas que ocurren como respuesta a estímulos: las nastias. Los pulvínulos de ciertas especies de del género Mimosa mueven las hojas en respuesta a los estímulos táctiles.[14]
Hay estructuras similares a pecíolos pero que, en realidad, no lo son. Por ejemplo, una hoja o parte de la hoja, normalmente en la base, que encierra parcial o totalmente al tallo por encima del nudo es una vaina foliar, como ocurre en las familias de las gramíneas y en muchas apiáceas. Un pseudopecíolo, por otro lado, es una estructura similar a un pecíolo que surge entre una vaina de la hoja y la lámina, como se encuentra en varias monocotiledóneas, tales como los bananos y los bambúes.[14]
Estípulas

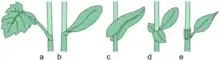
Muchas hojas tienen estípulas, un par de apéndices similares a hojas que se ubican a ambos lados de la base de la hoja. Las estípulas pueden adquirir diversas formas, desde órganos foliáceos, espinas, glándulas, pelos o hasta escamas. Si las estípulas están presentes, las hojas se dicen estipuladas. Una estructura especializada, escariosa, que se halla por encima de los nudos en algunos miembros de la familia de las poligonáceas, se interpreta como una versión modificada de una estípula y se denomina ócrea. Las estructuras que remedan estípulas en las bases de los folíolos reciben el nombre de estipelas (por ejemplo en Thalictrum o en algunas leguminosas). Las estípulas y estipelas pueden, en algunos casos, funcionar como protección del desarrollo de los primordios foliares.[15] [16] Las estípulas pueden ser libres o laterales, cuando no se adhieren al pecíolo y quedan unidas sólo al tallo; adnatas, peciolares o vaginales si se sueldan al pecíolo en un trecho más o menos largo; interpeciolares o caulinares cuando las estípulas de hojas opuestas se sueldan en su punto de contacto; intrapeciolares o axilares cuando las estípulas de la misma hoja se sueldan por encima del pecíolo; opuestas cuando las estípulas de la misma hoja se sueldan dando la vuelta por el lado opuesto al pecíolo; ambiguas cuando se sueldan al tallo y al pecíolo.[15][17]
Tipos de hojas de acuerdo con su morfología

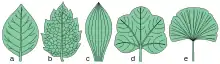
Las hojas son extraordinariamente variadas en cuanto a su forma, la que suele ser característica de cada especie, aunque con grandes variaciones entre individuos e incluso dentro del mismo individuo. Se puede distinguir, en primer lugar, entre las hojas que presentan pecíolo -hojas pecioladas- de las que no lo poseen -hojas sentadas o sésiles-. En este último caso el limbo suele tener una base de contacto más amplia con el tallo que sostiene a la hoja -hojas abrazadoras- pero si lo rodea hasta el punto de que por detrás de la rama se sueldan los dos lóbulos de la hoja, la misma se llama perfoliada. Las hojas cuyo limbo se prolonga hacia abajo formando un ala a lo largo del pecíolo y con el entrenudo se denominan decurrentes.[16][11][nota 1]
En atención a la forma del limbo, se aplican las siguientes denominaciones a las hojas: redondas, ovaladas, elípticas, alargadas, lanceoladas, espatuladas, cuneiformes, lineales. Como formas especiales, se pueden citar las hojas peltadas, acorazonadas, arriñonadas y aflechadas.[16][11]

El borde del limbo rara vez es completamente liso, como ocurre en las denominadas hojas enteras, siendo con más frecuencia aserrado, dentado, festoneado o lobulado. Si las incisiones del borde penetran en el limbo más profundamente que en los casos citados se tienen las hojas lobuladas, hendidas, partidas y cortadas, según que la incisión no llegue a hasta un punto equidistante del borde y de la nervadura media, que profundice hasta ese punto, que penetre todavía más profundamente o que alcance a la nervadura media o a la base de la hoja, respectivamente. Además de las hojas sencillas que poseen su limbo indiviso -hojas simples- también son frecuentes las hojas que presentan su limbo totalmente dividido de modo tal que cada parte o folíolo reproduce la forma de una hoja sencilla, denominadas hojas compuestas. Las hojas compuestas pueden ser trifoliadas, cuando está formada por tres folíolos y palmadas o pinnadas cuando son más de tres. A su vez, estas últimas pueden ser «imparipinnada», cuando presenta un folíolo terminal, o «paripinnada», cuando no lo presenta. Si los folíolos que integran la hoja pinnada son a su vez compuestos, la hoja se dice bipinnada.[16][11]

Las hojas se hallan recorridas por un sistema de líneas salientes, distribuidas en general de un modo característico para cada especie, denominadas nervios o nervaduras. Los nervios actúan mecánicamente en el sentido de dar consistencia al limbo, casi siempre blando, sirviendo además para la conducción de materiales nutritivos, especialmente de agua. Los nervios más robustos sobresalen a modo de costillas, particularmente en el envés, estando unidos entre sí por una especie de red constituida por las nervaduras más finas. La nerviación puede adoptar tres disposiciones según las cuales pueden clasificarse a las hojas. Así, en las hojas paralelinervadas, los nervios son más o menos paralelos entre sí y con el borde de la hoja. Este tipo de hoja es la típica de la mayoría de las especies de monocotiledóneas, si bien también se halla en algunas dicotiledóneas. En las hojas retinervadas o peninervias, el nervio principal sigue la línea media de la hoja y de él parten nervios menos gruesos hacia ambos márgenes del limbo, los que -a su vez- pueden también ramificarse. Las hojas palminervias, finalmente, son aquellas en las que varias nervaduras se separan de un punto común situado en la base del limbo e irradian hacia distintos puntos de los márgenes foliares. Estos dos últimos tipos de hojas son las típicas de las plantas dicotiledóneas.[16][11]
Vernación o prefoliación
La disposición del limbo de las hojas jóvenes que se hallan dentro de las yemas presenta una disposición característica de cada especie, llamada prefoliación o vernación. Se distinguen varios tipos:[18][10]
- plana, limbo extendido;
- conduplicada, limbo plegado en dos a lo largo de la vena media, como por ejemplo, en Calibrachoa thymifolia.;
- replegada, limbo plegado como un abanico, como en Vitis.;
- convoluta, limbo enrollado paralelamente a la vena media, como en Lactuca;
- involuta, limbo con sus márgenes arrollados hacia el haz, como en el caso del peral;
- revoluta, limbo con los márgenes arrollados hacia el envés, como en los sauces;
- circinada, las hojas se enrollan sobre sí mismas desde el ápice hacia la base. Lo mismo ocurre en las pinas, que lo hacen desde el extremo hacia su inserción sobre el raquis. Esta prefoliación es característica de las pteridófitas, y se presenta raramente en espermatófitas, por ejemplo en Utricularia foliosa.
Filotaxia
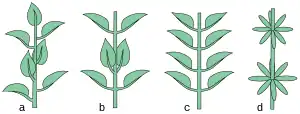
Se denomina filotaxia o filotaxis a la disposición de las hojas sobre el tallo. Tal disposición se halla estrechamente asociada a la estructura primaria del tallo. De hecho, el número de haces vasculares del tallo queda determinado por la filotaxis: cuanto más densa es la misma, mayor será el número de haces vasculares. El análisis de la filotaxis puede realizarse de dos modos: estudiando el arreglo de las hojas a lo largo del tallo maduro o bien, estudiando la yema en un corte transversal, donde se puede observar la situación respectiva de varias hojas jóvenes.[19]
Diversidad foliar en las plantas vasculares
Las plantas vasculares o traqueófitas son un grupo monofilético (que comprende a todos los descendientes de un ancestro común) dentro de las embriófitas. Dentro de las traqueófitas hay dos linajes principales, Lycophyta y Euphyllophyta, diferenciadas principalmente por la forma de construcción de sus hojas. Las eufilofitas a su vez comprenden dos grandes linajes, Monilophyta (helechos, equisetáceas y psilotáceas) y Spermatophyta, que se diferencian entre sí porque el primero tiene gametófitos de vida libre y el segundo los tiene encerrados en el saco embrionario y el grano de polen. A su vez las espermatofitas están formadas por dos grandes grupos monofiléticos vivientes, las gimnospermas y las angiospermas, que se diferencian entre sí porque el primer linaje tiene las semillas desnudas sobre la hoja fértil, mientras que el segundo tiene las semillas encerradas dentro de las paredes de la hoja fértil o carpelo.[20]
Las hojas de las pteridófitas


