Cuerpo basal
Un cuerpo basal o cinetosoma es una estructura que se presenta en la base de las prolongaciones móviles de los eucariotas (cilios o flagelos) y que sirve como punto de agregación para el crecimiento y el ordenamiento de los microtúbulos que componen el axonema.
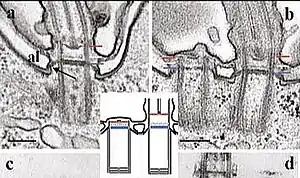
Los cuerpos basales y los centríolos son dos versiones del mismo orgánulo eucariota y ambos comparten dos propiedades: una simetría de nueve veces (nónuple) del eje de los microtúbulos y una capacidad para generar un nuevo orgánulo en una relación con el orgánulo "madre".[1] [2] [3]
Los defectos en el posicionamiento correcto del cuerpo basal, causan una variedad de síntomas de enfermedad que incluyen: hidrocefalia, situs inversus e infecciones, del oído medio y respiratorias.[4] [5]
Estructura del cuerpo basal
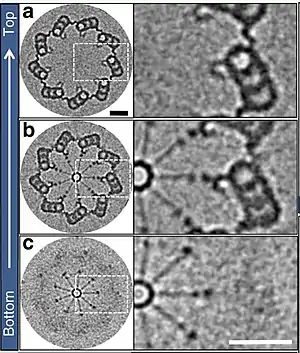
Los cuerpos basales son estructuralmente iguales, cada uno de ellos contiene una configuración helicoidal de microtúbulos en disposición de [9(3)+0] tripletes (9 grupos de 3 túbulos exteriores sin microtúbulos centrales) formando un cilindro hueco.
Los cuerpos basales tienen un diámetro promedio de 260 nm y una longitud de triplete cercana a 400 nm.[6]
En una escala comparativa: si establecemos el tamaño del ribosoma en una pelota de tenis, entonces el centríolo sería del tamaño de un tambor (barril) de 200 litros.[7]
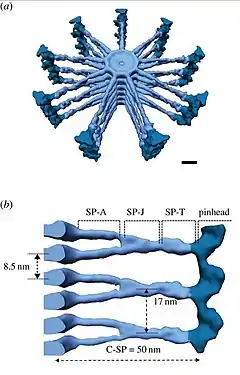
Los componentes del cuerpo basal se localizan en "dominios discretos" de proteínas. Los dominios basales que residen en el "disco generativo" o disco amorfo y en la "rueda de carro", son componentes para el ensamblaje temprano del nuevo cuerpo basal.[2][8]
La estructura completa de la "rueda de carro" es típicamente 70-100 nm de altura y ubicado en la parte proximal del cuerpo basal.
Nueve radios de aproximadamente 50 nm de largo, emanan del eje central (" hub") de ~22 nm de diámetro y se irradian hacia la periferia de la rueda, donde los radios que se originan de dos anillos superpuestos se fusionan. Cada radio se conecta al túbulo A del triplete de microtúbulos a través de una estructura abultada llamada "cabeza de alfiler" (pinhead) produciendo un espacio vertical de 17-20 nm entre dos ruedas.[9]
[10]
Las proteínas que se localizan en los dominios distales del cuerpo basal, están involucradas en la formación y función ciliar.[2]
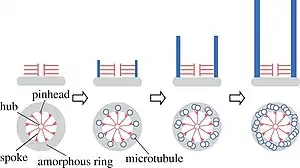

Los cuerpos basales, formados a partir de los centriolos, actúan como puntos de anclaje para las proteínas, que a su vez anclan los microtúbulos como un centro organizador de microtúbulos (MTOC en inglés), proporcionan la estructura, facilitan el movimiento de las vesículas en la célula eucariota y centran la formación de cilios y flagelos.[11]
Origen del cuerpo basal
Todos los cuerpos basales se desarrollan a partir de un cuerpo basal "materno"
preexistente. Se denominan así porque se ensamblaron en un ciclo celular anterior al actual, llamado "centríolo hijo".
