Eolambia caroljonesa
Eolambia caroljonesa es la única especie conocida del género extinto Eolambia (lat. “primer lambeosaurio”) de dinosaurio ornitópodo iguanodontiano, que vivió a mediados del período Cretácico, hace aproximadamente 110 y 95 millones de años, en el Albiense y Cenomaniense, en lo que es hoy Norteamérica.
| Eolambia caroljonesa | ||
|---|---|---|
| Rango temporal: 110 Ma - 95 Ma Cretácico inferior | ||
 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Ornithischia | |
| Suborden: | Neornithischia | |
| Infraorden: | Ornithopoda | |
| (sin rango): | Hadrosauriformes | |
| Superfamilia: | Hadrosauroidea | |
| Género: |
Eolambia Kirkland, 1998 | |
| Especie: |
E. caroljonesa Kirkland, 1998 | |
| Sinonimia | ||
| ||
Descripción
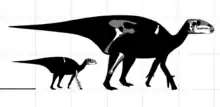
Eolambia es un gran hadrosauroide. Las estimaciones iniciales ubicaron la longitud de su cráneo en 1 metro de longitud,[1] aunque esto se debió a un hocico desproporcionadamente largo que luego se corrigió al descubrir material adicional.[2] En 2016, Gregory S. Paul estimó una longitud corporal de 6 metro y un peso de 1 tonelada para Eolambia,[3] que concuerda con una estimación previa de longitud corporal de 6,1 metros por Thomas R. Holtz Jr. en 2012.[4] Anteriormente, en 2008, se estimó que un espécimen adulto tenía una longitud de 5,2 metros y una altura en la cadera de 2 metros.[5]
Cráneo
El cráneo sin cresta de Eolambia tiene una forma general similar a la de Equijubus y Probactrosaurus . La parte frontal del hocico es muy áspera, y está marcada por muchos agujeros . En la punta de cada premaxilar, hay dos estructuras similares a dientes conocidas como dentículos, que también se ven en sus Protohadros su pariente más cercano. Más atrás, la parte posterior de la rama inferior del premaxilar se proyecta abruptamente hacia arriba, cerrando la fosa nasal en la parte posterior como en Probactrosaurus , Protohadros y otros hadrosauroides. Esta parte se une con los dos procesos similares a los dedos del maxilar , que es similar a Protohadros . El cuerpo del maxilar en sí no tiene un receso o cualquier indicación de una fenestra antorbital , como Equijubus , Protohadros y otros hadrosauroides. Una de las características utilizadas para distinguir Eolambia es el perfil cóncavo de la fila de dientes del maxilar cuando se ve desde el lado, que es como Equijubus , Probactrosaurus y varios otros hadrosauriformes, pero a diferencia de los Protohadros.[2]

Al igual que Probactrosaurus y otros hadrosauroides, la parte posterior del maxilar se conecta con el yugal, que bordea la parte inferior de la cuenca del ojo y la fenestra infratemporal , a través de una proyección similar a un dedo que cabe en un receso. El margen inferior del yugal tiene una brida fuerte debajo del nivel de la fenestra infratemporal, esto también se observa en Equijubus , Probactrosaurus , Protohadros y varios otros hadrosauroides. La conexión al yugal desde arriba es el posorbital , que tiene una superficie rugosa donde bordea las cuencas de los ojos, como en Prothadros, pero el lado del hueso es por lo demás liso. En la parte posterior del cráneo, el cuadrado se articula con el escamoso con una articulación que tiene forma de D cuando se ve desde la parte superior. Los escamosos de la izquierda y la derecha se habrían contactado extensamente, quedando separados por la parte posterior por un pequeño proceso parietal . El hueso supraoccipital , que forma la porción superior de la parte posterior del cráneo, es plano y casi vertical, como es el caso en Probactrosaurus y otros hadrosauroides.[2]
Al igual que con el premaxilar, el predentario de Eolambia llevaba dentículos. Hay un proceso dorsomedial prominente, una estructura similar a una pestaña que también se ve en Probactrosaurus y otros hadrosauriformes. Varios dentículos adicionales en forma de pestañas estaban presentes a ambos lados del proceso dorsomedial, que también están presentes en Probactrosaurus . El dentario , que constituye la mayor parte de la mandíbula inferior, se une al predentario en la parte posterior . Hay un receso corto, o diastema, entre la articulación del predentario con el dentario y la posición del primer diente en el dentario, que se observa en Equijubus, Probactrosaurus., y otros hadrosauroides. El frente del dentario se profundiza de manera característica, como en Protohadros, Ouranosaurus y Bactrosaurus . Dos protuberancias están presentes en la superficie externa del dentario, una de ellas representa el proceso coronoideo como en Probactrosaurus y otros hadrosauroides. Al igual que Protohadros y varios otros hadrosauriformes, pero a diferencia de Probactrosaurus, solo la parte delantera de este proceso está engrosada. Al igual que en Equijubus , Probactrosaurus y varios otros iguanodontidos , un foramen pequeño está presente en el lado del surangular., que se encuentra detrás del dentario.[2]

Al igual que en otros iguanodontidos, los dientes de Eolambia están dispuestos en filas estrechamente espaciadas y entrelazadas. En un momento dado, cada una de las 32 cavidades dentales superiores tiene tres dientes, mientras que cada una de las 30 cavidades dentarias tiene cuatro dientes. De estos, dos de los dientes en cada cavidad son dientes de reemplazo como los de Probactrosaurus, los otros son dientes activos. El diente muy activo tiene una faceta de desgaste . Al igual que Probactrosaurus, Gongpoquansaurus, Protohadros y otros hadrosauroides, cada corona del diente maxilar tiene solo una cresta, que está ligeramente desplazada hacia la línea media de la boca. Mientras tanto, cada corona dentaria.característicamente tiene una cresta primaria y una cresta accesoria más cerca de la línea media de la boca, una condición que también está presente en Protohadros y otros hadrosauroides. Todas las coronas también llevan pequeñas arrugas, similares a un dentículo, en los bordes delanteros y traseros, lo que también se ve en Probactrosaurus.[2]
Columna vertebral
Como Equijubus, Probactrosaurus, Gongpoquansaurus y otros styracosternanos, Eolambia tiene vértebras cervicales que son muy opistocoelicas, lo que significa que sus extremos frontales son fuertemente convexos mientras que sus extremos traseros son fuertemente cóncavos. Inusualmente, el extremo frontal de la tercera cervical se fija ligeramente más alto que el extremo posterior, que no se ve en ninguna otra vértebra cervical. Las vértebras de la parte posterior del cuello tienen un ángulo mayor entre los procesos articulares alargados conocidos como poszapófisis y también procesos transversales más alargados. En los procesos transversales, existen otros procesos articulares que se localizan en las parapofisis redondeadas y las diapofisis en forma de varilla. En los cervicales más posteriores, también hay una depresión profunda que separa los postzygapofisis, y las espinas neurales tienen márgenes frontales angulosos, lo que tiene el efecto de crear prominentes proyecciones en forma de espigas.[2][6]