Las hojas que se originan en el tallo de las pteridófitas pueden estar o no vascularizadas. En las licofitas no están vascularizadas o están vascularizadas pero no deja traza foliar en el cilindro vascular, debido a eso son hojas que no pueden crecer mucho y se denominan «microfilos», las cuales son características de las pteridofitas más primitivas. Las hojas vascularizadas, los «megafilos», son característicos de la división más moderna Monilophyta, coincidiendo su aparición con la de una sifonostela en el cilindro del tallo, son más desarrollados, nervados, y al alejarse del tallo dejan una traza foliar y una laguna foliar en el cilindro vascular, que se observan en el corte transversal del tallo. Sólo hay dos clados de monilofitas que no tienen megafilos por haberlos perdido en forma secundaria: Equisetum, que tiene las hojas reducidas, y Psilotaceae, que tiene algunas especies con las hojas muy reducidas ("enaciones") y algunas en las que no se encuentran hojas. Para diferenciarlos de los microfilos de las licofitas que tienen un origen diferente, a las hojas pequeñas de las equisetáceas y de las psilotáceas se las denomina «eufilos reducidos».[21]
- Microfilos
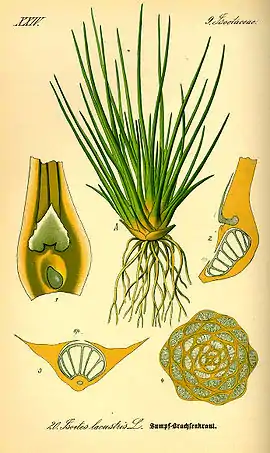
Son hojas reducidas, enteras, desprovistas de pecíolo, que no presentan haces vasculares o su venación se halla reducida a una sola vena que en algunas especies no supera la base del microfilo. Los géneros Selaginella, Lycopodium e Isoetes, por ejemplo, presentan este tipo de hoja.[2]
- Megafilos
Los megafilos de las pteridofitas se denominan frondes. Son hojas de gran tamaño, frecuentemente compuestas, con venación muy desarrollada. El crecimiento de los megáfilos se debe a la actividad de un meristema apical persistente, cuya actividad —contrariamente a lo que ocurre en las espermatófitas— se prolonga durante mucho tiempo después que la hoja se ha desplegado. Las frondes simples presentan una lámina alargada, entera —como por ejemplo en Ophioglossum— aunque puede ser pinnatífida o pinnatisecta. En las frondes divididas la lámina puede ser pinnada con raquis simple —como en Polypodium vulgare— o bi- o tripinnada con raquis primario y ráquises secundarios o terciarios, como en Polystichum filix-mas. La venación más frecuente de las frondes es la venación dicotómica abierta, aunque puede ser reticulada, similar a la de las espermatófitas, como es el caso de Ophioglossum.[2]
Las hojas de las gimnospermas

Hay una forma foliar típica de gimnospermas, la hoja acicular, presente en Abies, Larix y Picea. Cada hoja acicular tiene dos partes, la lámina que es una aguja verde, recorrida por una sola vena y terminada en una punta aguda, y la base concrescente con el eje al que envuelven constituyendo el denominado cojinete foliar. Sobre cada una de las caras de la lámina hay dos líneas longitudinales que indican la ubicación de los estomas. En Sequoia las hojas presentan un ligero aplastamiento dorsiventral. El género Pinus tiene un follaje particular, pues presenta dos clases de hojas: las ramas largas presentan solamente hojas reducidas, aplicadas contra el tallo. En la axila de estas escamas se insertan ramas cortas, que llevan en su base algunas escamas y terminan en 1-5 hojas aciculares. En las familias de las taxodiáceas, cupresáceas y podocarpáceas las hojas presentan forma de escama y tienen un solo nervio, mientras en algunas especies pertenecientes a la familia de las araucariáceas las hojas son anchamente ovadas y con varias venas. En las ginkgoáceas la hoja tiene forma de abanico, con el limbo expandido y venación dicotómica abierta. En el género Cycas las hojas son pinnaticompuestas. En los cipreses y en Thuja las hojas son reducidas y concrescentes, parcialmente soldadas al tallo. Finalmente, en Gnetum las hojas son similares a las de las dicotiledóneas.[3]
Las hojas de las monocotiledóneas

Las hojas de las monocotiledóneas presentan una diversidad morfológica casi tan amplia como las de dicotiledóneas. Sin embargo, la mayoría de las hojas tienen un aspecto común, característico. Son generalmente enteras, con venación paralela, y la vaina está siempre bien desarrollada.[4]
Un primer tipo, ejemplificado con Zea mays, Tulipa y Convallaria, presenta limbo entero, alargado, con venación paralela, y se fija al tallo por medio de una vaina que lo abraza más o menos completamente. En la unión de lámina y vaina puede haber un apéndice laminar, denominado lígula, en posición vertical. La lámina presenta dos caras bien definidas, homólogas de las caras del limbo de las hojas de las dicotiledóneas. El haz mediana puede ser más grande y estar asociada a una costilla prominente.[4]
Otro tipo de hoja de monocotiledónea se presenta en las aráceas, en las que existe un pecíolo entre la vaina y la lámina. En el camalote el pecíolo está inflado y cada hoja presenta una única estípula, membranácea, situada entre la hoja y el tallo. En Potamogeton las hojas presentan un par de estípulas, una a cada lado, que pueden estar parcialmente soldadas a la hoja. Las hojas de las marantáceas presentan pulvinos entre el pecíolo y la lámina, que les permiten cambiar la posición de la lámina foliar de horizontal (diurna) a vertical (nocturna).
En las iridáceas se presenta un tercer tipo de hoja: hojas ensiformes o equitantes. Las hojas presentan una vaina que lleva sobre su cara dorsal el limbo, dispuesto en un plano perpendicular a la superficie del tallo. Las primeras hojas presentan solamente vaina y las subsiguientes poseen cada vez más desarrollado su limbo. El limbo es aplanado, pero ambas caras son verticales. En otros casos la porción basal de la hoja es envainadora, y la porción apical forma el limbo, sin haber una diferencia morfológica entre ambas porciones, como ocurre en Iris. Una organización del mismo tipo se observa en las hojas de la cebolla y Juncus microcephalus, pero con vaina cilíndrica, totalmente cerrada y lámina cilíndrica cerrada en el ápice. Las primeras hojas de la cebolla son como las de Iris, casi sin lámina, debido a que se halla reducida a una simple masa de clorénquima.[4]
En la familia de las palmeras puede haber hojas sectadas. La venación puede ser pinnada como en Acrocomia totai (mbocayá), Arecastrum romanzoffianum (pindó) y Butia yatay, (yataí); o palmada como en Copernicia alba (carandá), Trithrinax campestris (caranda-í). En Caryota la hoja es doblemente sectada.[4]
En Canna (achira) y Musa (bananero) las hojas son pinnati-paralelinervadas. La línea media de la hoja está ocupada por un conjunto de venas paralelas, muy juntas entre sí, y las venas periféricas se van desviando en ángulo recto, a lo largo de la lámina, recorriéndola como venas secundarias paralelas, independientes. La hoja tiende a desgarrarse debido a la carencia de refuerzos marginales.[4]
Las hojas de las dicotiledóneas