Primero, los centriolos deben migrar a la región apropiada en la superficie celular en la que se fijarán y comenzarán a ensamblar los cilios. En segundo lugar, una vez que los centríolos alcanzan la superficie celular, deben orientarse con una dirección adecuada para los cilios móviles, o de modo que estén orientados para participar en la señalización como en el caso de un cilio primario.[12]
En las células de paramecium, durante la interfase uno o dos cuerpos basales están anclados permanentemente en la superficie celular en medio de cada unidad cortical e incrustados en la capa citoesquelética superficial llamada epiplasma.[13]
En los vertebrados el "cilio primario" es un derivado del centríolo materno, uno de los dos centríolos que organizan el centrosoma.[1] [14]
Cuerpo basal en Paramecium
Las unidades corticales de Paramecium contienen uno o dos cuerpos basales, con una distribución específica según la zona.
Coexisten varios tipos de unidades corticales que contienen cuerpos basales en diferentes etapas del proceso de duplicación: no duplicado, duplicado y completamente ensamblado.[14]
Todos los cuerpos basales de paramecios muestran una organización similar, pero su longitud individual varía entre 330-600 nm dependiendo de la zona de la corteza celular en la que se ubican.[13]
Cuerpo basal mamífero
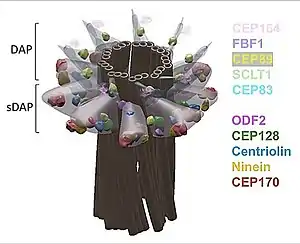
Los cuerpos basales (BB) mamíferos, son cilindros generalmente construidos a partir de nueve tripletes de microtúbulos (M.T.), organizados en una configuración simétrica repetida nueve veces: una estructura llamada rueda de carreta.[15] Los mamíferos tienen una región proximal de 200 nm donde el triplete de túbulos es el estado predeterminado, los conectores de los túbulos A-C (A-C linkers), la cabeza del alfiler y las estructuras de los cuerpos basales.[16]
Tomados en conjunto, estos cuerpos basales (BB) están estructurados con:
- la rueda de carreta con nueve radios,
- nueve microtúbulos triples que forman el barril del cuerpo basal y que se denominan A, B y C,
- la matriz proteica periférica o satélite compuesta por proteínas como la pericentrina, que nuclea los extremos de muchos microtúbulos citoplasmáticos,
- los apéndices subdistales (SAP en inglés) o pies basales (BF) en número de tres a nueve, se proyectan desde los lados del cuerpo basal, en la posición de 350 nm desde el extremo proximal del cuerpo basal.
- apéndices distales (DAP en inglés) también llamadas láminas alares, fibras de transición. Los nueve apéndices distales se proyectan hacia afuera desde el extremo distal del cuerpo basal, son necesarios para el acoplamiento de membrana y la cilogénesis.
- la raicilla ciliar.[17] [18] [19] [20]
Los cuerpos basales (BB) humanos contienen cinco tipos de tubulina: α, β, γ, δ y ε.
Los polímeros de los microtúbulos (M.T) consisten exclusivamente en heterodímeros de tubulina α y β.
La γ-tubulina está integrada en los complejos de anillo de gamma tubulina (γ-TURCs), que son responsables de nucleación de los MT.
La ε-tubulina se asocia con los apéndices subdistales (SAP) y es fundamental para la duplicación de centriolos y la organización de microtúbulos.[18]
La regulación de la producción del cuerpo basal y su orientación espacial es una función del dominio de enlace de los nucleótidos de la γ-tubulina (Shang et al, 2005).[21]
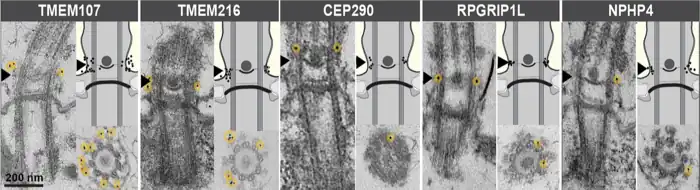
Zona de transición TZ. Placa axosómica indicada por una punta de flecha.
Véase también
Referencias
- Megías M. «5._Cuerpos basales. Cilios y flagelos.». Atlas de histología vegetal y animal. Universidad de Vigo. Consultado el 23 de abril de 2020.
- Pearson C.G., Winey M. (2009). «Basal Body Assembly in Ciliates: The Power of Numbers». Traffic 10 (5). Consultado el 21 de abril de 2020.
- Carvalho-Santos Z., Azimzadeh J., Pereira-Leal J.B., Bettencourt-Dias M. (2011). «Tracing the origins of centrioles, cilia, and flagella». J Cell Biol (Revisión) 194 (2): 165-175. Consultado el 21 de abril de 2020.