Las primeras vértebras dorsales son similares a las vértebras cervicales, pero tienen espinas neurales más altas y con más puntas. El resto son anfiplatos, lo que significa que son planos en ambos extremos. También tienen postzygapofisis que son menos alargadas y más parecidas a pedestales, además de espinas neurales más altas y más rectangulares. Las parapofisis, que son depresiones en lugar de proyecciones, se han movido de los procesos transversales al arco neural , entre los procesos transversos y los procesos articulares conocidos como prezygapofisis en el frente de las vértebras. Sin embargo, vuelven a la base de los procesos transversales en los últimos dorsales.[2] En la tercera o cuarta dorsal, la parapófisis se localiza muy cerca de la sutura.entre la espina neural y el centro , que es diferente a los otros dorsales pero similar a los primeros dorsales en Edmontosaurus. También como Edmontosaurus , la arista mediana que separa los prezigapófisis se hace más pronunciada en la parte posterior dorsal.[6]

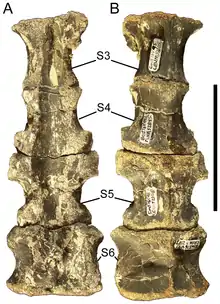
Entre los especímenes de Eolambia encontrados hasta la fecha, el sacro mejor conservado incluye siete vértebras. Dado que este individuo es inmaduro, y los hadrosaurios aumentan su número de vértebras sacras con la edad, los adultos pueden haber tenido más vértebras sacras.[1] Las vértebras caudales son anfócelas, teniendo los extremos delantero y trasero siendo cóncavos. Tienen prezigapofisis parecidas a pedestales y postzygapofisis similares a pestañas, estas últimas separadas por una depresión. Los prezygapofisis se alargan hasta convertirse en tallos y los postzygapofisis se encogen en los caudales medio y trasero. Además, las espinas neurales pasan de ser rectangulares a fuertemente curvadas, cóncavas en el frente y convexas en la parte posterior, los centros se vuelven más alargados y los procesos transversales también desaparecen.[2][6]
Cinturas y miembros
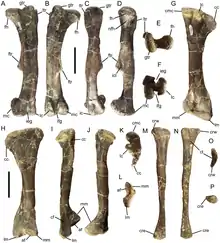
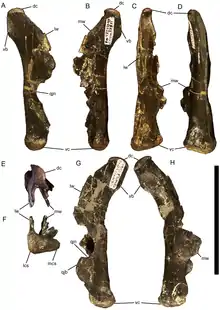
Al igual que Probactrosaurus , la lámina de la escapula de Eolambia es casi rectangular, con los lados de las lámina rectos y el extremo de la hoja en un ángulo casi recto. El húmero está inclinado hacia la línea media, y la cresta deltopectoral en el húmero está prominentemente engrosado. Hay un botón en la superficie exterior de la articulación con el radio en el húmero. Tanto el cúbito como el radio son huesos relativamente rectos. La parte superior del cúbito tiene tres "puntas", a saber, el proceso de olécranon y dos bridas triangulares adicionales. Mientras tanto, el radio tiene una superficie interna aplanada en el extremo superior y una plataforma distinta en la superficie interna en el extremo inferior, ambas para articular con el cúbito. La mano tiene cinco dígitos. Entre los huesos metacarpianos de la mano, el segundo es curvo, el tercero es recto y el cuarto es fuertemente curvado. Al igual que Iguanodon, Probactrosaurus y otros iguanodontianos basales, la garra en el primer dígito es distintivamente cónica. La segunda y tercera garras son similares a las pezuñas, con la tercera garra con bordes rugosos en sus lados, un rasgo que también se comparte con otros iguanodontes.[2]

El proceso preacetabular, es decir, ubicado frente a la cavidad de la cadera, del ilion lleva una "bota" horizontal que forma un ángulo obtuso con la parte inferior del proceso. Además, el borde superior de los proyectos de hueso hacia el exterior para formar un borde situado encima de la articulación ilion-isquion en el borde inferior del hueso. Estos rasgos también se ven en Probactrosaurus y otros iguanodontianos.[2] La cavidad de la cadera en sí es bastante grande en relación con el ilion para un iguanodontiano.[7] Un borde distintivo está presente a lo largo de la superficie superior del ilion, un rasgo que se comparte con un hadrosauroide sin nombre de la Formación Woodbine. El proceso postacetabular, es decir, ubicado detrás del zócalo de la cadera, del ilion se estrecha en su parte posterior sin interrupción en su margen superior. En el pubis , el proceso de proyección hacia adelante se expande hacia el frente, como Probactrosaurus , Gongpoquansaurus y otros iguanodontianos. El eje del isquion es característicamente recto en los adultos, los juveniles tienen un eje curvo, que es más común entre los iguanodontes, y la parte inferior del eje tiene una "bota" comprimida que se expande hacia adelante.[2][6]

Al igual que el húmero, el fémur está inclinado, pero hacia afuera. Sin embargo, característicamente, la mitad inferior del fémur es recta. El cuarto trocánter , ubicado a la mitad del fémur, es verticalmente alto y triangular, similar a Probactrosaurus , Gongpoquansaurus y otros iguanodontianos, pero diferente a los ornitópodos más basales. Al igual que el cúbito y el radio, la tibia y el peroné tienen ejes rectos y se articulan entre sí, una proyección rugosa en la parte inferior del peroné se ajusta entre dos cóndilos en forma de bulbo ubicados justo detrás de la cresta cnemial, en el extremo inferior de la tibia. La parte superior de la tibia es áspera, lo que indica la presencia de una tapa de cartílago entre el fémur y la tibia. El pie tiene cuatro dígitos. Al igual que los metacarpianos, el segundo metatarsiano es curvo, el tercero es recto y el cuarto es fuertemente curvado. Las falanges del pie son muy fuertes, y las falanges terminales cerca del final de cada dígito son particularmente fuertes. Al igual que con la segunda y tercera garras en la mano, la segunda, tercera y cuarta garras del pie son uniformemente anchas, planas y con forma de pezuña.[2]
Descubrimiento e investigación
En 1979, Peter Galton y James A. Jensen describieron un fémur fragmentario perteneciente a un hadrosauroide descubierto en el parque nacional Arches en Utah. También se encontraron varios dientes de hadrosauroides en las canteras de pequeños vertebrados en la región occidental de San Rafael Swell, cerca de Castle Dale en el condado de Emery, Utah. Fueron descritos en 1991 por J. Michael Parrish. Posteriormente, en 1993, Carole Jones y su esposo Ramal Jones descubrieron huesos fragmentarios en un sitio fósil ubicado en la región noroeste de Swell. Trajeron el sitio a la atención de Donald Burge, director de la institución que luego llamó al Museo Prehistórico del Colegio del Este de Utah (CEUM). El sitio, que se conoce formalmente como CEUM Locality 42em366v, posteriormente se llamaría el Sitio de Carol, en su honor. Los fósiles, almacenados bajo el número de muestra CEUM 9758, representan los restos parciales de un hadrosauroide adulto, que incluye partes del cráneo, vértebras, isquion y pata. CEUM 5212, una calavera parcial y miembro anterior de un adulto, se encontró cerca en la localidad de CEUM 42em369v.[1] CEUM 8786, un fémur izquierdo de un adulto, se descubrió más tarde en el sitio de Carol y no se describió hasta 2012.[2]

