Corteza auditiva primaria
La corteza auditiva primaria o A1 es la región del cerebro humano responsable del procesamiento de la información auditiva. Se localiza en el lóbulo temporal, concretamente en las circunvoluciones transversas o área de Heschl.[1][2] Aproximadamente, se corresponde con las antiguas áreas de Brodmann 41 y 42.
Además de recibir la información que le llega desde los oídos y los centros basales del cerebro, la corteza auditiva primaria también devuelve señales hacia estas áreas.[3]
| Corteza auditiva primaria | ||
|---|---|---|
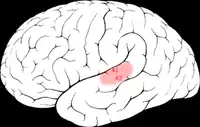 Áreas 41 y 42 de Brodmann en el cerebro humano | ||
| Es parte de |
Lóbulo temporal (cerebro) Sistema auditivo central | |
| Arterias |
Arteria cerebral media Arteria temporal posterior | |
| Sinónimos | ||
Áreas 41 y 42 z(1909) Áreas TC y TB (1925) Área auditiva A1 (1997) Áreas KAm y KAlt (1980) Área Te1.0 (2001) | ||
| Enlaces externos | ||
| Gray | pág.824 | |
| NeuroNames | Primary auditory cortex | |
Estructura
La corteza cerebral auditiva primaria (A1) se localiza en el lóbulo temporal. Su ubicación y estructura exacta varían entre los primates. Fueron necesarios estudios estructurales y funcionales para su completa caracterización.
Macro-estructura
Se ubica sobre la parte posterior de la circunvolución temporal superior, dentro de las denominadas circunvoluciones temporales transversas, concretamente, en el área de Heschl.[4][5]
En los cerebros humanos, cuando está presente solamente un área de Heschl, el núcleo visual se limita a esta área y ocupa sus tercios medial y central.
La corteza auditiva primaria (A1) ocupa aproximadamente la mitad del volumen total del área de Heschl.[6][7]
El grosor de la corteza sensorial humana en la superficie del giro temporal transverso, puede ser mayor de 3 mm en la corteza auditiva primaria (A1).[8]
Micro-estructura
El núcleo auditivo A1, se diferencia por ser una zona altamente celular de aspecto granular llamada koniocorteza.[9][10]
La corteza auditiva primaria (Al) presenta las capas II y IV densas y uniformes. Este núcleo tiene una capa celular IV bien desarrollada, debido a una aferencia densa de la parte auditiva del tálamo. La capa celular III del núcleo auditivo se caracteriza por la presencia de células piramidales de tamaño pequeño a mediano.
- Quimio-arquitectura
La corteza auditiva primaria A1, se caracteriza por presentar una alta actividad de acetilcolinesterasa (AChE) y citocromooxidasa (CO).[9]
La citocromo oxidasa o la Dihidrolipoil deshidrogenasa (NADH diaforasa) se concentran en la capa IV de las áreas sensoriales primarias, incluida la A1.[8]
- Mielo-arquitectura
Se ha utilizado el estudio de la densidad de localización de las fibras de mielina como indicador de ubicación de A1. Las áreas que reciben las grandes proyecciones sensoriales están fuertemente mielinizadas, y la mielinización disminuye con la distancia de estas áreas.
El área auditiva primaria se puede identificar en imágenes de resonancia magnética (RM) debido a su alto contenido de mielina.
La imagen de IRM estructural in vivo ya se está utilizando para distinguir entre el área auditiva primaria con sus altos niveles de mielina en las capas profundas y áreas adyacentes como LP, que tienen niveles mucho más bajos de mielina.[8]
En el área de Herschl medial, las secciones de Vogts-Hopf 146, 148 y 151 reflejan la corteza visual primaria (PAC) humana.[11]

En los primates, se pueden identificar tres sectores principales en esta corteza, que generalmente se denominan núcleo (core), cinturón interno (belt) y faja externa (parabelt).[4]
En la corteza auditiva primaria humana se sugieren dos sectores dentro del núcleo (core): el sector A1 y el sector R (rostral).[6]
Varios campos menos granulares rodean al núcleo auditivo primario. En estos campos periféricos el empaquetamiento de células es menos denso que en el núcleo, y las células piramidales en la capa III son más grandes y más numerosas.