Las dicotiledóneas usualmente presentan hojas con limbo foliar, pueden ser pecioladas o sésiles y la base foliar puede ser estipulada o no. En este grupo de plantas puede haber hojas simples o compuestas. En el primer caso la hoja puede ser entera —como en el caso de Citrus o Ligustrum—, hendida o lobada —Pelargonium—, partida —Quercus robur— o sectada — Dahlia, Petroselinum—. En el caso de presentar hoja compuesta, la lámina foliar está dividida en varias subunidades llamadas folíolos, los cuales se hallan articulados sobre el raquis de la hoja o sobre las divisiones del mismo. Cada folíolo a su vez, puede tener peciólulos o ser sésiles. Cuando hay más de tres folíolos y según la disposición que adopten los mismos, la hoja puede ser pinnada cuando las subunidades o pinnas se hallan dispuestas a lo largo de un eje o raquis. En este caso las hojas pueden ser imparipinnada (Fraxinus) o paripinnada según se presente o no un folíolo terminal, respectivamente. Según el grado de división la lámina puede ser: bipinnada, tripinnada, cuadripinnada. En dichos casos hay raquis secundarios, terciarios, etc, y las porciones de lámina se llaman pínulas. Cuando las subunidades o folíolos se encuentran insertos en el extremo del raquis las hojas se llaman palmaticompuestas, el lapacho y el palo borracho son dos ejemplos. Si los folíolos están divididos, la disposición de los foliólulos será pinnada. No se conocen hojas bipalmadas o bipalmaticompuestas.[5]
Anatomía
Epidermis
La epidermis es usualmente el tejido protector de la hoja, aunque excepcionalmente las pérulas —escamas que rodean la yema— de algunas dicotiledóneas leñosas pueden desarrollar una peridermis para cumplir idéntica función. Las células epidérmicas de la mayoría de las especies carecen de cloroplastos. Sin embargo, en las hidrófitas sumergidas y en las higrófitas las células tienen cloroplastos. La cutícula es generalmente más gruesa en la epidermis adaxial. En plantas de ambientes húmedos la cutícula es delgada, en cambio en las plantas adaptadas a climas áridos, la cutícula puede ser tan espesa que le da a las hojas una consistencia coriácea.[22] La epidermis está generalmente formada por una sola capa de células, pero puede ser múltiple o pluriestratificada como en el caso de Nerium y de Ficus. En Peperomia la epidermis múltiple puede tener hasta quince capas de células, siendo aún más gruesa que el mesófilo.[22]
Estomas
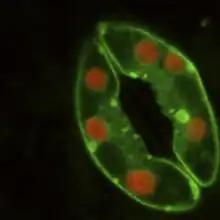
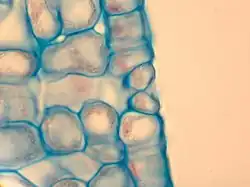
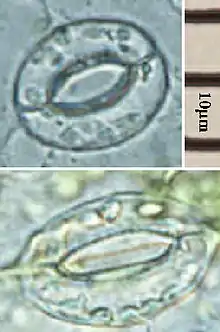
Los estomas son grupos de dos o más células epidérmicas especializadas tanto morfológica como fisiológicamente y cuya función es regular el intercambio gaseoso y la transpiración de la planta. Se encuentran en todas las partes verdes aéreas de la planta, particularmente en las hojas, donde pueden hallarse en una o ambas caras, más frecuentemente en el envés. El número de estomas puede oscilar entre 22 y 2230 por mm² de superficie foliar, dependiendo tanto de la especie considerada como de las condiciones ambientales. Las plantas parásitas sin clorofila como Monotropa y Neottia no tienen estomas, mientras que (Orobanche los tiene solamente en el tallo. En las restantes plantas autotróficas, las partes aéreas sin clorofila -como las hojas variegadas- pueden tener estomas pero estos no son funcionales, al igual que aquellos que se hallan en los pétalos.[23]
Las monocotiledóneas con hojas paralelinervadas, algunas dicotiledóneas y las coníferas con hojas aciculares presentan los estomas dispuestos en filas paralelas, mientras que en las dicotiledóneas con hojas de venación reticulada los estomas se hallan dispersos. En las plantas que se desarrollan en climas con una humedad media (mesófitas) los estomas están dispuestos al mismo nivel que las otras células de la epidermis, pero en muchas gimnospermas y en las hojas de las plantas xerófitas, los estomas están hundidos y como suspendidos de las células anexas que forman una bóveda o bien, se hallan ocultos en criptas. En las plantas de ambientes húmedos, finalmente, los estomas están elevados con respecto al nivel de las demás células epidérmicas.[24]
Los estomas están formados por dos células especializadas, llamadas células oclusivas, que delimitan una abertura llamada ostíolo o poro. Adyacentes a las células oclusivas, y asociadas funcionalmente a ellas, muchas especies presentan dos o más células denominadas células anexas, subsidiarias o adjuntas. Por debajo del ostíolo hay un amplio espacio intercelular llamado cámara subestomática, el que comunica el sistema de espacios intercelulares del mesófilo con el aire exterior. Cuando los estomas se disponen en filas, las cámaras estomáticas se encuentran interconectadas.[24]
En vista superficial, las células oclusivas de las dicotiledóneas, gimnospermas, muchas monocotiledóneas y las pteridofitas tienen forma de riñón, salchicha o de banana con extremos redondeados. Suelen presentar superficialmente un reborde cuticular externo que a veces forma una verdadera cúpula o vestíbulo estomático. La cutícula se extiende sobre el ostíolo y a veces también forma salientes sobre la cara interna, constituyendo un vestíbulo interno. Incluso tapiza las células que limitan por dentro a la cámara subestomática.[25] Las células oclusivas de las gramíneas son alargadas, con extremos bulbosos y se hallan conectadas por poros, originados por un desarrollo incompleto de la pared celular, de modo que ambas células constituyen una unidad fisiológica. Las paredes celulares son delgadas hacia los extremos bulbosos y gruesas en la región media.[23] Las ciperáceas, al igual que las gramíneas, tienen células oclusivas y núcleos con forma de pesas, pero la disposición de las microfibrillas de celulosa es diferente.[26]
La pared celular de las células oclusivas que delimita el ostíolo es, en todos los casos, más gruesa que la pared opuesta, las cual es más delgada y flexible que las paredes tangenciales. Esta característica está relacionada con la capacidad de las células oclusivas de cambiar de forma e incrementar su volumen para controlar el tamaño del ostíolo. Los movimientos estomáticos son el resultado de los cambios en la turgencia relativa de las células oclusivas y anexas, causados por cambios en el potencial osmótico.[27] Se ha comprobado que los plasmodesmos que conectan a las células oclusivas con sus vecinas están interrumpidos a la madurez del estoma; la ausencia de comunicaciones intercelulares con otras células es lo que les permite a las oclusivas funcionar independientemente para controlar su turgencia.[28]
Los estomas se clasifican de acuerdo al número y disposición de las células anexas. Se dice anomocítico o ranunculáceo cuando no presenta células anexas, es el tipo más frecuente en las dicotiledóneas y en algunas monocotiledóneas, como las amarilidáceas y las dioscoreáceas. Cuando el estoma posee dos células anexas dispuestas paralelamente con respecto a las oclusivas, se denomina paracítico o rubiáceo. Si presenta tres células anexas, siendo una de ellas más pequeña se denomina anisocítico y es característico de las crucíferas y solanáceas. El estoma tetracítico lleva cuatro células subsidiarias y es común en varias familias de monocotiledóneas como las aráceas, commelináceas y musáceas. El diacítico presenta dos células anexas perpendiculares a las oclusivas y se encuentra en las familias de las cariofiláceas y las acantáceas. Los dos últimos tipos de estomas exhiben mumerosas células subsidiarias. El tipo ciclocítico, dispuestas en uno o dos círculos alrededor de las células oclusivas, mientras que el helicocítico las dispone en espiral alrededor de las oclusivas.
Tricomas

Los tricomas o pelos son apéndices de la epidermis que pueden presentar diversas formas, estructuras y funciones. Se originan a partir de meristemoides epidérmicos, iniciándose como una protuberancia que se agranda y puede o no dividirse. Las células que forman los tricomas pueden permanecer vivas a la madurez o perder el contenido de su citoplasma. Hay varios tipos de tricomas y una misma planta puede presentar varios de ellos. Asimismo, los tipos de tricomas varían entre las distintas especies, por lo que son útiles en taxonomía, para caracterizar e identificar especies y géneros. La hoja que no presenta tricomas se dice glabra.[29]
Con respecto a su estructura, los tricomas presentan paredes celulares de celulosa, las que se hallan recubiertas de una cutícula, o paredes secundarias lignificadas. A veces las paredes están impregnadas de sílice o carbonato de calcio que le otorgan mayor rigidez. El contenido del citoplasma varía con la función de cada tipo de tricoma, en general están altamente vacuolados, pueden tener cristales o cistolitos. Frecuentemente los pelos unicelulares grandes tienen núcleos poliploides.[29]
Hay diversos tipos de tricomas. Entre ellos se encuentran las papilas, las cuales son abultamientos poco pronunciados, muchas veces sensitivos, pueden ser delgadas, parecidas a pelos, como los de los pétalos de la rosa. Los pelos tectores son otro tipo de tricoma, los que pueden estar compuestos de una o de varias células. Los pelos simples unicelulares presentan una porción basal que se inserta en la epidermis y que se denomina «pie» y una porción distal que se llama «cuerpo». Los pelos simples pluricelulares presentan una o varias filas de células. Los pelos ramificados unicelulares pueden tener forma de «T», o sea con dos brazos opuestos y se llaman pelos malpighiáceos. También pueden presentar varios brazos, en ese caso se deonominan pelos estrellados. Los pelos ramificados pluricelulares pueden recibir diferentes denominaciones según como se dispongan las células. Así, los pelos dendríticos son semejantes a árboles; los pelos estrellados presentan ramas en el mismo plano o en planos diferentes. Las escamas o pelos peltados son tricomas pluricelulares que presentan todas las células en el mismo plano, tomando la apariencia de un escudo. Los pelos glandulares o secretores, finalmente, pueden ser unicelulares y simples o pluricelulares y bastante complejos, generalmente tienen un pie y una cabeza secretora uni o pluricelular. El producto secretado frecuentemente se aloja entre la pared externa de la célula y la cutícula, que se levanta y al final se rompe. Esa cutícula en algunos casos puede regenerarse y repetirse la acumulación de líquido, o el tricoma puede degenerar después de una sola excreción.[29]
Existen tricomas especializados en la defensa de la planta contra los animales herbívoros. Un ejemplo es el de los tricomas de la epidermis de las hojas y de las estípulas de la alfalfa (Medicago scutellata), que presenta pelos secretores capitados, con un largo estípite, que defienden a la planta contra las larvas del gorgojo de la alfalfa, Hypera postica. La secreción de estos pelos se acumula debajo de la cutícula y en contacto con las larvas del insecto se exuda un fluido pegajoso, el cual las inmoviliza. En las especies silvestres de papa hay tricomas defensivos que atrapan áfidos. La resistencia de Passiflora adenopoda a las larvas de las mariposas del género Heliconius se debe a sus tricomas ya que las larvas al desplazarse sobre la hoja quedan empaladas en ellos.[30][31]


Otros tricomas glandulares defensivos son urticantes. Este tipo de tricoma es característico de cuatro familias de dicotiledóneas: urticáceas, euforbiáceas, loasáceas e hidrofiláceas. Sirven como mecanismos de defensa de la planta debido a que almacenan toxinas en la vacuola central de sus células, las que pueden ser expulsadas cuando un animal toca la planta. Son células epidérmicas o subepidérmicas muy largas, de cuello largo y afilado y con la base inserta en una emergencia en forma de copa compuesta por varias células. Cuando se toca la cabeza de este tipo de tricoma, se rompe el cuello siguiendo una línea oblicua de modo que el cuerpo del pelo queda convertido en una aguja hipodérmica que inyecta el líquido irritante contenido en su interior. Ese líquido está commpuesto por ácido fórmico además de acetilcolina e histamina entre otros compuestos.[30]
En las plantas insectívoras la epidermis foliar esta cubierta por glándulas digestivas. En Drosera, por ejemplo, la cabeza de estas glándulas complejas está recubierta por una cutícula provista de numerosos poros; las capas más externas son células secretoras con paredes laberínticas; la capa subyacente está formada por células de tipo endodérmico, con engrosamientos radiales impermeables. El centro está ocupado por traqueidas rodeadas por una capa de células de transferencia. Las células secretoras son muy complejas en su funcionamiento porque además de secretar enzimas digestivas absorben los nutrientes digeridos.[32] Utricularia, un género de plantas palustres o acuáticas sumergidas perteneciente a las lentibulariáceas, posee órganos vegetativos especializados para la captura y la digestión de pequeños organismos, los que se denominan «trampas» o utrículos. Tales trampas tienen forma globosa y presentan una boca rodeada de apéndices filiformes, con un zaguán cerrado por una membrana que constituye la «puerta». La membrana presenta apéndices sensitivos que, al ser tocados por una presa, determinan el brusco desplazamiento de la membrana hacia dentro de la trampa, provocando una súbita corriente de agua hacia el interior que arrastra también a la presa. Cuando la trampa se llena de agua la membrana cae, cerrando el utrículo. Internamente, los utrículos presentan pelos glandulares especializados en la secreción de enzimas que digieren a las presas.[33] Pinguicula es otra planta carnívora perteneciente también a las lentibulariáceas, sus hojas capturan pequeños insectos, que luego digiere. Vive en ambientes donde el nitrógeno es escaso, y suple su falta con el nitrógeno de las proteínas de los insectos. Como todos los miembros de su género, estas láminas foliares están densamente cubiertas por glándulas pediceladas mucilaginosas y con glándulas digestivas sésiles y planas. Las glándulas pediceladas están constituidas por un grupo de células secretoras en la parte superior de un pie unicelular y producen una secreción mucilaginosa con la cual forman gotas visibles en la superficie de la hoja. Esta apariencia húmeda probablemente ayude a atraer a sus presas en busca de agua, un fenómeno similar se observa también en las droseras. Estas gotas secretadas tienen como función principal atrapar a los insectos, que al hacer contacto, inducen la liberación de una cantidad adicional de mucílago. El insecto pelea, tocando y activando más glándulas y atrapándose aún más a sí mismo en el mucílago. Por otra parte, Pinguicula puede doblar el borde de sus hojas ligeramente gracias a su tigmotropismo, haciendo que una mayor cantidad de glándulas entre en contacto con el insecto atrapado. Luego, mediante la liberación del contenido de sus glándulas sésiles, las cuales se encuentran sobre la superficie de la hoja, digiere al insecto por acción de diversas enzimas. Estas enzimas, entre las cuales se incluyen la amilasa, la esterasa, la fosfatasa, la peptidasa y la ribonucleasa digieren la mayor parte de los componentes del cuerpo del insecto. Estos fluidos posteriormente son absorbidos a través de los poros en la cutícula de sus hojas, dejando solo en la superficie el exoesqueleto de quitina de los insectos más grandes.[34]
Hidatodos