Al mismo tiempo, el Museo de Historia Natural de Oklahoma había recuperado restos pertenecientes al mismo hadrosauroide de las excavaciones en la región suroeste de Swell.[1] Las muestras descubiertas por el museo de Oklahoma inicialmente consistían en seis individuos de cinco localidades, dos esqueletos juveniles,[7] incluyendo vértebras, escápulas, cúbito, ilion e isquion, y elementos de extremidades posteriores de la localidad OMNH v237, un esqueleto juvenil parcial, que incluye partes del cráneo, extremidades anteriores y miembros posteriores junto con una vértebra dorsal de OMNH v824, OMNH 27749, un sacro e isquion de OMNH v696, OMNH 24389, un isquion de OMNH v214 y OMNH 32812, un esqueleto parcial que incluye una escápula, dos vértebras caudales y otros elementos no excavados de OMNH v866. El Museo de Historia Natural de Oklahoma también hizo descubrimientos adicionales de los dientes del hadrosauroide en varias canteras. Richard Cifelli, del Museo de Historia Natural de Oklahoma, le permitió a James Kirkland estudiar las muestras recolectadas.[1] Kirkland continuó describiendo preliminarmente los especímenes en 1994 en un resumen de la conferencia en la reunión anual de la Sociedad de Paleontología de Vertebrados.[8]
En un artículo de 1998 publicado en el boletín del Museo de Historia Natural y Ciencia de Nuevo México, Kirkland nombró formalmente a los especímenes como un nuevo género , Eolambia, con la única especie y tipo, Eolambia caroljonesa. El nombre genérico combina el prefijo griego ἠώς, eos, "amanecer", "mañana", que implica "temprano" con el sufijo lambia derivado de Lambeosaurus , que a su vez lleva el nombre del paleontólogo canadiense Lawrence Lambe. En total, el nombre del género significa "lambeosaurino temprano", en referencia a su supuesta posición como el lambeosaurino más basal. Mientras tanto, el nombre específico honra a Carole Jones. El paleoartista Michael Skrepnick sugirió el nombre Eolambia ; reemplazó el nombre informal "Eohadrosaurus caroljonesi", que Kirkland usó antes de Documento de 1998.[1]

Excavaciones adicionales realizadas desde 1998 han revelado elementos esqueléticos adicionales que pertenecen a Eolambia . Ahora es el iguanodóntido más completamente conocido entre los estadios de Cenomaniano y Santoniano. Casi todos los elementos del esqueleto que pertenecen al taxón están representados por múltiples fósiles en las colecciones existentes. Al menos doce individuos, incluidos al menos ocho juveniles,[5] fueron descubiertos en una cantera al sur de la ciudad de Emery , incluidas partes del cráneo y del resto del cuerpo. La cantera es conocida como la Cifelli#2 Cantera de o más formalmente CEUM localidad 42em432v. Otra cantera al sur de la ciudad de Emery, la cantera Willow Springs 8 o la localidad CEE 42Em576v, conserva al menos cuatro juveniles. El material de estas canteras fue descrito en 2012 por Kirkland junto con Andrew McDonald, John Bird y Peter Dodson, quienes también proporcionaron un diagnóstico actualizado para Eolambia basado en este material.[2]

Un espécimen descubierto en la cantera Cifelli#2, un dentario derecho catalogado como CEUM 34447, es inusual en comparación con otros especímenes de Eolambia, incluidos adultos y juveniles. Mientras que la relación entre la profundidad punto medio y la longitud total de los dentarios en otros individuos generalmente oscila desde 0,19 hasta 0,24, el mismo valor en CEUM 34447 es 0,31, lo que es inusualmente profunda. El dentario tampoco se profundiza sustancialmente en la parte delantera, y por lo tanto carece de uno de los rasgos distintivos de los dentarios de Eolambia . Sin embargo, el otro rasgo distintivo, que es la expansión del extremo frontal del proceso coronoideo, está presente en la muestra. Aunque es posible que este individuo represente un espécimen inusual de Eolambia., McDonald y sus colegas advirtieron que puede representar un hadrosauroideo distinto que difiere en la morfología del dentario de Eolambia aunque sea algo que no se sugiere por cualquiera de los otros materiales en la cantera. Por lo tanto, lo consideraron como un hadrosauroide basal indeterminado.[2]
El Museo Field de Historia Natural también realizó excavaciones al sur de la ciudad de Emery de 2009 a 2010, con permisos del Servicio Geológico de Utah. Estas excavaciones se realizaron en un sitio descubierto por Akiko Shinya en 2008, en la localidad de FMNH UT080821-1, que ha sido nombrada el sitio de Akiko en su honor. Durante las excavaciones, se recolectaron 167 elementos desarticulados pero estrechamente asociados en dos bochones de yeso. Una de estos bochones de yeso, que contenía elementos de la cadera y las vértebras, recibió el número de muestra FMNH PR 3847. McDonald describió posteriormente el espécimen junto con Terry Gates, Lindsay Zanno y Peter Makovicky en 2017. Otra localidad descubierta por el Museo Field es la cantera Triple Peak, localidad UTN30304-2 del FMNH, que contiene un hueso de más de 400 huesos juveniles que pertenecen a por lo menos cuatro individuos.[6]
Datación del sitio
Todas las localidades en las que se descubrió Eolambia pertenecen al Miembro Mussentuchit de la formación Cedar Mountain. Las rocas predominantes en Mussentuchit consisten en capas entrelazadas de gris claro a gris de arenisca fangosa y areniscas. Los depósitos que representan dos sistemas de canales fluviales separados también están presentes en el Mussentuchit. El más bajo de estos sistemas consta de cuatro capas, respectivamente, de arenisca fangosa, arenisca de grano fino, arenisca limosa y arenisca de grano fino, y las dos últimas capas contienen restos de plantas. La cantera Cifelli#2 está asociada con este sistema. El sistema superior consiste en arenisca de grano fino verdosa. Tres capas de las capas de arcilla de bentonita, que representan depósitos de cenizas volcánicas , también están presentes en el Mussentuchit, que son respectivamente de color gris blanquecino, amarillo y tostado. La mitad de estas capas atraviesa la cantera Cifelli#2.[5] La bentonita también está presente dentro de la piedra arenisca en sí en todo el Mussentuchit.[6] En la cantera Cifelli#2, los huesos están ennegrecidos y han sido reemplazados por calcita , cuarzo , pirita y carbonato - hidroxiapatita.[5]
La palinología, el estudio del polen,[9][10][11] así como los estudios de megaflora e invertebrados fósiles,[12] habían sugerido inicialmente que la formación Cedar Mountain superior, incluida la Mussentuchit, databa del Albiano.[1] Una datación argón-argón de 1997 de cristales de sanidina recuperados de una capa de ceniza volcánica en la formación de Cedar Mountain superior por Cifelli y sus colegas indicó una edad de 97 ± 0.1 millones de años, que luego se consideró justo después del límite entre Etapas albiana y cenomaniana. Esto es similar a la temprana edad de los habitantes del cenomaniano reportada para la superpuesta formación Dakota.[13] Posteriormente, en 2007, James Garrison y sus colegas salieron de la capa de ceniza volcánica que pasa a través de la cantera Cifelli#2, que probablemente sea la misma capa que antes Cifelli y colegas, hace 96.7 ± 0.5 millones de años. A un nivel aproximado, las estimaciones de Cifelli y Garrison concuerdan entre sí, lo que indica una edad de Cenomaniana para Eolambia. Garrison y sus colegas también volvieron a analizar el ensamblaje de polen de las localidades de todo Mussentuchit, concluyendo que el ensamblaje abarca la frontera albiana-cenomaniana.[5]
Clasificación
Primera clasificación como Lambeosaurinae