Un área con una estructura granular reducida llamada parakoniocortex, con grandes neuronas piramidales en la capa IIIc se ha informado en el "cinturón externo" (parabelt).[8]
Existen otras áreas de la corteza cerebral involucradas en el procesamiento del sonido, en el lóbulo frontal y el lóbulo parietal. Se han realizado estudios con animales que indican que las zonas auditivas de la corteza cerebral reciben un input ascendente procedente del tálamo auditivo. Estas zonas están interconectadas entre sí y con las del hemisferio cerebral opuesto. Así, la corteza auditiva se compone de diferentes zonas que se diferencian tanto por su estructura como por su función.[12]
Función
Al igual que ocurre en otras áreas de la corteza primaria sensorial, las sensaciones auditivas solamente desencadenan el proceso de percepción sonora si son recibidas y procesadas en una zona de corteza cerebral. Este fenómeno se ha comprobado mediante estudios de lesiones en pacientes humanos que presentaban daños importantes en áreas corticales, debidos a un tumor cerebral o un accidente cerebrovascular.
- También se han realizado experimentos con animales a los que se les desactivaban determinadas áreas corticales mediante técnicas de enfriamiento, o la aplicación local de fármacos.
En los seres humanos, los daños en la corteza auditiva primaria provocan una pérdida de conciencia del sonido, aunque se conserva la capacidad de reaccionar de forma refleja a los sonidos, ya que existe una gran cantidad de procesamiento subcortical que tiene lugar a nivel del tronco del encéfalo y del mesencéfalo

Las neuronas de la corteza auditiva están organizadas según la frecuencia de los sonidos a los que responden con mayor eficacia. Las neuronas situadas en un extremo de la corteza auditiva responden mejor a las frecuencias bajas (2 Hz), y las ubicadas en el otro extremo, responden mejor a las frecuencias más altas (128 Hz).[13] Existen múltiples áreas auditivas, que pueden distinguirse anatómicamente, y que conforman un completo "mapa de frecuencias".[14] El propósito de este mapa de frecuencias (conocido como mapa tonotópico) es desconocido, y parece reflejar el hecho de que la cóclea está organizada asimismo para responder selectivamente ante las diferentes frecuencias sonoras. La corteza auditiva está implicada en tareas como la identificación y diferenciación de "objetos" auditivos, así como en la localización espacial de un sonido.
Algunos estudios realizados mediante técnicas de escáner cerebral han mostrado que una pequeña región periférica de esta área cerebral se activa al intentar identificar la altura de un sonido. Las células individuales son excitadas consistentemente por determinados sonidos emitidos a unas frecuencias específicas.
La corteza auditiva es una parte importante, aunque ambigua, del proceso auditivo. No está totalmente claro lo que ocurre exactamente cuando los impulsos sonoros llegan a la corteza. El músico y científico James Beament escribió al respecto: «La corteza es tan complicada que lo máximo a lo que podemos aspirar es a conocerla superficialmente, dado que las pruebas de que disponemos en la actualidad sugieren que no existen dos cortezas que actúen exactamente del mismo modo».[15]
En el proceso de audición se reciben múltiples sonidos simultáneamente. El papel del sistema auditivo es el de decidir qué componentes conforman el sonido y desdeñar el resto. Se ha sospechado que la unión de estos sonidos relevantes se basa en la localización de la procedencia de los sonidos; no obstante, existen numerosas distorsiones de los sonidos cuando rebotan en diferentes medios, lo que hace de esta teoría algo poco probable. Una teoría alternativa es que la corteza auditiva agrupa los sonidos basándose en otro tipo de información más fiable. Por ejemplo, en lo que a percepción musical se refiere, influirían la armonía y la altura de los sonidos, entre otros factores.[16]
El número de zonas varía según las especies; desde las dos de los roedores hasta las 15 del mono rhesus. El número, localización y organización de las áreas de la corteza auditiva humana no se conoce exactamente a día de hoy. Todo lo que se conoce acerca de la corteza auditiva humana proviene de los estudios realizados con mamíferos, incluyendo primates. Estos datos se usan para interpretar los estudios realizados mediante tests electrofisiológicos técnicas de neuroimagen funcional con seres humanos.