Las hojas de 350 géneros pertenecientes a 115 familias eliminan agua líquida a través de hidatodos por el proceso de gutación. En algunas plantas tropicales, como Colocasia (Araceae), la cantidad eliminada por noche supera los 300 ml.[35]
Los hidatodos pueden ser de tipo activo o de tipo pasivo. Estos últimos, frecuentes en las gramíneas, eliminan agua por ósmosis cuando aumenta la presión radical. Los hidatodos activos, en cambio, eliminan agua con independencia de la presión de las raíces. Los hidatodos epidérmicos o tricómicos secretan una solución acuosa con algunos compuestos orgánicos o inorgánicos (secreción ecrina, por bombeo de iones metálicos a través de la membrana plasmática). En las hojas jóvenes de Cicer arietinum los hidatodos epidérmicos consisten en un pedúnculo uniseriado y una cabeza oval pluricelular. La solución acuosa se acumula bajo la cutícula y cuando alcanza un cierto volumen se abren poros en la cutícula y aparecen gotitas en la superficie. Los hidatodos epitémicos presentan un tejido parenquimático especializado, el epitema, situado al final de una o varias vénulas de las hojas. Rodeando el epitema hay una vaina de células dispuestas apretadamente, a menudo con las paredes adyacentes cutinizadas, formando una capa de tipo endodérmico, que impide el reflujo hacia el apoplasto. Muchas de estas células están diferenciadas como células de transferencia. Hacia el exterior, el epitema está limitado por la epidermis, y la salida del agua se produce a través de uno o varios estomas modificados, que se diferencian porque las células oclusivas usualmente no pueden cerrar el ostíolo.[35]
Mesófilo
El interior de la hoja, entre la epidermis adaxial y abaxial, se denomina mesófilo. El mesófilo está constituido por tejidos vasculares, que forman las venas de las hojas, y un tejido parenquimático especializado para realizar la fotosíntesis denominado clorénquima.
Clorénquima


Generalmente las células del parénquima clorofiliano tienen paredes delgadas. Dejan abundantes espacios intercelulares que constituyen un sistema de aireación bien desarrollado para facilitar el intercambio de gases necesario para permitir la asimilación del dióxido de carbono. Sus células tienen un número variable de cloroplastos, que durante ciertos momentos del día pueden contener almidón de asimilación. Presentan numerosas vacuolas o una sola. El clorénquima puede ser de dos tipos. El parénquima clorofiliano en empalizada está formado por células cilíndricas, alargadas, que logran así más superficie y menor volumen, muy ricas en cloroplastos y con espacios intercelulares pequeños. El segundo tipo es el parénquima clorofiliano lagunar o esponjoso que está formado de células cortas, redondeadas o variadamente lobuladas, menos ricas en cloroplastos y que dejan espacios intercelulares grandes, llamados lagunas, por donde circula el aire necesario para el intercambio gaseoso y la respiración.[36]
Estructura del mesófilo
El mesófilo puede ser homogéneo, cuando está formado un solo tipo de células de clorénquima. En este caso, las células pueden ser más o menos redondeadas, como sucede en plantas herbáceas como el lino (Linum usitatissimum) o la lechuga (Lactuca sativa), o estar constituido por células alargadas como sucede en la remolacha (Beta vulgaris). Las especies de Eucalyptus con hojas péndulas también presentan este tipo de mesófilo.[36]

Cuando el clorénquima de la hoja se diferencia en parénquima en empalizada y parénquima lagunoso, el mesófilo se dice heterogéneo. Según la ubicación de los dos tipos de clorénquima se distinguen tres tipos de estructura del mesófilo. El mesófilo dorsiventral es el tipo de estructura en el que el clorénquima en empalizada está hacia la cara adaxial de la hoja y el clorénquima lagunoso hacia la cara abaxial, como es el caso de la vid (Vitis) y el ligustro (Ligustrum). Esta organización se encuentra usualmente en las hojas que se orientan horizontalmente. El número de capas de clorénquima en empalizada es variable: hay una sola en el caso del tabaco (Nicotiana tabacum) y de la batata (Ipomoea batatas), dos en el laurel rosa (Nerium oleander) y en la alfalfa (Medicago sativa) y tres en el caso del peral (Pyrus). El número de capas del clorénquima en empalizada puede incluso variar de una hoja a otra del mismo individuo. Así, en el caso del arce (Acer platanoides), un árbol de copa amplia, las hojas periféricas tienen más capas de clorénquima que las que se encuentran en interior de la copa. Asimismo, las hojas que están más expuestas al sol son más gruesas que las que están en la sombra, presentan un mayor desarrollo del clorénquima en empalizada y un mayor número de cloroplastos. El mesófilo isobilateral es aquella estructura en la que el clorénquima en empalizada se encuentra sobre las dos caras del limbo foliar y el parénquima lagunoso queda entre medio de ellas. Esta organización es característica de las plantas con hojas erguidas o péndulas, tales como el clavel (Dianthus). El mesófilo céntrico es un tipo de estructura que se encuentra en especies con hojas muy angostas, casi cilíndricas. En este tipo de organización del mesófilo el parénquima en empalizada adaxial forma una capa casi continua con el abaxial, como por ejemplo en Salsola. Esta estructura es frecuente en plantas xerofíticas y plantas suculentas, cuyo mesófilo contiene células pequeñas, el clorénquima en empalizada está más desarrollado que el esponjoso y frecuentemente está reforzado por esclerénquima.[37]
Funciones
Transpiración