En la descripción inicial de Kirkland de Eolambia, lo consideraba un miembro de los Hadrosauridae, tal como lo definieron David B. Weishampel , David B. Norman y Dan Grigorescu en 1993. Weishampel y sus colegas utilizaron siete características unificadoras para definir el Hadrosauridae. La expansión ascendente del proceso ascendente del maxilar, la ausencia del foramen paracuadrado, que separa el cuadrados y quadratojugal, la ubicación del angular en la superficie interna de la mandíbula inferior, la ausencia del foramen surangular en el surangular, los dientes estrechos del maxilar, la presencia de tres o más dientes en cada posición del diente dentario y la reducción del margen superior de la lámina escapular.[14] El primero, quinto, sexto y séptimo de estos rasgos fueron reconocidos en .Eolambia, y el resto se desconoce debido a la falta de material. Kirkland asignó además a Eolambia a la Euhadrosauria, definida por Weishampel y sus colegas para incluir al ancestro común de Hadrosaurinae, ahora Saurolophinae y Lambeosaurinae, las dos ramas principales de los hadrosauridos, y todos sus descendientes.[14] Esta asignación se basó en cinco características, todas presentes en Eolambia la presencia de dentículos en la premaxila, la estrecha articulación del cuadrado con la mandíbula inferior, los dientes dentarios estrechos, la presencia de un solo borde, o carina, en cada diente dentario y la cresta deltopectoral en ángulo sobre el húmero.[1]

Dentro de los Hadrosauridae, Kirkland consideró además que .Eolambia era un miembro basal de los Lambeosaurinae, o el grupo hermano de Lambeosaurinae. Identificó cinco características compartidas con las Lambeosaurinas, según se definió en varios estudios, la ausencia de un foramen en la premaxila, el encierro, al menos parcial, de la fosa nasal por parte de la premaxila, el desarrollo de una estantería sobre el maxilar, las espinas neurales muy altas de las vértebras caudales, la robustez del húmero; y la gran "bota" del isquion en adultos.[14][15][16][17] Kirkland descubrió que dos rasgos adicionales separaban la cresta de Eolambia de otros lambeosaurines con cresta, que se consideraron relacionados con el desarrollo de la cresta, la elevación de la cavidad nasal sobre la cavidad del ojo y el acortamiento del parietal. Si Eolambia fuera reconocido como una lambeosaurino, entonces la Lambeosaurinae tendría que ser redefinida para excluir esos dos rasgos. Alternativamente, si Eolambia fuera el grupo hermano de las Lambeosaurinas, entonces representaría una morfología en los hadrosauridos que está cerca de la divergencia entre las hadrosaurinos y las lambeosaurinos. Esta posibilidad está respaldada por la presencia de un surco en el fondo del sacro de Eolambia, que fue reconocido por Kirkland como un rasgo definitorio de los hadrosaurinos,[16][17] aunque presente con convergencia en la anquilasauria y la ceratopsia.[1]
Para apoyar las afinidades de Lambeosaurinae de Eolambia, Kirkland realizó un análisis filogenético, el árbol recuperado por su análisis, que en consecuencia coloca a Eolambia como el Lambeosaurinae más basal, se reproduce a continuación. A pesar de la evidencia que citó en apoyo de esta posición, Kirkland también notó un rasgo aparentemente inconsistente. Como se observó en el ilion juvenil que describió inicialmente, el antitrocanter, una proyección en el ilion que limita al trocánter mayor del fémur, está muy poco desarrollado de lo que se esperaría para una Hadrosauridae basal. Michael Brett-Surman notó que los adultos pueden haber tenido un antitrocántico mejor desarrollado. Por otro lado, los derivados Lambeosaurinae como Hypacrosaurus tiene un antitrocánter bien desarrollado incluso en la fase de cría. Kirkland notó la posibilidad de que los hadrosauridos basales hayan desarrollado su antitrocanter más tarde en la vida que los hadrosauridos derivados, como Hypacrosaurus.[18] Por lo tanto, observó que los especímenes adultos de Eolambia resolverían este problema.[1]
Filogenia como Lambeosaurinae
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Reidentificación como no Hadrosauridae
En 2001, Jason Head revaluó la posición filogenética de Eolambia basándose en muestras almacenadas en el Museo de Oklahoma. Reconoció que la identificación de Kirkland de los supuestos rasgos distintivos de Lambeosaurinae en Eolambia era errónea o dudosa. Como lo demostró Head, en realidad hay un foramen en la premaxila, la premaxila solo limita la fosa nasal en el borde inferior, hay dos procesos similares a los dedos en la parte frontal del maxilar, además de la plataforma, Las espinas neurales altas de las vértebras caudales también aparecen en los no lambeosaurinos como Ouranosaurus y Bactrosaurus y la expansión de la punta del isquion también ocurre en Camptosaurus, Iguanodon, Ouranosaurus, Gilmoreosaurus y Bactrosaurus , lo que indica que la expansión es plesiomórfica para los iguanodontos y se perdió secundariamente en los Hadrosauridae.[17] Mientras Casanovas y sus colegas en 1999 caracterizan lambeosaurinos como teniendo un "pie ampliado" en la punta del isquion, en oposición a una condición de "tipo garrote",[19] Head argumento que Corythosaurus y Lambeosaurus[20] tienen Las puntas isquiales que no son más extensas que las de Mantellisaurus u Ouranosaurus.[7][21]
Head también demostró que los dientes de Eolambia eran más anchos y asimétricos de lo que Kirkland había asumido, también señaló que los subadultos solo poseían dos dientes por posición de diente, a diferencia de los hadrosáuridos,[7] aunque las muestras adultas de hecho tienen las tres o más posiciones de diente como lo demostró Kirkland originalmente.[2] La cabeza demostró además que Eolambia poseía siete vértebras sacras, a diferencia de ocho o más en los hadrosauridos derivados, y que su antitrocanter estaba en realidad menos desarrollado que el de Ouranosaurus y Altirhinus.[22] El examen de los especímenes del Museo de Oklahoma demostró además que la región postacetabular del ilion no formó un proceso distinto, y las articulaciones en la parte inferior del fémur no son muy prominentes, y ambas separan a Eolambia de iguanodontos más derivados, como los hadrosáuridos. Por lo tanto, Head concluyó que Eolambia era un miembro basal de la Hadrosauroidea, fuera de la Hadrosauridae, una conclusión apoyada por un análisis filogenético que descubrió que era un hadrosauroide no hadrosáurido estrechamente relacionado con Probactrosaurus.[7]
En 2004, Norman incluyó a Eolambia por primera vez en un análisis filogenético completo de iguanodontos, como parte de un capítulo que se centra en los iguanodontos en la segunda edición de "The Dinosauria". Encontró que Eolambia era el grupo hermano de Altirhinus y que ambos eran más basales que Protohadros, Probactrosaurus o Hadrosauridae.[23] Dentro del mismo volumen, Horner, Weishampel y Catherine Forster realizaron un análisis filogenético separado de los hadrosauridos, que también incluye a Eolambia. Encontraron a Eolambia de nuevo como un hadrosauroide no hadrosaúrido, aunque como el grupo hermano de Protohadros. Aunque Head había redefinido específicamente las Hadrosauridae, basadas en características compartidas, para incluir Protohadros,[7][24] Horner y sus colegas adaptaron una definición basada en taxón que excluía a Protohadros y, por lo tanto, a Eolambia. También identificaron características adicionales que diferenciaban a Eolambia de los hadrosáuridos, existen dentículos gruesos en los dientes del dentario y el proceso coronoideo está débilmente expandido.[25]