Cuando cada instrumento de una orquesta sinfónica o de una banda de jazz tocan la misma nota, las características de cada sonido son diferentes, aunque los músicos perciban que todas ellas tienen la misma tonalidad. Las neuronas de la corteza auditiva son capaces de responder ante una tonalidad determinada. Se han realizado estudios con monos que muestran que las neuronas que responden específicamente ante determinadas tonalidades se localizan en una región cortical cercana al borde anterolateral de la corteza auditiva primaria. Recientemente, se ha podido encontrar este mismo tipo de neuronas específicas de tonalidad en seres humanos, mediante técnicas de neuroimagen funcional.[17][18] La corteza auditiva primaria se ve modulada por numerosos neurotransmisores como la norepinefrina, que reduce la excitabilidad celular en todas las capas de la corteza temporal. La norepinefrina reduce el potencial excitatorio postsináptico glutamatérgico en los receptores AMPA mediante la activación de los receptores adrenérgicos Alfa 1.[19]
Áreas 41 y 42 de Brodmann
En la investigación, un primer enfoque fue dividir la corteza en áreas anatómicas pequeñas de acuerdo con su microestructura histológica, como las áreas de Brodmann.[6]
El área de Brodmann 41 también se conoce como área temporal transversal anterior 41. Es una subdivisión de la superficie dorsal del lóbulo temporal de la corteza cerebral. Esta comprendida en el
giro temporal transverso anterior de la circunvolucion temporal superior. El área 41 de Brodmann está unida por su parte medial (interna) al área parainsular 52, y por su parte lateral (externa) al área 42, (áreas de Brodmann, 1909).
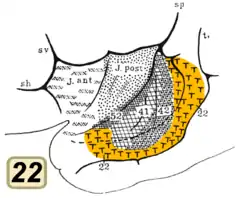
El área 42 también se conoce como área temporal transversal posterior. Es una subdivisión de la región temporal, localizada junto al surco lateral, en la superficie dorsal del lóbulo temporal. El área 42 está unida por su parte medial (interna) al área temporal transversal anterior (41), y por su parte lateral (externa) está unida al giro temporal superior (área 22) (Brodmann, 1909).
Relaciones con el sistema auditivo
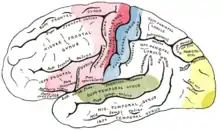
La corteza auditiva es la unidad cerebral de procesamiento de sonidos más organizada. Esta área cortical es crucial para la audición, y en los seres humanos, también para el lenguaje y la música. La corteza auditiva se divide en tres partes diferenciadas: las cortezas auditivas primaria, secundaria y terciaria. Estas estructuras se disponen de manera concéntrica, encontrándose la corteza primaria en el centro, y la terciaria en la periferia.
La corteza auditiva primaria está tonotópicamente organizada, lo que significa que las células de la corteza que son vecinas entre sí, responden a frecuencias similares.[20] Esta propiedad se manifiesta a lo largo de la mayor parte del circuito de audición. Se cree que la corteza auditiva se encarga de identificar los elementos básicos de la música, como el volumen o el tono. Esta hipótesis es la más lógica, dado que es el área que recibe un input directo del núcleo geniculado medial del tálamo. Se ha indicado que la corteza auditiva secundaria está implicada en el procesamiento de patrones rítmicos, melódicos y armónicos. Por último, se supone que la corteza auditiva terciaria es la encargada de integrar toda la información hasta crear la experiencia global de la percepción musical.[21]
Klinke et al. realizaron un estudio de respuestas evocadas sobre gatos con ceguera congénita para medir la plasticidad de la corteza auditiva. Los gatos eran estimulados y comparados con un grupo control de gatos cogénitamente ciegos que no recibían estimulación, y otro grupo control de gatos sanos. Los potenciales medidos de los gatos artificialmente estimulados fueron mucho más fuertes que los obtenidos en el caso de los gatos sanos.[22] Estos resultados concuerdan con los obtenidos por Eckart Altenmuller en un estudio en el que observó que aquellos estudiantes que recibían clases de música mostraban una mayor activación cortical que aquellos que no las recibían.[23]