La transpiración es un proceso similar al de evaporación. Es una parte del ciclo del agua y consiste en la pérdida de vapor de agua a partir de partes de la planta, especialmente de las hojas, aunque también puede producirse a partir de los tallos. La mayor parte de la transpiración, no obstante, se produce a través de los estomas. La apertura y cierre de estas estructuras tiene un costo energético asociado, pero permite la difusión del dióxido de carbono necesario para la fotosíntesis desde el aire hacia el interior de la hoja, la salida de oxígeno desde dentro de la hoja hacia el exterior y la pérdida de vapor de agua. La transpiración también refresca a las plantas y permite el flujo de masa de los nutrientes minerales y del agua desde las raíces hacia las hojas. Este flujo de masa de agua desde las raíces hasta las hojas está causado por la disminución de la presión hidrostática en las partes superiores de las plantas debido a la difusión de agua desde los estomas a la ambiente. El agua es absorbida desde el suelo hacia las raíces por el proceso de ósmosis, y cualquier mineral disuelto en el agua será acarreado hacia las hojas a través del xilema.[38]
La tasa de transpiración está directamente relacionada con la evaporación de las moléculas de agua de la superficie de la planta. La transpiración estomática representa la mayor parte de la pérdida de agua por una planta, si bien algo de evaporación directa también se lleva a cabo a través de la cutícula de las hojas y de los tallos jóvenes. La cantidad de agua que se desprende depende en cierto modo de la cantidad de agua que las raíces de la planta han absorbido. También depende de las condiciones ambientales tales como la intensidad de luz, la humedad, la velocidad del viento y la temperatura. La transpiración se produce cuando el sol calienta el agua dentro de la hoja. Este calentamiento cambia gran parte del agua en vapor de agua y este gas puede escapar al medio a través de los poros estomáticos. La transpiración ayuda a enfriar el interior de la hoja debido a que el vapor que escapa hacia la atmósfera ha absorbido el calor. La cantidad de agua que una planta pierde por la transpiración estomática depende de su tamaño, de la intensidad lumínica, del grado de apertura de los estomas y la demanda evaporativa de la atmósfera que rodea a la hoja.[39] Un árbol completamente crecido puede perder varios cientos de litros de agua a través de sus hojas en un día caluroso y seco. Alrededor del 90% del agua que entra en las raíces de la planta se usa para este proceso. En tal sentido, se denomina eficiencia del uso del agua a la relación entre la masa de agua transpirada con respecto a la masa de materia seca producida por una planta. Esta relación se halla entre 200 y 1000 para la mayoría de los cultivos, es decir, las plantas de los cultivos transpiran entre 200 a 1000 kg de agua por cada kg de materia seca producida.[40]
Fotosíntesis
La fotosíntesis es el proceso por el cual la materia inorgánica se convierte en compuestos orgánicos gracias a la energía aportada por la luz. En este proceso la energía lumínica se transforma en energía química estable, siendo el adenosín trifosfato (ATP) la primera molécula en la que queda almacenada esa energía química. Con posterioridad, el ATP es utilizado en interior de la célula para sintetizar moléculas orgánicas de mayor estabilidad, tales como carbohidratos. Cada año los organismos fotosintetizadores fijan en forma de materia orgánica en torno a 100.000 millones de toneladas de carbono (3000 toneladas de carbono por segundo aproximadamente).[41][42]
Los orgánulos citoplasmáticos encargados de la realización de la fotosíntesis son los cloroplastos, unas estructuras polimorfas y de color verde (esta coloración es debida a la presencia del pigmento clorofila) propias de las células vegetales. En el interior de estos orgánulos se halla una cámara que contiene un medio interno llamado estroma, que alberga diversos componentes, entre los que cabe destacar enzimas encargadas de la transformación del dióxido de carbono en materia orgánica y unos sáculos aplastados denominados tilacoides o lamelas, cuya membrana contiene pigmentos fotosintéticos. En términos medios, una célula foliar tiene entre cincuenta y sesenta cloroplastos en su interior.[43][41] Los organismos que tienen la capacidad de llevar a cabo la fotosíntesis son llamados fotoautótrofos (otra nomenclatura posible es la de autótrofos, pero se debe tener en cuenta que bajo esta denominación también se engloban aquellas bacterias que realizan la quimiosíntesis) y fijan el CO2 atmosférico. En la actualidad se diferencian dos tipos de procesos fotosintéticos, que son la fotosíntesis oxigénica y la fotosíntesis anoxigénica. La primera de las modalidades es la propia de las plantas superiores, las algas y las cianobacterias, donde el dador de electrones es el agua y, como consecuencia, se desprende oxígeno. Mientras que la segunda, también conocida con el nombre de fotosíntesis bacteriana, la realizan las bacterias purpúreas y verdes del azufre, en las que en dador de electrones es el sulfuro de hidrógeno, y consecuentemente, el elemento químico liberado no será oxígeno sino azufre, que puede ser acumulado en el interior de la bacteria, o en su defecto, expulsado al agua.[43]
La ecuación química general para la fotosíntesis es:
Dióxido de carbono + donante de electrones + energía lumínica → carbohidrato + donante de electrones oxidado En la fotosíntesis oxigénica el agua es el donante de electrones y, debido a que su hidólisis libera oxígeno, la ecuación para este proceso es:
- 2n CO2 + 4n H2O + fotones → 2(CH2O)n + 2n O2 + 2n H2O
- Dióxido de carbono + agua + energía lumínica → carbohidrato + oxígeno + agua
Frecuentemente 2n moléculas de agua se cancelan en ambos miembros de la ecuación, lo que produce:
Productividad
- Producción primaria neta de los principales ecosistemas terrestres.[44]
| Ecosistema | Producción primaria neta (peso seco en g·m-2·año-1) |
|---|---|
| Tundras árticas | 100 |
| Tundras de arbustos pequeños | 250 |
| Taiga del norte | 450 |
| Taiga del sur | 850 |
| Bosque de hayas | 1300 |
| Bosque de robles | 900 |
| Estepas templada | 120 |
| Estepa seca | 420 |
| Desierto de semi arbustos pequeños | 122 |
| Desiertos subtropicales | 50 |
| Bosques subtropicales | 2450 |
| Bosque tropical lluvioso | 3250 |
| Pantanos de Sphagnum con bosque | 340 |
| Manglares | 930 |
| Océano abierto | 125 |
| Tierra agrícola | 650 |
Adaptaciones especiales de las hojas
Algunas plantas, como los cactus, han transformado sus hojas en espinas; son los troncos, carnosos y aplanados, los que ejercen la función fotosintética. Las hojas de los troncos subterráneos, como en la cebolla, pueden transformarse en órganos de reserva de nutrientes. El caso más extremo parece ser el de las plantas carnívoras, en que la hoja se transforma en una trampa, como si de un predador se tratara.
- Catáfilos: son hojas sencillas, escamiformes, papiráceas y sin clorofila. Cuando son un intermedio entre cotiledones y hojas verdaderas se denominan protofilos. También son catáfilos las escamas de las yemas invernantes así como también las que recubren los bulbos, así como cualquier hoja que queda por debajo de los nomofilos y por encima de los cotiledones.

- Zarcillos: son órganos de los que se sirven ciertas plantas para trepar o sujetarse a otras, enroscándose. Existe una gran variedad de zarcillos, siendo los más importantes de tipo caulinar —derivados de tallos— o bien, de tipo foliar —derivados de hojas— que son los que interesan aquí. Son hojas modificadas que crecen helicoidalmente, de manera que si el cuerpo al que están unidos, crece o se aleja, el zarcillo puede estirarse sin romperse ni hacer perder el soporte al vegetal. Se observan zarcillos, por ejemplo, en la vid (Vitis vinifera) o en el tumbo (Passiflora mollissima).

- Espinas: son hojas aciculares cortas y lignificadas, no clorofilianas, que cumplen función de protección. Igual que en el caso de los zarcillos, no todas las espinas son hojas modificadas; algunas son apéndices epidérmicos —los aguijones de las rosas—, otras son tallos modificados y, por último, muchas son hojas modificadas. Este último es el caso en las crasuláceas y las cactáceas, que son las más populares de las plantas espinosas. Estas plantas almacenan agua en sus hojas y tallos suculentos ya que su hábitat se halla típicamente en zonas secas y calurosas, donde el agua es escasa.
Heterofilia foliar

Es el fenómeno consistente en que en una misma planta aparezcan hojas asimiladoras (nomofilos) diferentes. En muchas plantas existen diferencias entre, por ejemplo, las hojas de las ramas más altas y las más bajas, o entre los ejemplares arbustivos y los arbóreos. Hay heterofilia propiamente dicha cuando hay dos o más clases de hojas con diferencias cualitativas de morfología o tamaño.
Ejemplos de heterofilia los encontramos en ciertos helechos tropicales, en Eichornia azurea que tiene hojas ovales y lineares, o en ranúnculos acuáticos, cuyas hojas superficiales son muy diferentes de las sumergidas. En el género Juniperus hay especies con hojas en aguja y otras, como Juniperus thurifera, con hojas en escama, pero en estas últimas las hojas de las ramas juveniles son aciculares.
También el guapuruvú (Schizolobium parahyba) presenta hojas similares a las del helecho arborescente al principio y hojas grandes y distintas al desarrollarse la planta, y cuando aparecen estas hojas es cuando se desarrolla la floración. Además, en esta planta no existe diferenciación entre las hojas, tronco y ramas, ya que en todo el árbol se produce la fotosíntesis.
Falsas hojas
- Filodios y filocladios: Son órganos laminares semejantes a hojas, pero que por su origen evolutivo, su ontogénesis (su proceso de desarrollo) y su anatomía interna, resultan no serlo. En general se trata de peciolos y tallos dilatados que cumplen en algunas plantas la función asimiladora que normalmente corresponde a las hojas, a las que funcionalmente sustituyen. Uno de los ejemplos más conocidos de filodios lo ofrecen especies de los géneros Acacia y Ruscus; los ejemplares nuevos y las ramas del año llevan hojas verdaderas compuestas, mientras la mayor parte del "follaje" está constituida por filodios o filocladios que son más duros y alargados.
- Filoides. Es el nombre que se da a órganos semejantes en su morfología y función a las hojas, pero sin su anatomía interna, que se encuentran en plantas terrestres (musgos y hepáticas) que divergieron evolutivamente de las plantas vasculares, portadoras de verdaderas hojas, antes de la evolución de la anatomía foliar.
Senescencia foliar
La senescencia de las hojas es una serie de eventos ordenados y cuidadosamente controlados que permiten a las plantas conservar sus recursos, prepararse para un período de reposo y descartar tejidos ineficiente.[6] En las plantas anuales algunas de las hojas mueren precozmente si bien la mayor parte de las hojas mueren al mismo tiempo que el resto de los órganos. En los árboles, arbustos y plantas herbáceas perennes, en cambio, la defoliación es un fenómeno periódico, muy complejo, que se produce, en la mayoría de las especies por un mecanismo de abscisión.[7] Siguiendo el ritmo y el modo en el que ocurre la defoliación, se pueden distinguir distintos tipos de plantas. Así, hay plantas con defoliación anual (Caducifolias), en las que las hojas viven solo algunos meses, generalmente desde la primavera hasta el otoño. Dentro de este grupo se reconocen aquellas —como los encinos— en las que las hojas amarillean y mueren sin separarse del tallo, de las plantas de hoja caduca en las que la defoliación es un fenómeno activo que ocurre a través de tejidos especializados que forman la denominada zona de abscisión. En las primeras, las hojas que quedan sujetas a los tallos por mucho tiempo ya que su caída es pasiva y generalmente determinada por el viento. En las segundas, la caída de las hojas es precedida por una migración de sustancias desde las hojas hacia el tallo. Las hojas amarillean y, a veces, enrojecen por la acumulación de antocianinas y se separan del tallo estando todavía vivas, muriendo después de caer.[7] Por otro lado, hay plantas con follaje persistente (Perennifolias) —como los pinos— cuyas hojas viven de dos a cinco años. El conjunto de las hojas, el follaje, se renueva parcialmente cada año. La caída de las hojas en este caso es también un fenómeno activo ya que está determinado por la formación de una zona de abscisión.[7]
El color de las hojas en otoño