La variación en las posiciones filogenéticas recuperadas para Eolambia persistió en los años siguientes. En la descripción de 2009 de Levnesovia , Hans-Dieter Sues y Alexander Averianov encontraron que -Protohadros ocupaba una posición intermedia en relación con Altirhinus y Probactrosaurus, siendo el grupo hermano de Fukuisaurus.[26] Mientras tanto, de acuerdo con los resultados de Horner y sus colegas, Albert Prieto-Márquez y Mark Norell encontraron en 2010 que Eolambia era en cambio el grupo hermano de Protohadros. También señaló características de Eolambia que eran convergentes sobre los hadrosáuridos, la presencia de una carina de un solo diente, la esquina exterior casi cuadrada del predentario y el punto medio de la muesca cuadratojugal, que se articula con el cuadratojugal, se ubica aproximadamente a la mitad de la superficie del cuadrante, se ubica a menos del 60% de la altura del hueso desde la parte superior.[27] En 2012, Holtz clasificó a ,,Eolambia,, como un miembro primitivo de Styracosterna junto con Altirhinus.[4]
En 2012, McDonald realizó un análisis filogenético de iguanodontos que incorporó datos de nuevas muestras de Eolambia. Contrariamente a los autores anteriores, descubrió que Eolambia era el grupo hermano de Probactrosaurus. Esta estrecha relación se basó en dos características compartidas, lel cuadrado es recto excepto para el extremo superior, que se curva hacia atrás y los márgenes superior e inferior de la lámina escapular son casi paralelos.[2] Wenjie Zheng y sus colegas agregaron Jintasaurus al grupo en 2014,[28] mientras que José Gasca y sus colegas recuperaron el grupo original en 2015.[29] Sin embargo, McDonald revisó exhaustivamente su conjunto de datos filogenéticos en preparación para la descripción de 2017 de los especímenes de Museo Field. Al igual que con Horner y sus colegas, así como con Prieto-Márquez y Norell, McDonald y sus colegas encontraron que Eolambia era el grupo hermano de Protohadros, y que ambos se colocaron como hadrosauromorfos basales como se ve a continuación.[6] Utilizando la versión 2012 del conjunto de datos de McDonald, Francisco Verdú y sus colegas recuperaron de forma independiente un resultado similar[30][31] y Eolambia está estrechamente relacionada con Protohadros y Jeyawati. Sin embargo, McDonald y sus colegas encontraron que este último se derivó más en 2017.[6]
Filogenia como no Hadrosauridae

| Hadrosauriformes |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Últimos Hallazgos
En 2018, Terry Gates y sus colegas describieron el nuevo género iguanodotiano, Choyrodon, de la misma localidad que Altirhinus, considerado en el pasado un pariente de Eolambia. Su análisis filogenético basado en la matriz de Norman de 2015 encontró que Choyrodon era el grupo hermano de Eolmabia. Protohadros el relativos comúnmente recuperados no estaban presentes en esta matriz. Tres características apoyaron esta relación, un occipucio de forma trapezoidal y dos rasgos compartidos de los dientes. La ubicación de estos géneros en relación con otros Hadrosauriformes dependía de si la fenestra antorbital estaba codificada como presente o ausente, en Choyrodon el espécimen tipo es un juvenil tiene una abierta, pero no se puede descartar la posibilidad de que los adultos tengan uno cerrado. Los siguientes dos árboles muestran los resultados con el código codificado como presente y ausente respectivamente, el primero muestra una politomía grande.[32]
Posición dentro de Hadrosauriformes
|
|
Paleobiología
Cáscaras de huevo posiblemente pertenecientes a Eolambia se han descubierto en varias localidades junto con dientes aislados. Tienen patrones de superficie reticular, similar a una red y tienen un grosor de 2 milímetros.[1]
Conocidos por una variedad de especímenes juveniles y adultos, los cambios que Eolambia experimentó a medida que crecieron están bien documentados. El número de dientes en el maxilar aumentó con la edad, de 23 en un juvenil a 33 en un adulto. Un aumento similar ocurrió en los dientes dentarios , de 18 a 22 en los juveniles a 25 a 30 en los adultos. Mientras que los juveniles carecen de crestas secundarias en las coronas de sus dientes dentarios, las crestas secundarias débiles están presentes en los adultos.[2] El tamaño de la "bota" del isquion también aumentó con la edad, según lo documentado por Kirkland.[1] También pueden haberse producido cambios en la rectitud del eje del isquion, según lo documentado por las muestras descubiertas por el Museo Field.[6]
McDonald y sus colegas informaron los resultados de un análisis histológico realizado en una costilla del espécimen FMNH PR 3847 en 2017. Se ha producido una remodelación ósea extensa en la matriz interna de la costilla. La cavidad medular, que alberga la médula, está marcada por la presencia de muchas cavidades o lagunas. En la superficie externa del hueso, los osteones han reemplazado casi completamente el interior del hueso. El crecimiento en capas del hueso es más visible en el interior del hueso, donde se encuentran cinco líneas de crecimiento detenido son visibles. Una sexta línea de crecimiento retenida está posiblemente presente dentro de la región medular, pero tiene un alcance relativamente limitado, lo que hace que esta identificación sea cuestionable.[6]

Dado que las líneas de crecimiento retenida pueden ser eliminadas por la remodelación ósea, esto representa una subestimación del número real de las líneas de crecimiento retenida que realmente se han desarrollado. McDonald y sus colegas estimaron tres o cuatro líneas de crecimiento retenida perdidas al examinar el espacio entre las líneas de crecimiento retenidas visibles. Esto indica que el animal tenía entre ocho y nueve años cuando murió. El espaciado entre las líneas de crecimiento retenidas disminuye significativamente hacia la superficie exterior, lo que indica que el crecimiento se estaba desacelerando, pero la falta del sistema fundamental externo, una serie de líneas de crecimiento retenidas estrechamente espaciadas indica que el crecimiento aún no ha cesado. Esto es consistente con las suturas no fusionadas entre las espinas centrales y neurales en las vértebras cervicales y dorsales, y las suturas no fusionadas entre los centros sacros y sus costillas correspondientes.[33] En general, la tasa de crecimiento de Eolambia:Parece haber sido comparable al de Hypacrosaurus y Maiasaura, que alcanzaron la madurez después de siete u ocho años.[6][34][35]
Paleoecología
Medio ambiente y tafonomía