La corteza auditiva muestra un extraño comportamiento en cuanto a las frecuencias de ondas gamma. cuando los sujetos se exponen a tres o cuatro ciclos de 40 hertzios, aparece un pico anormal en el electroencefalograma que no se observa durante la presentación de cualquier otro estímulo. Este pico en la actividad neuronal asociado a una frecuencia determinada no está restringido a la organización tonotópica de la corteza auditiva. Se ha teorizado que existe una "frecuencia de resonancia" para ciertas áreas del cerebro, y parece afectar igualmente a la corteza visual.[24] Se ha encontrado una activación de la banda gamma (entre 20 y 40 hertzios) durante la percepción de eventos sensoriales, y durante el proceso de reconocimiento. Kneif et al., en un estudio del año 2000, presentaron a los participantes ocho notas musicales de canciones muy conocidas, como Yankee Doodle y Frère Jacques. La sexta y la séptima nota eran omitidas al azar, y se empleó un electroencefalograma y un magnetoencefalograma para medir los resultados neuronales. Más concretamente, se medía la presencia de ondas gamma inducidas por la tarea auditiva. La respuesta a la omisión del estímulo (ROE) se localizó en una posición ligeramente diferente. Además, las grabaciones de la ROE fueron más bajas, en comparación con las obtenidas durante la ejecución completa del conjunto de las notas musicales. Se asumió que las respuestas evocadas emitidas durante la sexta y la séptima notas omitidas fueron imaginadas, y eran característicamente diferentes, especialmente en el hemisferio derecho.[25] La corteza auditiva derecha se ha mostrado más sensible a la tonalidad, mientras que la izquierda parece responder en mayor medida a las diferencias secuenciales que se presentan en el sonido, como por ejemplo durante el transcurso de un discurso.
Se ha comprobado que las alucinaciones producen oscilaciones paralelas (aunque no exactamente iguales) al rango de frecuencia gamma. Sperling demostró en un estudio realizado en el año 2004 que las alucinaciones auditivas producen bandas formadas por ondas entre el rango de 12,5 a 30 hertzios. Estas bandas aparecían en la corteza auditiva izquierda de sujetos esquizofrénicos. Estos resultados concuerdan con los obtenidos en investigaciones realizadas con sujetos a los que se les pedía que recordaran mentalmente una canción. En estos casos, los sujetos no percibían sonido alguno, pero experimentaban mentalmente la melodía, el ritmo, y en suma, la experiencia global del sonido. Cuando los esquizofrénicos experimentan alucinaciones, se activa la corteza primaria auditiva. Este fenómeno es característicamente diferente de recordar un estímulo sonoro, que solamentelo produce una débil activación de la corteza auditiva terciaria.[21]
La percepción de la característica musical del tono no solamente tiene lugar en la corteza auditiva; existe un área especialmente interesante a este respecto: la corteza prefrontal rostromedial.[26] Janata et al., en un estudio realizado en el año 2002, exploraron las áreas cerebrales que se activan durante el procesamiento de la tonalidad, mediante la técnica de imagen por resonancia magnética funcional. Los resultaron mostraron la activación de varias áreas que usualmente no se consideran parte del proceso auditivo. La corteza prefrontal rostro medial es una subsección de la corteza prefrontal medial, que se proyecta hacia la amígdala, y se cree que ayuda al proceso de inhibición de las emociones negativas.[27] La corteza prefrontal rostromedial es sensible a la tonalidad, lo que significa que se ve activada por los tonos y frecuencias producidos por los sonidos y la música.
Desarrollo
Al igual que ocurre con otras muchas áreas del neocórtex, las propiedades funcionales de la corteza primaria auditiva adulta (A1) dependen en gran medida de los sonidos percibidos durante la infancia. Esto se ha estudiado principalmente con modelos animales, especialmente gatos y ratas.
- En la rata, la exposición a una frecuencia única durante los días 11 a 13 a partir del nacimiento puede provocar una expansión en la representación de esa frecuencia en la corteza auditiva.[28]
Este cambio es persistente, ya que permanece a lo largo de toda la vida del animal, y es específico, ya que si la misma exposición a esa frecuencia se realiza fuera de este periodo concreto, no se producen cambios en el mapa tonotópico de corteza.
Disfunción
Los efectos del funcionamiento anómalo o una lesión de la corteza auditiva son estudiadas por la medicina y la psicología.[29]
En la mayoría de los adultos, los centros del lenguaje están situados en el hemisferio izquierdo (dominancia).
