Ecuaciones de Lotka-Volterra
Las ecuaciones de Lotka-Volterra, también conocidas como ecuaciones predador-presa o presa-predador, son un par de ecuaciones diferenciales de primer orden no lineales que se usan para describir dinámicas de sistemas biológicos en el que dos especies interactúan, una como presa y otra como depredador. Las ecuaciones fueron propuestas de forma independiente por Alfred J. Lotka en 1925 y Vito Volterra en 1926. Tales ecuaciones se definen como:



donde:
- y es el número de algún predador (por ejemplo, un lobo);
- x es el número de sus presas (por ejemplo, conejos);
- dy/dt y dx/dt representa el crecimiento de las dos poblaciones en el tiempo;
- t representa el tiempo; y
- α, β, γ y δ son parámetros (positivos) que representan las interacciones de las dos especies.
Historia
El modelo depredador-presa de Lotka-Volterra fue propuesto inicialmente por Alfred J. Lotka en la teoría de las reacciones químicas autocatalíticas en 1910.[1][1] Esta fue efectivamente la ecuación logística,[2] derivada originalmente por Pierre François Verhulst.[3] En 1920, Lotka extendió el modelo, a través de Andrey Kolmogorov, a "sistemas orgánicos" usando una especie de planta y una especie de animal herbívoro como ejemplo [4] y en 1925 usó las ecuaciones para analizar las interacciones depredador-presa en su libro sobre biomatemáticas.[5] El mismo conjunto de ecuaciones fue publicado en 1926 por Vito Volterra, un matemático y físico que se había interesado por la biología matemática.[6][10][7] La investigación de Volterra se inspiró en sus interacciones con el biólogo marino Umberto D'Ancona, quien estaba cortejando a su hija en ese momento y luego se convertiría en su yerno. D'Ancona estudió las capturas de peces en el mar Adriático y notó que el porcentaje de peces depredadores capturados había aumentado durante los años de la Primera Guerra Mundial (1914-18). Esto lo desconcertó, ya que el esfuerzo de pesca se había reducido mucho durante los años de guerra. Volterra desarrolló su modelo independientemente de Lotka y lo usó para explicar la observación de d'Ancona.[8]
Un ejemplo simple
.svg.png.webp)
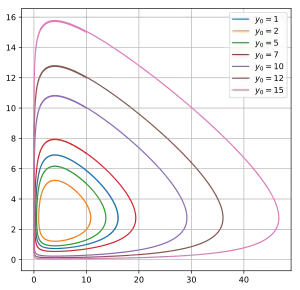
Supongamos que hay dos especies de animales, un babuino (presa) y un guepardo (depredador). Si las condiciones iniciales son 10 babuinos y 10 guepardos, se puede trazar la progresión de las dos especies a lo largo del tiempo; dados los parámetros de que las tasas de crecimiento y muerte de los babuinos son 1,1 y 0,4 mientras que las de los guepardos son 0,1 y 0,4 respectivamente. La elección del intervalo de tiempo es arbitraria.
También se pueden trazar soluciones paramétricamente como órbitas en el espacio de fase, sin representar el tiempo, pero con un eje que representa el número de presas y el otro eje que representa el número de depredadores para todos los tiempos.
Esto corresponde a eliminar el tiempo de las dos ecuaciones diferenciales anteriores para producir una sola ecuación diferencial

relacionar las variables x y y . Las soluciones de esta ecuación son curvas cerradas. Es susceptible de separación de variables: integrando

produce la relación implícita

donde V es una cantidad constante que depende de las condiciones iniciales y se conserva en cada curva.
Dinámica del sistema
En el sistema modelo, los depredadores prosperan cuando hay presas abundantes, pero, en última instancia, superan su suministro de alimentos y disminuyen. Como la población de depredadores es baja, la población de presas aumentará nuevamente. Estas dinámicas continúan en un ciclo poblacional de crecimiento y declive.
Equilibrio de la población
El equilibrio de la población se produce en el modelo cuando ninguno de los niveles de población cambia, es decir, cuando ambas derivadas son iguales a 0:


El sistema de ecuaciones anteriores nos da dos soluciones:

&

Por lo tanto, hay dos equilibrios en las ecuaciónes.
La primera solución representa efectivamente la extinción de ambas especies. Si ambas poblaciones están en 0, entonces seguirán estándolo indefinidamente. La segunda solución representa un punto fijo en el que ambas poblaciones mantienen sus números actuales distintos de 0 y, en el modelo simplificado, lo hacen indefinidamente. Los niveles de población en los que se logra este equilibrio dependen de los valores elegidos de los parámetros α, β, γ, & δ.
Estabilidad de los puntos fijos
La estabilidad del punto fijo en el origen se puede determinar realizando un linealización usando derivadas parciales.
La Matriz Jacobiana del modelo depredador-presa es

y esto se conoce como matriz comunitaria.
Primer punto fijo (extinción)
Cuando se evalúa en el estado estacionario de (0, 0), la matriz jacobiana J se trnansforma en

Los eigenvalores de la matriz son:

En el modelo α y γ son siempre mayores que cero y, como tal, el signo de los valores propios anteriores siempre será diferente. Por lo tanto, el punto fijo en el origen es un [Punto de silla|punto silla].
La inestabilidad de este punto fijo es importante. Si fuera estable, las poblaciones distintas de cero podrían verse atraídas hacia él y, como tal, la dinámica del sistema podría conducir a la extinción de ambas especies para muchos casos de los niveles de población iniciales. Sin embargo, como el punto fijo en el origen es un punto silla y, por lo tanto, inestable, se deduce que la extinción de ambas especies es difícil en el modelo. (De hecho, esto solo podría ocurrir si la presa fuera completamente erradicada artificialmente, causando que los depredadores mueran de hambre. Si los depredadores fueran erradicados, la población de presas crecería sin límites en este modelo simple). Las poblaciones de presas y depredadores pueden acercarse infinitesimalmente a cero y aun así recuperarse.
Segundo punto fijo (oscilaciones)
Evaluando J en el segundo punto fijo conduce a

Los eigenvalores de esta matriz son

Como los valores propios son puramente imaginarios y conjugados entre sí, este punto fijo debe ser un centro para órbitas cerradas en la vecindad local o una espiral atractiva o repulsiva. En los sistemas conservativos, debe haber órbitas cerradas en la vecindad local de puntos fijos que existen en los mínimos y máximos de la cantidad conservada. La cantidad conservada se deriva arriba para ser en las orbitas. Así, las órbitas alrededor del punto fijo son cerradas y elípticas, entonces las soluciones son periódicas, oscilando en una pequeña elipse alrededor del punto fijo, con una frecuencia & un periodo .



Como se ilustra en las oscilaciones circulantes de la figura anterior, las curvas de nivel están cerradas en órbitas que rodea el punto fijo: los niveles de las poblaciones de depredadores y presas ciclan y oscilan sin amortiguamiento alrededor del punto fijo con frecuencia .

El valor de la constante de movimiento V, o su equivlanete, K = exp(−V), , se puede encontrar para las órbitas cerradas cerca del punto fijo.

El incremento de K mueve una órbita cerrada más cerca del punto fijo. El mayor valor de la constante. K se obtiene resolviendo el problema de optimización:

El máximo valor de K se alcanza así en el punto estacionario (fijo) e incrementa a


donde e es el número de Euler.
Explicación de las ecuaciones
Usando las series de Taylor se obtiene una solución lineal a las ecuaciones:

.

Con estos coeficientes se puede estudiar los modelos de competición, enfermedad y mutualismo (biología) en un ecosistema.
Presa Lotka

Se asume que las presas tienen suministro de comida ilimitado por tiempo definido, y se reproducen exponencialmente a menos que exista algún predador. Este crecimiento exponencial está representado en la ecuación por el término αx. El término de la ecuación βxy viene a representar el encuentro de las dos especies y su interacción. Si x o y son cero no existe interacción.
Se puede interpretar la ecuación como el cambio del número de presas viene dado por su propio crecimiento menos la tasa de encuentros con predadores.
Depredador

En esta ecuación, δxy representa el crecimiento de los depredadores (fíjese en la similitud con la ecuación para las presas, pero en este caso para el crecimiento de los depredadores es necesario usar la razón a la que se consumen las presas, x). γy representa la muerte natural de los depredadores de forma exponencial; a más depredadores es necesario que el número de víctimas o presa aumente para mantener la población.
Se puede interpretar la ecuación como el crecimiento de los depredadores por la caza de presas menos la muerte natural de estos.
Referencias
- Lotka, Alfred J. (1 de marzo de 1910). «Contribution to the Theory of Periodic Reactions». The Journal of Physical Chemistry (en inglés) 14 (3): 271-274. ISSN 0092-7325. doi:10.1021/j150111a004. Consultado el 24 de marzo de 2023.
- Berryman, Alan A. (1992-10). «The Orgins and Evolution of Predator-Prey Theory». Ecology (en inglés) 73 (5): 1530-1535. doi:10.2307/1940005. Consultado el 24 de marzo de 2023.
- Garnier, Jean-Guillaume; Quetelet, Adolphe (1838). Correspondance mathématique et physique (en francés). M.Hayez, imprimeur. Consultado el 24 de marzo de 2023.
- Lotka, A. J. (1920-07). «Analytical Note on Certain Rhythmic Relations in Organic Systems». Proceedings of the National Academy of Sciences of the United States of America 6 (7): 410-415. ISSN 0027-8424. PMC 1084562. PMID 16576509. doi:10.1073/pnas.6.7.410. Consultado el 24 de marzo de 2023.
- J., Lotka, A. (1925). Elements of physical biology.. Williams & Wilkins. OCLC 709362232. Consultado el 24 de marzo de 2023.
- Maitra, S. C.; Montroll, E. W. (1971). On the Volterra and other nonlinear models of interacting populations. Academic Press. ISBN 0-12-287450-1. OCLC 204495. Consultado el 24 de marzo de 2023.
- Volterra, V. (1 de abril de 1928). «Variations and Fluctuations of the Number of Individuals in Animal Species living together». ICES Journal of Marine Science 3 (1): 3-51. ISSN 1054-3139. doi:10.1093/icesjms/3.1.3. Consultado el 24 de marzo de 2023.
- Kingsland, Sharon E. (1995). Modeling nature : episodes in the history of population ecology (2nd ed edición). University of Chicago Press. ISBN 0-226-43728-0. OCLC 32347988. Consultado el 24 de marzo de 2023.
Bibliografía
- Nicolas Bacaër, Rafael Bravo de la Parra, Jordi Ripoll. Breve historia de los modelos matemáticos en dinámica de poblaciones. Paris. ISBN 9791034365883. Pdf
| Control de autoridades |
|
|---|
 Datos: Q150052
Datos: Q150052 Multimedia: Lotka–Volterra equations / Q150052
Multimedia: Lotka–Volterra equations / Q150052