Los datos sedimentológicos del miembro de Mussentuchit indican que Eolambia vivía en una llanura de inundación mal drenada, alrededor de un sistema de grandes lagos perennes. Los lagos fluctuaron entre los niveles de agua altos y bajos en los ciclos de Milankovitch de aproximadamente 21,000 años, causados por la precesión axial de la Tierra. Estos ciclos se refleja por la alternancia de capas de arenisca fangosa y lutolita en depósitos de roca de hoy en día. Durante los períodos de sequía, el lecho del lago expuesto formó una amplia playa desprovisto de plantas. El barro agrietado del lakebed produjo clastos de barro, mientras que los desechos orgánicos y los cadáveres barridos se esparcían por la orilla del agua a medida que el lago se retiraba progresivamente. Mientras tanto, durante los períodos en que aumentó el nivel del mar, pequeños ríos invadieron la llanura de inundación, que volvió a trabajar y desplazó los escombros que se habían depositado durante la estación seca.[5]
La mayoría de los fósiles descubiertos en Mussentuchit están dispersos y desarticulados, y los esqueletos intactos son relativamente raros. Aunque los lagos no destruyeron activamente los huesos, tampoco fue particularmente propicio para una buena conservación debido a la larga duración del proceso de enterramiento. Antes del entierro, los carroñeros podrían haber alterado el material del esqueleto, lo que es evidente en las cortadas de una tibia de Eolambia que muestran las marcas de dientes descubiertas en la cantera Cifelli#2. La regresión del lago podría haber creado ocasionalmente pantanos , pero no habrían sido lo suficientemente permanentes como para atrapar y preservar vertebrados más grandes. La acción de las corrientes de agua parece haber jugado un papel en la deposición de huesos de Eolambia en la cantera Cifelli#2, donde los huesos se depositan en gran parte a los 28°, 69°, 93°, 131° y 161° acimut, es decir, en relación con el norte. Estas direcciones habrían sido paralelas o sub-paralelas a los bordes del lago, indicando la orientación de las corrientes lacustres, a excepción de los acimutes de 69 ° y 93 °, que probablemente representan las corrientes de los ríos.[5]
La abundancia de restos de plantas, combinada con la ausencia de depósitos de sumideros o playa, indica que el clima local era relativamente húmedo durante la deposición de Mussentuchit. Esto está en marcado contraste con los depósitos anteriores en el área, que son calcáreos, lo que indica períodos de aridez que duran hasta 10000 años. Las reconstrucciones paleoclimáticas apoyan estas interpretaciones, con la invasión del Mar de Mowry desde el norte, un proceso que eventualmente formaría la vía marítima interior occidental, que resulta en climas áridos, interior de latitud media continental, siendo reemplazados por más húmedos, costa este de latitud media.[36] Con una paleolatitud de 46 ° N, el Mussentuchit habría tenido una temperatura superficial media cálida de 40–45 °C, aproximadamente 6–8 °C más cálida que la actual, según modelos de circulación general.[5][37]
Ecosistema
Los crocodilomorfos son abundantes en los depósitos de Mussentuchit, representados en gran parte por dientes. Los dientes más grandes conocidos se han referido a la Pholidosauridae,[38][6] son anchos con bases redondeadas, y algunas de ellas tienen crestas longitudinales.[6] Estos grandes cocodrilos hubieran sido capaces de atacar al menos Eolambia juveniles al borde del agua. La abundancia de ejemplares juveniles de Eolambia en el material fósil de la orilla del lago apoya esta hipótesis. Eolambia más grandes habría sido invulnerable a los cocodrilos.[5] Los dientes de cocodrilo más pequeños también están presentes, con dos morfotipos distintos: una forma ovoide que tiene crestas, que se ha referido a dos especies diferentes de Bernissartia.[5] y una forma triangular comprimida que lleva una pesada ornamentación, que se ha atribuido a Atoposauridae.[13][39] Sin embargo, dado el tamaño pequeño y uniforme de estos dientes y la co-ocurrencia de estos morfotipos en algunos cocodrilos,[38] pueden pertenecer a la misma especie.[6]
Los Eolambia adultos habría sido vulnerable[5] a los grandes neovenatoridos como Siats,[40] que fue descubierto por las expediciones de la Museo Field.[6] Una variedad de terópodos manirptoranos más pequeños también están presentes en el Mussentuchit, habría funcionado como carroñeros, sus restos incluyen dientes atribuidos a Dromaeosauridae, Paronychodon y Richardoestesia.[39] En cuanto a los dinosaurios herbívoros, el nodosáuridos Animantarx fue descubierto en el sitio de Carol, cerca del espécimen tipo de Eolambia.[41] También se descubrieron dientes pertenecientes a más ornitópodos basales y en 1999 se refirieron dientes similares a la Pachycephalosauria o a aves dentadas.[5][39] Restos más completos se refieren a dos ornitópodos diferentes, un orodrominído y un iguanodonte basal que permanece inédito.[42] También se ha descubiertoun oviraptorosauriano gigante,[43] junto con dientes de ceratopsianos y saurópodos.[6][39]
Los peces, que pueden haber sido cazados por pequeños cocodrilos, están representados por dientes y escamas. Incluyen un raya Myliobatiformes del género Baibisha,[39] una miembro indeterminado de Neopterygii, un semionotiformes referida al género Lepidotes, un pejelagarto relacionado con Atractosteus,[6] un pycnodontidae se refirió al género Stephanodus y un amiiformes indeterminado. Salamandras indeterminadas y mamíferos también están presentes. En cuanto a las plantas, las muestras de polen indican un bosque escalonado que rodea los lagos de Mussentuchit, que contiene un dosel de helechos arborescentes, araucarias y coníferas, que acompañan una maleza de helechos, gimnospermas pequeñas y plantas con flores. Las algas también habrían estado presentes en las aguas de los lagos.[10][44] Estas plantas habrían proporcionado una fuente abundante de alimento para Eolambia.[5]
Paleobiogeografía
Kirkland, Cifelli y sus colegas notaron que la fauna de Mussentuchit compuesta de iguanodontos, paquicefalosaurianos y ceratopsianos tienen fuertes similitudes con las faunas asiáticas contemporáneas. Propusieron que Eolambia era parte de una afluencia de dinosaurios asiáticos a América del Norte durante el Cenomaniano, que suplantó a la fauna nativa de baja diversidad anterior.[1][13] Esta hipótesis está respaldada por la estrecha relación entre Eolambia y Probactrosaurus o Fukuisaurus, que se han recuperado respectivamente por las filogenias de Head y Sues & Averianov.[2][7][26]
Sin embargo, los resultados de Horner y sus colegas, Prieto-Márquez, y McDonald y sus colegas, que consideran que el norteamericano Protohadros es el pariente más cercano de Eolambia, contradicen esta hipótesis.[6][25][27][45] Esta interpretación alternativa sugiere que el ancestro común de Eolambia, Protohadros y más hadrosauroides derivados no fue exclusivamente asiático, sino que se distribuyó en Asia y América del Norte.[2] McDonald y sus colegas sugirieron además que Eolambia y Protohadros representan un grupo localizado de hadrosauroides endémicos de América del Norte, al igual que otros grupos de styracosternanos en todo el Cretácico. Tales grupos contemporáneos incluían un grupo de Asia central de Shuangmiaosaurus, Zhanghenglong y Plesiohadros.[6]
Referencias
- Kirkland, J.I. (1998). «A new hadrosaurid from the upper Cedar Mountain Formation (Albian-Cenomanian: Cretaceous) of eastern Utah – the oldest known hadrosaurid (lambeosaurine?)». En Lucas, S.G.; Kirkland, J.I.; Estep, J.W., eds. Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History and Science Bulletin 14. pp. 283-295.