- Corteza auditiva primaria:
las lesiones unilaterales del núcleo auditivo son subclínicas. Las lesiones bilaterales de la corteza primaria provocan sordera cortical.[30][29]
- Cortezas auditivas de asociación:
- En el Hemisferio izquierdo
- Afasia sensorial de Wernicke
- Afasia transcortical sensorial
- Afasia anómica
- Afasia de conducción acústico-amnésica
- Amnesia para material verbal
- Amusia y Agnosia auditiva[30]
- En el Hemisferio derecho
- Amusia receptiva o sensorial
- Disprosodia sensorial
- Amnesia para material visual
- Bilaterales
- Agnosia auditiva
- Sordera verbal pura[30]
La afasia de Wernicke es sensorial y afecta las imágenes auditivas de las palabras. Se presenta por una lesión izquierda temporal-parietal. En esta afasia, la comprensión es lo más alterado, siendo la fluencia normal. Sin embargo el contenido del lenguaje de estos pacientes también está alterado en la forma que a veces se ha denominado "ensalada de palabras" (las palabras están bien pronunciadas pero su contenido solo se ajusta parcialmente a la gramática y objetivo comunicativo del sujeto).
Véase también
Referencias
- Anatomía de Gray, «The Fore-brain or Prosencephalon». p. 825.
- Peña-Casanova, J. (2007). Neurología de la conducta y neuropsicología. Madrid: Editorial médica panamericana. p. 185. ISBN 9788498350357. Consultado el 17 de julio de 2011.
- Manuel Malmierca, David Ryugo (2011). «cap:9 Descending Connections of Auditory Cortex to the Midbrainand Brain Stem». En Winer J.A., Schreiner C.E., ed. The Auditory Cortex. Researchgate.
- Zatorre, Robert J. (2002). «Auditory Cortex». Encyclopedia of Human Brain. pp. 289-301.
- Gage N.M., Baars B.J. (2018). «Cap.5 Sound, Speech, and Music Perception». Fundamentals of Cognitive Neuroscience (2 edición). pp. 143-184. Consultado el 14 de setiembre de 2019.
- Moerel M; De Martino F; Formisano E (2014). «An anatomical and functional topography of human auditory cortical areas». Front. Neurosci. (Revisión).
- Oren Poliva (2017). «From where to what: a neuroanatomically based evolutionary model of the emergence of speech in humans». F1000Research. Consultado el 8 de noviembre de 2018.
- Wallace M, Cronin MJ, Bowtell RW, Scott IS, Palmer AR, Gowland PA (2016). «Histological Basis of Laminar MRI Patterns in High Resolution Images of Fixed Human Auditory Cortex.». Front. Neurosci. Consultado el 8 de noviembre de 2018.
- Jara N, Délano PH (2014). «Avances en corteza auditiva». Rev. Otorrinolaringol. Cir. Cabeza Cuello (Revisión) 74 (3). Consultado el 8 de noviembre de 2018.
- Budinger E, Heil P (2006). «cap7 Anatomy of the auditory cortex». En Greenberg S, Ainsworth W, Ainsworth WA, ed. Listening to Speech: An Auditory Perspective. Psychology Press. pp. 91-93.
- Rudolf Nieuwenhuys; Cees A. J. Broere (2017). «A map of the human neocortex showing the estimated overall myelin content of the individual architectonic areas based on the studies of Adolf Hopf». Brain Struct Funct. 222 (1): 465-480. PMC 5225164. PMID 27138385. doi:10.1007/s00429-016-1228-7.
- Cant, N.B.; Benson, C.G. (15 de junio de 2003). «Parallel auditory pathways: projection patterns of the different neuronal populations in the dorsal and ventral cochlear nuclei». Brain Res Bull (en inglés) 60 (5–6): 457-74. PMID 12787867. doi:10.1016/S0361-9230(03)00050-9.
- Simon Baumann, Olivier Joly, Adrian Rees, Christopher I Petkov, Li Sun, Alexander Thiele, Timothy D Griffiths (2015). «The topography of frequency and time representation in primate auditory cortices». eLife 4: e03256. doi:10.7554/eLife.03256.