_crop.jpg.webp)
Las hojas de muchas especies muestran un color rojo bien diferenciado durante el otoño. En algunos árboles, como el arce rojo Americano (Acer rubrum) o el roble escarlata (Quercus coccinea), los flavonoles —un tipo de flavonoide— incoloros se convierten en antocianinas rojas cuando la clorofila de sus hojas se degrada.[8] En otoño, cuando la clorofila se descompone, los flavonoides incoloros se ven privados del átomo de oxígeno unido a su anillo central, lo que los convierte en antocianinas, dando colores brillantes. Esta transformación química que consiste solo en la pérdida de un átomo de oxígeno es la responsable de nuestra percepción de los colores del otoño. Estos pigmentos probablemente son las que protegen a las hojas del efecto de los rayos UV del Sol. Se especula que esta protección de las hojas aumenta su efectividad para transportar nutrientes durante su senescencia.[45][46]
Mecanismo anatómico y regulación hormonal de la abscisión
Las heladas y bajas temperaturas del otoño dañan de forma rápida los tejidos vivos de las hojas y de los pecíolos de muchas especies. Una vez muertas, los recursos que las células de las hojas poseen no se hallarán disponibles para la planta. Por esta razón las especies caducifolias utilizan una secuencia de procesos coordinados para extraer sistemáticamente los valiosos recursos de las hojas antes que estas mueran. Una vez que tales recursos son capturados por las plantas, los tejidos muertos y moribundos de las hojas se descartan. Hacia el final de este proceso de senescencia se desarrollan líneas de fractura en la base de las hojas. Estas líneas de fractura impiden que la planta se exponga a daños subsecuentes por la caída de las hojas gracias al sellado de los tejidos que pronto serán descartados.[6] La caída de las hojas antes de su muerte se debe a la intervención de tejidos especializados que constituyen una región definida en la base de la hoja, a menudo visible exteriormente como una constricción en forma de anillo, que se denomina zona de abscisión. Esta zona aparece en forma de un disco de células pequeñas con paredes delgadas, alineadas regularmente y sin ningún tejido de sostén. La zona de abscisión se diferencia después en dos capas superpuestas: una capa de abscisión o separación hacia la hoja y una capa protectora suberosa hacia el tallo. En la capa de abscisión o separación las laminillas medias y a menudo las paredes primarias de las células se gelifican, de manera que las células ya no están soldadas entre sí y se separan. La hoja queda sostenida solamente por los hacecillos vasculares, donde los vasos son obstruidos por tílides, y entonces cae, ya sea por su propio peso o por la acción del viento. En la capa protectora suberosa las paredes celulares se impregnan de suberina y forman un súber cicatricial que protege la cicatriz dejada sobre el tallo por la caída de las hojas. El súber cicatricial puede formarse antes de la caída de las hojas —como en el caso de la magnolia— o inmediatamente después —como en la higuera—. En las especies leñosas la capa protectora tarde o temprano es reemplazada por la peridermis que se desarrolla debajo de la capa protectora en continuidad con la peridermis de otras partes del tallo. Las auxinas son agentes inhibidores de la abscisión foliar, mientras el etileno es un agente inductor, que se utiliza como defoliante en la cosecha mecánica de algunos cultivos.[7] La regulación de la abscisión es una herramienta corriente en agricultuta en conexión con la creciente mecanización de muchas prácticas, tales como la defoliación controlada, el raleo de los frutos y el ajuste del momento de la cosecha de los mismos.
Evolución de las hojas
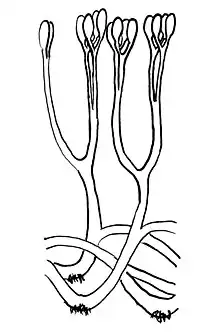
Durante la evolución de las plantas las hojas se han originado al menos en dos ocasiones independientes. El legado de estos eventos evolutivos históricos está representado en las floras actuales por los microfilos de las licofitas (licopodios, Selaginella e Isoetes) y los megafilos de las eufilofitas (helechos, gimnospermas y angiospermas). Se considera que los microfilos, con su vasculatura característica y su ausencia de ramificación, han evolucionado a partir de excrecencias espinosas y son anteriores a los megafilos en el registro fósil de las plantas terrestres. De mayor importancia, sin embargo, fue el origen de los megafilos en las plantas vasculares a través de la modificación del desarrollo de las ramas laterales debido a que la gran mayoría de las aproximadamente 250.000 especies de plantas con flores existentes, así como la mayoría de las gimnospermas, utilizan un megafilo de lámina plana con una red de venas para la captación de la energía solar y la asimilación de carbono fotosintético. Los megafilos alteraron, de hecho, la trayectoria evolutiva de las plantas y de los animales terrestres, los ciclos biogeoquímicos de los nutrientes, del agua y del dióxido de carbono y el intercambio de energía entre la superficie terrestre y la atmósfera.[48] Una medida de su éxito en ambientes terrestres es la capacidad de las hojas para soportar condiciones climáticas extremas entre los trópicos y la tundra, mientras que al mismo tiempo facilitan la fijación neta de aproximadamente 207 000 millones de toneladas de CO2 (56,4 × 1015 g C) año-1 a escala global. Esta producción primaria proporciona energía para casi todas las formas de vida terrestre en la Tierra, especialmente a los tetrápodos y a los insectos, y enlazan muchos ecosistemas y procesos biogeoquímicos.[49]

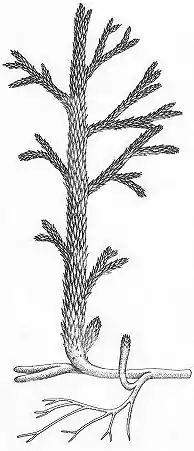
Evidentemente, las hojas son un éxito evolutivo de las plantas terrestres. Sin embargo, la llegada de los grandes megafilos tuvo lugar alrededor de 40-50 millones de años después del origen de las plantas vasculares, lo que sugiere que estaban lejos de ser una consecuencia inevitable de la evolución. Las primeras plantas vasculares ancestrales, que datan de finales del Silúrico, 410 millones de años atrás, estaban compuestos por un tallo simple o ramificado con esporangios, pero sin hojas.[51] Sorprendentemente, las plantas continuaron careciendo de hojas durante los siguientes 40-50 millones de años, cuando los megafilos finalmente se generalizaron hacia finales del período Devónico, 360 millones de años atrás.[52][53][54][55] Esta aparición tardía de las hojas, una modificación estructural aparentemente simple, es sorpresiva al menos por tres razones. En primer lugar, la evidencia paleontológica muestra que el marco estructural necesario para el montaje evolutivo de una hoja simple (meristemas, vascularización, cutícula y la epidermis) se hallaban establecidos entre las plantas mucho antes de la llegada de los grandes megafilos. En segundo lugar, durante el mismo intervalo ocurrió una explosión sin precedentes de la innovación evolutiva de la historia de vida de las plantas, la cual fue testigo del ascenso de los árboles a partir de ancestros herbáceos, así como la evolución de los ciclos de vida complejos, incluyendo la invención de la semilla. En tercer lugar, el pequeño megafilo de la planta Eophyllophyton bellum de principios del Devónico, muestra que las plantas tenían la capacidad para producir un megafilo simple mucho tiempo antes de que estas estructuras se generalizaran.[56][57]
En casi todas las plantas actuales las hojas constituyen una adaptación para incrementar la cantidad de luz solar que puede ser capturada para realizar la fotosíntesis. Se acepta que las hojas han evolucionado más de una vez en el transcurso de la evolución de las plantas y que, probablemente, se originaron como estructuras espinosas que actuaban como estructuras de defensa contra los herbívoros.
Las riniópsidas (Rhyniopsida) del yacimiento paleontológico de Rhynie Chert no estaban compuestas más que por ejes delgados sin ningún tipo de ramificación. Las trimerópsidas (Trimerophytopsida) del Devónico temprano son la primera evidencia que se dispone de algo que puede ser considerado como una hoja. Este grupo de plantas vasculares son reconocibles por sus masas de esporangios terminales, que adornan los extremos de los ejes que se pueden bifurcar o trifurcar.[58] Algunos organismos, tales como Psilophyton presentaban excrecencias sobre los tallos, pequeñas y espinosas, que carecían de su propio aporte vascular.[59][60] Casi al mismo tiempo, las Zosterophyllopsida fueron adquiriendo importancia. Este grupo es reconocible por sus esporangios con forma de riñón que crecían sobre cortas ramas laterales cercanas a los ejes principales.[58] La mayoría de los miembros de este grupo tenía espinas pronunciado sobre sus ejes. Sin embargo, tales espinas carecían de traza foliar, y la primera evidencia de enaciones vascularizados ocurrió en el género Asteroxylon . Las espinas de Asteroxylon presentaban una primitiva fuente vascular, ya que cada «hoja» estaba inervada por una traza foliar que surgía de la protoestela. El género Baragwanathiaaparece en el registro fósil un poco antes, a finales del Silúrico.[61] En este organismo, tales trazas foliares se continúan dentro de la «hojas» para formar la nervadura central.[62]
Asteroxylon[63] y Baragwanathia se consideran como licopodios primitivos.[58] Los licopodios existen todavía en la actualidad, siendo Isoetes un ejemplo bastante conocido. Estos licopodios actuales también presentan microfilos con una sola traza foliar y de pequeño tamaño. La mayor diferencia con los Lepidodendrales extintos, es que estos exhibían microfilos de hasta un metro de longitud, pero en casi todos los casos, solo existía una única traza foliar.[58]
Las hojas más familiares, los megafilos, han evolucionado en cuatro oportunidades diferentes y de modo independiente: en los helechos, las colas de caballo, las progimnospermas y en las plantas con semilla.[64] Parece que se han originado a partir de la dicotomización de ramas, que por primera vez se superpusieron entre sí, y, finalmente se desarrolló una "palma" que gradualmente evolucionó en estructuras similares a hojas. Entonces, según esta teoría del teleoma, los megafilos se componen de un grupo de ramas palmeadas[62] En cada uno de los cuatro grupos que habrían de desarrollar megafilos, sus hojas evolucionaron por primera vez durante el Devónico tardío y el Carbonífero temprano, diversificando rápidamente hasta que los diseños estructurales básicos quedaron establecidos en el Carbonífero medio.[64] La falta de diversificación posterior puede ser atribuida a limitantes de desarrollo.[64] No obstante, persiste también la duda acerca de las razones por las cuales las hojas tardaron tanto tiempo en aparecer sobre el planeta. De hecho, las plantas han estado sobre la superficie por lo menos por 50 millones de años antes de que los megafilos se hicieran significativos. Sin embargo, los mesófilos se conocen desde el Devónico temprano (el género Eophyllophyton) por lo que el desarrollo no pudo ser una barrera o una limitante para la aparición de las hojas ya que uno de sus constituyentes principales ya existía.[65] la mejor explicación indica que el CO₂ atmosférico estaba declinando rápidamente durante este tiempo, cayendo cerca del 90% durante el devónico.[66] Esto se correspondió con un incremento de 100 veces en la densidad de los estomas. Parece ser que la baja densidad estomática en el Devónico temprano significaba que la evaporación era limitada y que las hojas se sobre-calentarían si alcanzaban un mayor tamaño. La densidad estomática no podría incrementar, ya que las primitivas estelas y los sistemas radicales limitados no serían capaces de suministrar suficiente agua para suplir la tasa de transpiración.[67]
Usos de las hojas