- McDonald, A.T.; Bird, J.; Kirkland, J.I.; Dodson, P. (2012). «Osteology of the Basal Hadrosauroid Eolambia caroljonesa (Dinosauria: Ornithopoda) from the Cedar Mountain Formation of Utah». PLoS ONE 7 (10): e45712. Bibcode:2012PLoSO...745712M. PMC 3471925. PMID 23077495. doi:10.1371/journal.pone.0045712.
- Paul, G.S. (2010). «Ornithischians». The Princeton Field Guide to Dinosaurs. Princeton: Princeton University Press. p. 327. ISBN 9780691167664.
- Holtz, T.R. (2012). «Genus List for Holtz (2007) Dinosaurs». Supplementary Information to Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages by Thomas R. Holtz, Jr., illustrations by Luis Rey. Department of Geology, University of Maryland. Consultado el 30 de agosto de 2017.
- Garrison, J.R. Jr.; Brinkman, D.; Nichols, D.J.; Layer, P.; Burge, D.; Thayn, D. (2007). «A multidisciplinary study of the Lower Cretaceous Cedar Mountain Formation, Mussentuchit Wash, Utah: a determination of the paleoenvironment and paleoecology of the Eolambia caroljonesa dinosaur quarry». Cretaceous Research 28 (3): 461-494. doi:10.1016/j.cretres.2006.07.007.
- McDonald, A.T.; Gates, T.A.; Zanno, L.E.; Makovicky, P.J. (2017). «Anatomy, taphonomy, and phylogenetic implications of a new specimen of Eolambia caroljonesa (Dinosauria: Ornithopoda) from the Cedar Mountain Formation, Utah, USA». PLoS ONE 12 (5): e0176896. Bibcode:2017PLoSO..1276896M. doi:10.1371/journal.pone.0176896.
- Head, J.J. (2001). «A reanalysis of the phylogenetic position of Eolambia caroljonesa (Dinosauria, Iguanodontia)». Journal of Vertebrate Paleontology 21 (2): 392-396. doi:10.1671/0272-4634(2001)021[0392:AROTPP]2.0.CO;2.
- Kirkland, J.I.; Burge, D. (1994). «A large primitive hadrosaur from the Lower Cretaceous of Utah». Fifty-Fourth Annual Meeting, Society of Vertebrate PaleontologyJournal of Vertebrate Paleontology 14 (3): 32A. JSTOR i406457.
- Craig, L.C. (1981). «Lower Cretaceous rocks, southwestern Colorado and southeastern Utah». Rocky Mountain Association of Geologists 1981 Field Conference: 195-200.
- Tschudy, R.H.; Tschudy, B.D.; Craig, L.C. (1984), Palynological evaluation of Cedar Mountain and Burro Canyon formations, Colorado Plateau, US Geological Survey, Professional Paper 1821, US Geological Survey, pp. 1-24.
- Nichols, D.J.; Sweet, A.R. (1993). «Biostratigraphy of Upper Cretaceous nonmarine palynofloras in a north-south transect of the Western Interior Basin». En Caldwell, W.G.E.; Kauffman, E.G., eds. Evolution of the Western Interior Basin. Geological Association of Canada, Special Paper 39. Geological Association of Canada. pp. 539-584.
- Young, R.G. (1960). «Dakota Group of Colorado Plateau». American Association of Petroleum Geologists Bulletin 44 (2): 156-194. doi:10.1306/0bda5fd2-16bd-11d7-8645000102c1865d.
- Cifelli, R.L.; Kirkland, J.I.; Weil, A.; Deino, A.L.; Kowallis, B.J. (1997). «High-precision 40Ar/39Ar geochronology and the advent of North America’s Late Cretaceous terrestrial fauna». Proceedings of the National Academy of Sciences 94 (21): 11163-11167. Bibcode:1997PNAS...9411163C. PMC 23402. doi:10.1073/pnas.94.21.11163.
- Weishampel, D.B.; Norman, D.B.; Grigorescu, D. (1993). «Telmatosaurus transsylvanicus from the Late Cretaceous of Romania: the most basal hadrosaurid dinosaur». Palaeontology 36 (2): 361-385.
- Sereno, P.C. (1986). «Phylogeny of the bird-hipped dinosaurs (Order Ornithischia)». National Geographic Research 2: 234-256.
- Brett-Surman, M.K. (1989). A revision of the Hadrosauridae (Reptilia: Ornithischia) and their evolution during the Campanian and Maastrichtian (Ph.D.). Washington D.C.: George Washington University. pp. 1-373.
- Weishampel, D.B.; Horner, J.R. (1990). «Hadrosauridae». En Weishampel, D.B.; Dodson, P.; Osmolska, H., eds. The Dinosauria (1st edición). Berkley: University of California Press. pp. 534-561.
- Horner, J.R.; Currie, P.J. (1994). «Embryonic and neonatal morphology and ontogeny of a new species of Hypacrosaurus (Ornithischia, Lambeosauridae) from Montana and Alberta». En Carpenter, K.; Hirsch, K.F.; Horner, J.R., eds. Dinosaur Eggs and Babies. Cambridge: Cambridge University Press. pp. 312-336. ISBN 9780521567237.
- Casanovas, M.L.; Pereda Suberbiola, X.; Santafe, J.V.; Weishampel, D.B. (1999). «First lambeosaurine hadrosaurid from Europe: palaeobiogeographical implications». Geological Magazine 136 (2): 205-211. Bibcode:1999GeoM..136..205C. doi:10.1017/s0016756899002319.
- Lull, R.S.; Wright, N.E. (1942). «Hadrosaurian Dinosaurs of North America». Geological Society of America Special Papers 40: 1-242. doi:10.1130/SPE40-p1.
- Norman, D.B. (1986). «On the anatomy of Iguanodon atherfieldensis (Ornithischia: Ornithopoda)». Bulletin de L'Institut Royal Des Sciences Naturelles de Belgique 56: 281-372.
- Norman, D.B. (1998). «On Asian ornithopods (Dinosauria: Ornithischia). 3. A new species of iguanodontid dinosaur». Zoological Journal of the Linnean Society 122 (1): 291-348. doi:10.1111/j.1096-3642.1998.tb02533.x.
- Norman, D.B. (2004). «Basal Iguanodontia». En Weishampel, D.B.; Dodson, P.; Osmólska, H., eds. The Dinosauria (2nd edición). Berkeley: University of California Press. pp. 413-437.
- Head, J.J. (1998). «A new species of basal hadrosaurid (Dinosauria, Ornithischia) from the Cenomanian of Texas». Journal of Vertebrate Paleontology 18 (4): 718-738. doi:10.1080/02724634.1998.10011101.
- Horner, J.R.; Weishampel, D.B.; Forster, C.A. (2004). «Basal Iguanodontia». En Weishampel, D.B.; Dodson, P.; Osmólska, H., eds. The Dinosauria (2nd edición). Berkeley: University of California Press. pp. 438-463.