- W. Kalat, James (2004). «Percepción tonal en la corteza cerebral». Psicología biológica (8ª edición). Paraninfo. p. 170. ISBN 8497322851. Consultado el 17 de julio de 2011.
- Beament, James (2001). How We Hear Music: the Relationship Between Music and the Hearing Mechanism. Woodbridge: Boydell Press. p. 93. ISBN 9780851159409.
- Deutsch, Diana (Febrero). «Hearing Music in Ensembles». Physics Today (en inglés). p. 40. (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- Bendor, D.; Wang, X. (2005). «The neuronal representation of pitch in primate auditory cortex». Nature (en inglés) 436 (7054): 1161-5. PMC 1780171. PMID 16121182. doi:10.1038/nature03867. Archivado desde el original el 8 de marzo de 2010.
- Zatorre, R.J. (2005). «Neuroscience: finding the missing fundamental». Nature (en inglés) 436 (7054): 1093-4. PMID 16121160. doi:10.1038/4361093a. Archivado desde el original el 28 de noviembre de 2010.
- Dinh, L.; Nguyen T.; Salgado H.; Atzori M. (2009). «Norepinephrine homogeneously inhibits alpha-amino-3-hydroxyl-5-methyl-4-isoxazole-propionate- (AMPAR-) mediated currents in all layers of the temporal cortex of the rat». Neurochem Res (en inglés) 34 (11): 1896-906. PMID 19357950. doi:10.1007/s11064-009-9966-z.
- Lauter, Judith L.; P. Herscovitch; C. Formby; M.E. Raichle (1985). «Tonotopic organization in human auditory cortex revealed by positron emission tomography». Hearing Research (en inglés) (Elsevier B.V.) 20 (3): 199-205. PMID 3878839.
- Abbott, Alison (12 de marzo de 2002). «Neurobiology: Music, maestro, please!». Nature (en inglés) 416 (6876): 12-14. doi:10.1038/416012a. Archivado desde el original el 13 de octubre de 2008.
- Klinke, Rainer; Kral, Andrej; Heid, Silvia; Tillein, Jochen; Hartmann, Rainer (10 de septiembre de 1999). «Recruitment of the auditory cortex in congenitally deaf cats by long-term cochlear electrostimulation». Science (en inglés) 285 (5434): 1729-33. PMID 10481008. doi:10.1126/science.285.5434.1729.
- Strickland, Susan (2001). «Music and the brain in childhood development». Childhood Education (en inglés) 78 (2): 100-4.
- Bertrand, O.; Tallon-Baudry, C.; Fischer, C.; Pernier, J. Object representation and gamma oscillations (en inglés). Archivado desde el original el 19 de mayo de 2006.
- Knief, A.; Schulte, M.; Fujiki, N.; Pantev, C. Oscillatory Gamma band and Slow brain Activity Evoked by Real and Imaginary Musical Stimuli (en inglés).
- Petr Janata et al (2002). «The Cortical Topography of Tonal Structures Underlying Western Music». Science (en inglés) 298 (5601): 2167-2170. PMID 12481131. doi:10.1126/science.1076262.
- Cassel (septiembre de 1986). «Topography of projections from the medial prefrontal cortex to the amygdala in the rat». Brain Research Bulletin (en inglés) 17 (3): 321-33. PMID 2429740. doi:10.1016/0361-9230(86)90237-6.
- de Villers-Sidani, Etienne; E.F. Chang; S. Bao; M.M. Merzenich (2007). «Critical period window for spectral tuning defined in the primary auditory cortex (A1) in the rat». Journal of Neuroscience (en inglés) 27 (1): 180-9. PMID 17202485. doi:10.1523/JNEUROSCI.3227-06.2007.
- Guevara, E.; Alarcón, V. (2017). «Las Agnosias Auditivas: Una revisión teórica». Rev. Chil. Neuropsicol. (redalyc) 12 (1): 29-33. doi:10.5839/rcnp.2017.12.01.08.
- Alom Poveda J; Peña-Casanova J (2007). «cap. 2 Neuroanatomía conductual y Síndromes focales cerebrales; esquemas básicos». En Peña-Casanova J, ed. Panamericana. Neurología de la conducta y neuropsicología.
 Datos: Q18676
Datos: Q18676 Multimedia: Auditory cortex / Q18676
Multimedia: Auditory cortex / Q18676