Los seres humanos utilizamos una gran cantidad de plantas por sus hojas. En algunos casos, las hojas son comestibles (lechuga, acelga, escarola) y son una fuente de fibras y minerales para la dieta. En el caso de muchas plantas condimenticias, las hojas frescas o secas se utilizan para sazonar carnes, sopas, estofados y otros alimentos (romero, salvia, menta). También las hojas pueden llevar sustancias químicas que las convierten en una excelente fuente de esencias perfumíferas, como es el caso del lemongrass (Cymbopogon citratus). Las hojas secas del tabaco se utilizan para fumar en todo el mundo.
Las hojas adultas se usan para techar casas rurales (Sabal mexicana entre otras palmeras) Las hojas de huano que se usan para techar, si están bien trabajadas, duran de 15 a 20 años. Las hojas preferidas para el techado son las de los individuos jóvenes ya que estos aún no alcanzan grandes alturas y se obtienen más fácilmente y además son de mayor tamaño. [68][69]
La práctica de mascar las hojas de ciertas plantas es usual en muchas culturas. Así, en los Andes peruanos los pueblos Quechua, Aymará entre otros, aprecian las hojas de Erythroxylum coca por las virtudes mágicas que se cree que éstas poseen. De hecho, juegan un papel importante en la medicina popular ya que es prescripta para quebraduras de huesos, dolores, infecciones y disfunciones respiratorias. Esta costumbre es más que accidental ya que las hojas contienen cocaína, el cual es un potente anestésico. El uso de las hojas de coca y la mitología asociada han existido por más de 4000 años.[70][71] Las artesanías son actividades, destrezas o técnicas empíricas, practicadas tradicionalmente por el pueblo, mediante las cuales con intención o elementos artísticos, se crean o producen objetos destinados a cumplir una función utilitaria cualquiera, o bien se las adorna o decora con el mismo o distinto material realizando una labor manual individualmente o en grupos reducidos, por lo común familiares e infundiendo en los caracteres o estilos típicos, generalmente concordes con los predominantes en la cultura tradicional de la comunidad".
Existen diferentes tipos de artesanías que utilizan hojas en su confección y que se realizan por diferentes etnias a lo largo de todo el mundo. Entre ellas, la cestería o tejidos duros, los tejidos blandos, los adornos corporales, los instrumentos de caza y pesca, los instrumentos musicales, etc. Para su confección se utilizan hojas de diferentes tipos de palmas.[72]
| Nombre científico | Nombre común | Familia |
|---|---|---|
| Lactuca sativa | Lechuga | Asteraceae (Compositae) |
| Spinacia oleracea | Espinaca | Chenopodiaceae |
| Brassica sp. | coles y repollos | Brassicaceae |
| Allium cepa y Allium sativum | cebolla y ajo | Alliaceae |
| Allium fistulosum y Allium porrum | cebolla de verdeo y puerro | Alliaceae |
| Beta vulgaris | acelga | Chenopodiaceae |
| Petroselinum crispum | perejil | Apiaceae |
| Nasturtium officinale | berro | Brassicaceae |
| Salvia rosmarinus | romero | Lamiaceae |
| Origanum vulgare | orégano | Lamiaceae |
| Cichorium intybus | endivia | Asteraceae |
| Borago officinalis | borraja | Boraginaceae |
| Camelia sinensis | té | Theaceae |
| Ilex paraguariensis | yerba mate | Aquifoliaceae |
| Stevia rebaudiana | azúcar verde | Asteraceae |
| Mentha aquatica | menta | Lamiaceae |
| Taraxacum officinale | diente de léon, taraxacón | Asteraceae |
| Foeniculum vulgare | hinojo | Apiaceae |
Notas
- Una base de hoja decurrente no está en realidad causada por la unión, en sí, de la hoja al tallo, sino por el crecimiento/multiplicación, al nivel del punto de unión, de las células del primordio foliar en sendas decurrencias laterales y/o, eventualmente, en alas caulinares más o menos largas mientras el tallo crece y desarrollándose, también, dicho primordio hacia arriba en hoja propiamente dicha; y así sucesivamente con cada uno de los primordios foliares que aparezcan a lo largo del tallo.
Referencias
- Jürgen R. Hoppe. «5 Morphologie des Blattes». Allgemeine Botanik (en alemán). Alemania: Institut für Systematische Botanik und Ökologie, Universität Ulm. p. 70. Consultado el 10 de septiembre de 2011.
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema 2: Hoja, 2.7. Hojas de Pteridofitas». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 31 de octubre de 2009. Consultado el 10 de septiembre de 2010.
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema 2: Hoja, Hojas de Gimnospermas». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 31 de octubre de 2009. Consultado el 10 de septiembre de 2010.
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema 2: Hoja. 2.5. Hoja de Monocotiledóneas». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 22 de enero de 2009. Consultado el 10 de septiembre de 2010.
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema 2: Hoja. 2.2. Hoja de Dicotiledóneas». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 27 de septiembre de 2010. Consultado el 10 de septiembre de 2010.
- Coder, K.D. 1999. Falling Tree Leaves: Leaf Abscission University of Georgia Daniel B.Warnell School of Forest Resources Extension publication.
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema 21: Anatomía foliar. 21.10. Abscisión». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 10 de abril de 2009. Consultado el 10 de junio de 2011.
- Clifford M. N.(2000): Anthocyanins – nature, occurrence and dietary burden. In: Journal of the Science of Food and Agriculture. Bd. 80, Nr. 7, S. 1063–1072. doi <1063::AID-JSFA605>3.0.CO;2-Q 10.1002/(SICI)1097-0010(20000515)80:7<1063::AID-JSFA605>3.0.CO;2-Q
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema 2: Hoja, 2.1. Definición, origen, función y sucesión foliar». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 23 de marzo de 2009. Consultado el 10 de septiembre de 2010.
- Strassburger, E. 1994. Tratado de Botánica. 8.ª edición. Omega, Barcelona, 1088 p. ISBN 84-7102-990-1
- Gola, G., Negri, G. y Cappeletti, C. 1965. Tratado de Botánica. 2.ª edición. Editorial Labor S.A., Barcelona, 1110 p.
- Charles R. Tischler, Justin D. Derner, H. Wayne Polley, and Hyrum B. Johnson, Response of Seedlings of Two Hypogeal Brush Species to CO2 Enrichment, Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, pp. 104-106, archivado desde el original el 28 de septiembre de 2011, consultado el 18 de junio de 2011.
- Baraloto, C.; Forget, P.-M. (2007), «Seed size, seedling morphology, and response to deep shade and damage in neotropical rain forest trees», American Journal of Botany 94: 901, doi:10.3732/ajb.94.6.901.
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema 2: Hoja, 2.4. Pecíolo y base foliar». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 9 de diciembre de 2010. Consultado el 10 de septiembre de 2010.
- Font Quer, P. (1982). Diccionario de Botánica. 8ª reimpresión. Barcelona: Editorial Labor, S. A. 84-335-5804-8.
- Nordhausen, M. (1930). Morfología y organografía de las plantas. Barcelona: Editorial Labor, S. A.
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema 2: Hoja. Base foliar y estípulas». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 9 de diciembre de 2010. Consultado el 10 de septiembre de 2010.
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema 2: Hoja, 2.9. Prefoliación o Vernación». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 12 de marzo de 2009. Consultado el 10 de septiembre de 2010.
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema 2: Hoja, 2.8. Filotaxis y vernación». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 12 de marzo de 2009. Consultado el 10 de septiembre de 2010.
- Judd, W. S.; Campbell, C. S.; Kellogg, E. A.; Stevens, P. F.; Donoghue, M. J. 2002. Plant systematics: a phylogenetic approach, Second Edition. Sinauer Axxoc, USA.
- Cabrera, A.L. (Ed.) 1977. Flora de la provincia de Jujuy. Parte II: Pteridophyta. Colección Científica, Instituto Nacional de Tecnología Agropecuaria, Tomo XIII.
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema 21:ANATOMÍA FOLIAR». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 24 de febrero de 2011. Consultado el 10 de septiembre de 2010.
- Louguet P., CoudretA., Couot-Gastelier J., Lasceve G. 1990. Structure and ultrastructure of stomata. Biochem.Physiol.Pflanzen 186: 273-287
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema 13: Epidermis 13.4. Estomas». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 10 de abril de 2009. Consultado el 10 de septiembre de 2010.