- Ross M.H., Pawlina W. (2007). «5:Tejido epitelial». Histología: Texto y Atlas. Médica Panamericana. pp. 116-118. Consultado el 7 de mayo de 2020.
- O'Toole E.T., Dutcher S.K. (2013). «Site‐specific basal body duplication in Chlamydomonas». Cytoskeleton 71 (2). Consultado el 6 de mayo de 2020.
- Li S., Fernández J.J., Marshall W.F., Agard D.A. (2012). «Three‐dimensional structure of basal body triplet revealed by electron cryo‐tomography». EMBO J. 31: 552-562. Consultado el 1 de mayo de 2020.
- Winey M., O'Toole E. (2014). «Centriole structure». Philosophical Transactions of the Royal Society, Biological Sciences (Revisión) 369 (1650). PMID 25047611. Consultado el 1 de mayo de 2020.
- Guichard P., Hamel V., Gönczy V. (2018). «The Rise of the Cartwheel: Seeding the Centriole Organelle». BioEssays 40 (4). Consultado el 12 de mayo de 2020.
- Guichard P., Hamel V., Le Guennec M., Banterle N., Iacovache I., Nemčíková V., Flückiger I., Goldie K.N., Stahlberg H., Lévy D., Zuber B., Gönczy P. (2017). «Cell-free reconstitution reveals centriole cartwheel assembly mechanisms». Nature Communications 8 (14813). Consultado el 3 de mayo de 2020.
- Hirono M. (2014). «Cartwheel assembly». Philos Trans R Soc Lond B Biol Sci. 5 (369(1650)): 20130458. Consultado el 3 de mayo de 2020.
- Azimzadeh J., Marshall W.F. (2010). «Building the Centriole». Curr Biol. (Revisión) 20 (18): R816-R825. PMC 2956124. PMID 20869612. doi:10.1016/j.cub.2010.08.010. Consultado el 23 de abril de 2020.
- Feldman J.L., Geimer S., Marshall W.F. (2007). «The Mother Centriole Plays an Instructive Role in Defining Cell Geometry». PLoS Biology 5 (6): e149. Consultado el 24 de abril de 2020.
- Tassin A-M., Lemullois L., Aubusson-Fleury A. (2016). «Paramecium Tetraurelia Basal Body Structure». Cilia (Revisión) 5 (6). PMID 28367320. doi:10.1186/s13630-017-0050-z. Consultado el 22 de abril de 2020.
- Bengueddach H., Lemullois M., Aubusson-Fleury A., Koll F. (2017). «Basal body positioning and anchoring in the multiciliated cell Paramecium tetraurelia: roles of OFD1 and VFL3». Cilia 6 (6). Consultado el 23 de abril de 2020.
- Anderson, R.G.W. (1972). «The three-dimensional structure of the basal body from the rhesus monkey oviduct». J. Cell Biol. (pdf) 54: 246-265. Consultado el 29 de abril de 2020.
- Greenan G.A., Keszthelyi B., Vale R.D., Agard D.A. (2018). «Insights into centriole geometry revealed by cryotomography of doublet and triplet centrioles». eLife 7. Consultado el 7 de mayo de 2020.
- Tateishi K., Yamazaki Y., Nishida T., Watanabe S., Kunimoto K., Ishikawa H., Tsukita S. (2013). «Two appendages homologous between basal bodies and centrioles are formed using distinct Odf2 domains». J Cell Biol 203 (3): 417-425. Consultado el 30 de abril de 2020.
- Vertii A., Hung H., Hehnly H. (2016). «Human basal body basics». Cilia (Revisión) 5 (13). Consultado el 26 de abril de 2020.
- Uzbekov R., Alieva I. (2018). «Who are you, subdistal appendages of centriole?». Open Biol (Revisión) 8 (7): 180062. PMID 30045886. Consultado el 1 de mayo de 2020.
- Galo Garcia III & Jeremy F. Reiter (2016). «A primer on the mouse basal body». Cilia (Revisión). 5, (17). Consultado el 7 de mayo de 2020.
- Y. Shang, C.-C. Tsao, M. A. Gorovsky. (2005). «Mutational analyses reveal a novel function of the nucleotide-binding domain of gamma-tubulin in the regulation of basal body biogenesis». J. Cell Biol. 171 (6): 1035-1044. PMID 16344310.