- Sues, H.-D.; Averianov, A. (2009). «A new basal hadrosauroid dinosaur from the Late Cretaceous of Uzbekistan and the early radiation of duck-billed dinosaurs». Proceedings of the Royal Society B 276 (1667): 2549-2555. PMC 2686654. doi:10.1098/rspb.2009.0229.
- Prieto-Márquez, A.; Norell, M.A. (2010). «Anatomy and Relationships of Gilmoreosaurus mongoliensis (Dinosauria: Hadrosauroidea) from the Late Cretaceous of Central Asia». American Museum Novitates 3694: 1-49. doi:10.1206/3694.2.
- Zheng, W.; Jin, X.; Shibata, M.; Azuma, Y. (2014). «An early juvenile specimen of Bolong yixianensis (Ornithopoda: Iguanodontia) from the Lower Cretaceous of Ningcheng County, Nei Mongol, China». Historical Biology 26 (2): 236-251. doi:10.1080/08912963.2013.809347.
- Gasca, J.M.; Moreno-Azanza, M.; Ruiz-Omeñaca, J.I.; Canudo, J.I. (2015). «New material and phylogenetic position of the basal iguanodont dinosaur Delapparentia turolensis from the Barremian (Early Cretaceous) of Spain». Journal of Iberian Geology 41 (1): 57-70. doi:10.5209/rev_jige.2015.v41.n1.48655.
- Verdú, F.J.; Royo-Torres, R.; Cobos, A.; Alcalá, L. (2015). «Perinates of a new species of Iguanodon (Ornithischia: Ornithopoda) from the lower Barremian of Galve (Teruel, Spain)». Cretaceous Research 56: 250-264. doi:10.1016/j.cretres.2015.05.010.
- Verdú, F.J.; Royo-Torres, R.; Cobos, A.; Alcalá, L. (2017). «New systematic and phylogenetic data about the early Barremian Iguanodon galvensis (Ornithopoda: Iguanodontoidea) from Spain». Historical Biology 30 (4): 1-38. doi:10.1080/08912963.2017.1287179.
- Terry A. Gates; Khishigjav Tsogtbaatar; Lindsay E. Zanno; Tsogtbaatar Chinzorig; Mahito Watabe (2018). «A new iguanodontian (Dinosauria: Ornithopoda) from the Early Cretaceous of Mongolia». PeerJ 6: e5300. doi:10.7717/peerj.5300.
- Brochu, C.A. (1994). «Closure of neurocentral sutures during crocodilian ontogeny: Implications for maturity assessment in fossil archosaurs». Journal of Vertebrate Paleontology 16 (1): 49-62. doi:10.1080/02724634.1996.10011283.
- Horner, J.R.; de Ricqlès, A.; Padian, K. (1999). «Variation in dinosaur skeletochronology indicators: implications for age assessment and physiology». Paleobiology 25 (3): 295-304. doi:10.1017/S0094837300021308.
- Woodward, H.N.; Freedman Fowler, E.A.; Farlow, J.O.; Horner, J.R. (2015). «Maiasaura, a model organism for extinct vertebrate population biology: a large sample statistical assessment of growth dynamics and survivorship». Paleobiology 41 (4): 503-527. doi:10.1017/pab.2015.19.
- Elder, W.P.; Kirkland, J.I. «Cretaceous Paleogeography of the Southern Western Interior Region». En Caputo, M.V.; Peterson, J.A.; Franczyk, K.J., eds. Mesozoic Systems of the Rocky Mountain Region, USA. Denver: Rocky Mountain Section SEPM. pp. 415-440.
- Glancy, T.J. Jr.; Arthur, M.A.; Barron, E.J.; Kaufman, E.G. (1993). «A paleoclimate model for the North American Cretaceous (Cenomanian-Turonian) epicontinental sea». En Caldwell, W.G.E.; Kauffman, E.G., eds. Evolution of the Western Interior Basin. Geological Association of Canada, Special Papers 39. Geological Association of Canada. pp. 219-241.
- Irmis, R.B.; Hutchison, J.H.; Sertich, J.J.W.; Titus, A.L. (2013). «Crocodyliforms from the Late Cretaceous of Grand Staircase-Escalante National Monument and Vicinity, Southern Utah, USA». En Titus, A.L.; Loewen, M.A., eds. At the Top of the Grand Staircase: the Late Cretaceous of Southern Utah. Bloomington: Indiana University Press. pp. 424-444. ISBN 9780253008961.
- Cifelli, R.L.; Nydam, R.L.; Gardner, J.D.; Weil, A.; Eaton, J.G.; Kirkland, J.I.; Madsen, S.K. (1999). «Medial Cretaceous vertebrates from the Cedar Mountain Formation, Emery County, Utah: the Mussentuchit local fauna». En Gillette, D., ed. Vertebrate Paleontology in Utah. Utah Geological Survey Miscellaneous Publications. 99–1. pp. 219-242.
- Zanno, L.E.; Makovicky, P.J. (2013). «Neovenatorid theropods are apex predators in the Late Cretaceous of North America». Nature Communications 4: 2827. Bibcode:2013NatCo...4E2827Z. PMID 24264527. doi:10.1038/ncomms3827.
- Carpenter, K.; Kirkland, J.I.; Burge, D.L.; Bird, J. (1999). «Ankylosaurs (Dinosauria: Ornithischia) of the Cedar Mountain Formation, Utah, and their stratigraphic distribution». En Gillette, D., ed. Vertebrate Paleontology in Utah. Utah Geological Survey Miscellaneous Publications. 99–1. pp. 243-251.
- Zanno, L.E.; Makovicky, P.J. (2016). A New Species of Early Diverging Ornithopod Increases the Paleobiodiversity of Herbivorous Dinosaurs in Late Cretaceous Ecosystems in North America. Society of Vertebrate Paleontology 76th Annual Meeting. Salt Lake City: Society of Vertebrate Paleontology. p. 256. Archivado desde el original el 18 de octubre de 2016. Consultado el 24 de octubre de 2018.
- Makovicky, P.J.; Zanno, L.E. (2014). Kirkland, J.I.; Hunt-Foster, R.; Liggett, G.A. et al., eds. New Discoveries Add to the Diversity of the Mussentuchit Member (Cedar Mountain Formation) Dinosaur Fauna. Mid-Mesozoic: The Age of Dinosaurs in Transition. Fruita: Utah Geological Survey. pp. 70-71.
- Ravn, R.L. (1995). «Miospores from the Muddy Sandstone (Upper Albian), Wind River Basin, Wyoming, USA». Palaeontographica Abteilung B 234 (3): 41-91.
- Prieto-Márquez, A. (2010). «Global historical biogeography of hadrosaurid dinosaurs». Zoological Journal of the Linnean Society 159 (2): 503-525. doi:10.1111/j.1096-3642.2010.00642.x.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikimedia Commons alberga una categoría multimedia sobre Eolambia caroljonesa.
Wikimedia Commons alberga una categoría multimedia sobre Eolambia caroljonesa. Wikispecies tiene un artículo sobre Eolambia caroljonesa.
Wikispecies tiene un artículo sobre Eolambia caroljonesa.- Ficha e imagen del eolambia, en Dinosaurier-Web (en alemán)
- Eolambia dentro de Iguanodontia en The Thescelsossaurus (en inglés)
- Significado y pronunciación del nombre (en inglés)
- Reclasificación del Eolambia (en inglés)