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema 13: Epidermis, 13.5. Células Oclusivas». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 31 de julio de 2009. Consultado el 10 de septiembre de 2010.
- Sack F.D. 1994. Structure of the stomatal complex of the monocot Flagellaria indica. Amer. J. Bot. 81: 339-344
- Heath, O.V.S. 1981. Stomata. Carolina Biological Supply, Scientific Publications Division.
- Willmer C.M. & R. Sexton. 1979. Stomata and Plasmodesmata. Protoplasma 100: 113-124
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema 13: Epidermis, 13.7. Tricomas». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 27 de febrero de 2009. Consultado el 10 de septiembre de 2010.
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema 14: Estructuras glandulares». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 26 de agosto de 2010. Consultado el 10 de septiembre de 2010.
- Fahn, A. 1990. Plant Anatomy, 4th ed. Pergamon Press
- Heslop-Harrison, Y. 1976. Carnivorous plants a century after Darwin. Endeavour 35(126):114-122
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Glándulas de plantas carnívoras». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 31 de octubre de 2009. Consultado el 10 de septiembre de 2010.
- Legendre L (2000). "The genus Pinguicula L. (Lentibulariaceae): an overview". Acta Botanica Gallica 141 (1): 77–95.
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema14: ESTRUCTURAS GLANDULARES, 14.8. Osmóforos e Hidátodos». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 19 de junio de 2009. Consultado el 10 de septiembre de 2010.
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema 11: Parénquima 11.2. Parénquima clorofiliano o clorénquima». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 16 de julio de 2010. Consultado el 10 de septiembre de 2010.
- González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema 21:ANATOMÍA FOLIAR 21.2. HOJAS DE DICOTILEDÓNEAS, 2) SISTEMA FUNDAMENTAL O MESÓFILO». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 8 de julio de 2009. Consultado el 10 de septiembre de 2010.
- Benjamin Cummins (2007), Biological Science (3 edición), Freeman, Scott, p. 215.
- Swarthout D. & C.M. Hogan. 2010. Stomata. Encyclopedia of Earth. National Council for Science and the Environment, Washington DC
- Martin, J.; Leonard, W.; Stamp, D. (1976), Principles of Field Crop Production (Third Edition), New York: Macmillan Publishing Co., Inc., ISBN 0-02-376720-0.
- Universidad Politécnica de Valencia. «La Fotosíntesis». Consultado el 5 de diciembre de 2009.
- Field CB, Behrenfeld MJ, Randerson JT, Falkowski P (1998). «Primary production of the biosphere: integrating terrestrial and oceanic components». Science 281. 237 - 240.
- “Photosynthesis,” McGraw-Hill Encyclopedia of Science and Technology, Vol. 13, 2007
- Heal, O. W.; Maclean, S. F. 1980. productividad comparada en los ecosistemas: productividad secundaria. En: Conceptos unificadores en ecología. Dobben, W. H. et al. (edits.). Editorial Blume, primera edición, Barcelona, España.
- Feild T. S., D. W. Lee, N. M. Holbrook (2001). «Why leaves turn red in autumn. The role of anthocyanins in senescing leaves of red-osier dogwood» (PDF). Plant Physiol 127 (2): 566-574. Archivado desde el original el 20 de julio de 2008.
- G. Mazza, E. Miniati: Anthocyanins in fruits, vegetables, and grains. CRC Press, Boca Raton 1993. ISBN 0-8493-0172-6
- Niklas, Karl J. (1997). The Evolutionary Biology of Plants. University of Chicago Press. ISBN 0-226-58083-0.
- Gifford EM, Foster AS. 1988. Morphology and evolution of vascular plants. New York, NY, Freeman.
- Field CB, Behrenfeld MJ, Randerson JT, Falkowski P 1998. Primary production of the biosphere: integrating terrestrial and oceanic components. Science 281: 237–240.
- Kidston, R., & Lang, W. H. (1920). On old red sandstone plants showing structure, from the Rhynie Chert bed, Aberdeenshire: Part III. Asteroxylon mackiei. Royal Society of Edinburgh Transactions 52: 643-680.
- Edwards D, Wellman C. 2001. Embryophytes invade the land. In: Gensel PG, Edwards D, eds. Plants invade the land. New York, NY: Columbia University Press, 3–28.
- Kenrick P, Crane PR. 1997. The origin and early evolution of plants on land. Nature 389: 33–39.
- Boyce CK, Knoll AH. 2002. Evolution of developmental potential and the multiple independent origins of leaves in Paleozoic vascular plants. Paleobiology 28: 70–100.
- Osborne CP, Beerling DJ, Lomax BH, Chaloner WG. 2004a. Biophysical constraints on the origin of leaves inferred from the fossil record. Proceedings of the National Academy of Sciences of the USA 101: 10360–10362.
- Osborne CP, Chaloner WG, Beerling DJ. 2004b. Falling atmospheric CO2 – the key to megaphyll leaf origins. In: Poole I, Hemsley AR. The evolution of plant physiology. Kew: Elsevier Academic Press, 197–215
- Hao SG, Beck CB. 1993. Further observations on Eophyllophyton bellum from the lower Devonian (Siegenian) of Yunnan, China. Palaeontographica 230: 27–47.
- Hao S, Beck CB, Deming W. 2003. Structure of the earliest leaves: adaptations to high concentrations of CO2. International Journal of Plant Sciences 164: 71–75.
- Stewart, W.N.; Rothwell, G.W. (1993). Paleobiology and the evolution of plants. Cambridge University Press. pp. 521pp.
- Crane, P.R.; Herendeen, P. & Friis, E.M. (2004), «Fossils and plant phylogeny», American Journal of Botany 91: 1683-99, doi:10.3732/ajb.91.10.1683, archivado desde el original el 18 de julio de 2010, consultado el 16 de junio de 2011.
- Taylor, T.N.; Taylor, E.L. & Krings, M. (2009), Paleobotany, The Biology and Evolution of Fossil Plants (2nd edición), Amsterdam; Boston: Academic Press, ISBN 978-0-12-373972-8., p. 227
- Rickards, R.B. (2000). «The age of the earliest club mosses: the Silurian Baragwanathia flora in Victoria, Australia» (abstract). Geological Magazine 137 (2): 207-209. doi:10.1017/S0016756800003800. Consultado el 25 de octubre de 2007.
- Kaplan, D.R. (2001). «The Science of Plant Morphology: Definition, History, and Role in Modern Biology». American Journal of Botany 88 (10): 1711-1741. doi:10.2307/3558347. Consultado el 31 de enero de 2008.
- Taylor, T.N.; Hass, H.; Kerp, H.; Krings, M.; Hanlin, R.T. (2005). «Perithecial ascomycetes from the 400 million year old Rhynie chert: an example of ancestral polymorphism» (abstract). Mycologia 97 (1): 269-285. PMID 16389979. doi:10.3852/mycologia.97.1.269. Consultado el 7 de abril de 2008.
- Boyce, C.K.; Knoll, A.H. (2002). «Evolution of developmental potential and the multiple independent origins of leaves in Paleozoic vascular plants». Paleobiology 28 (1): 70-100. doi:10.1666/0094-8373(2002)028<0070:EODPAT>2.0.CO;2.
- Hao, S.; Beck, C.B.; Deming, W. (2003). «Structure of the Earliest Leaves: Adaptations to High Concentrations of Atmospheric CO2». International Journal of Plant Sciences 164 (1): 71-75. doi:10.1086/344557.
- Berner, R.A.; Kothavala, Z. (2001). «Geocarb III: A Revised Model of Atmospheric CO2 over Phanerozoic Time» (abstract). American Journal of Science 301 (2): 182. doi:10.2475/ajs.301.2.182. Archivado desde el original el 29 de enero de 2009. Consultado el 7 de abril de 2008.
- Beerling, D.J.; Osborne, C.P.; Chaloner, W.G. (2001). «Evolution of leaf-form in land plants linked to atmospheric CO2 decline in the Late Palaeozoic era». Nature 410 (6826): 287-394. PMID 11268207. doi:10.1038/35066546.
- PEREZ, Martha y REBOLLAR, Silvia. Anatomía y usos de las hojas maduras de tres especies de Sabal (Arecaceae) de la Península de Yucatán, México. Rev. biol. trop [online]. 2003, vol.51, n.2, pp. 333-344. ISSN 0034-7744.
- Joyal, E. 1996. The use of Sabal uresana (Arecaceae) and other palms in Sonora, México. Econ. Bot. 50: 429-445
- Laming, E. 1967. Perú before the Incas. Englewood Cliffs, NJ: Prentice-Hall.
- Martin, R. T. 1970. The Role of Coca in the History, Religion and Medicine of South American Indians. Economic Botany 24:422-434.
- Amerindia. Artesanía Archivado el 21 de junio de 2012 en Wayback Machine.. Universidad Nacional de Táchira. Consultado 1 de junio 2012.
- World Vegetables: Principles, Production, and Nutritive Values; Second Ed. by Vincent E. Rubatzky and Mas Yamaguchi.
Enlaces externos
 Wikimedia Commons alberga una galería multimedia sobre Hoja.
Wikimedia Commons alberga una galería multimedia sobre Hoja. Wikcionario tiene definiciones y otra información sobre hoja.
Wikcionario tiene definiciones y otra información sobre hoja.