Edmontosaurus
Edmontosaurus ("lagarto de Edmonton") es un género extinto conocido de dinosaurio ornitópodo hadrosáurido. Contiene dos especies conocidas: Edmontosaurus regalis y Edmontosaurus annectens. Se han encontrado fósiles de E. regalis en rocas del oeste de América del Norte que datan de fines del Campaniense hace 73 millones de años, mientras que las de E. annectens se encontraron en la misma región geográfica, pero en rocas que datan de finales del Maastrichtiense, hace 66 millones de años. Edmontosaurus fue uno de los últimos dinosaurios no aviares, y vivió junto a dinosaurios como Triceratops, Tyrannosaurus y Pachycephalosaurus poco antes de la extinción masiva del Cretácico-Paleógeno.
| Edmontosaurus | ||
|---|---|---|
| Rango temporal: 73 Ma - 66 Ma Cretácico Superior | ||
 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Ornithischia | |
| Suborden: | Ornithopoda | |
| (sin rango): | Iguanodontia | |
| Superfamilia: | Hadrosauroidea | |
| Familia: | Hadrosauridae | |
| Subfamilia: | Saurolophinae | |
| Tribu: | Edmontosaurini | |
| Género: |
Edmontosaurus Lambe, 1917 | |
| Especie tipo | ||
|
Edmontosaurus regalis Lambe, 1917 | ||
| Otras especies | ||
| ||
| Sinonimia | ||
| ||
Edmontosaurus incluyó algunas de las especies de hadrosáuridos más grandes, midiendo hasta 12 metros de largo y pesando alrededor de 4 toneladas. Existe evidencia en forma de dos especímenes fosilizados alojados en el Museo de las Rocosas para un tamaño máximo aún mayor de 15 metros y un peso de 10 toneladas para Edmontosaurus annectens. Se conocen varios especímenes bien conservados que incluyen no solo huesos, sino en algunos casos impresiones extensas de piel y posible contenido intestinal. Se clasifica como un género de saurolofino (o hadrosaurino) hadrosáurido, un miembro del grupo de hadrosáuridos que carecía de crestas grandes y huecas, y en su lugar tenía crestas sólidas más pequeñas o peines carnosos.
Los primeros fósiles de Edmontosaurus fueron descubiertos al sureste de Alberta, Canadá, en la zona usualmente llamada "Edmonton inferior" que pertenece a la Formación Cañón Herradura. La especie tipo, E. regalis, fue nombrada por Lawrence Lambe en 1917, aunque varias otras especies que ahora están consideran en Edmontosaurus fueron nombradas antes. La más conocida de estas probablemente sea E. annectens, nombrada por Othniel Charles Marsh en 1892; como una especie de Claosaurus, conocida durante muchos años como una especie de Trachodon y más tarde como Anatosaurus annectens. Anatosaurus y Anatotitan ahora se consideran generalmente sinónimos de Edmontosaurus.
Edmontosaurus se distribuyó ampliamente en el oeste de América del Norte. La distribución de los fósiles de Edmontosaurus sugiere que prefirió las costas y las llanuras costeras . Era un herbívoro que podía moverse en dos y cuatro patas. Debido a que se sabe de varios lechos óseos , se cree que Edmontosaurus vivió en grupos y también pudo haber sido migratorio. La riqueza de los fósiles ha permitido a los investigadores estudiar su paleobiología en detalle, incluido su cerebro, cómo se pudo haber alimentado y sus lesiones y patologías , como la evidencia de ataques de tiranosaurios en algunos especímenes de edmontosaurios.
Descripción
Edmontosaurus se describe a partir de numerosos especímenes fósiles.[2][3][4][5] Al igual que otros hadrosaurios, las patas delanteras eran menos fuertes que las traseras, pero igualmente lo suficientemente largas como para ser usadas al estar en pie o en movimiento. Por lo que su posición bípeda o tetrápoda variaba con la velocidad a la que deseaba desplazarse o si estaba detenido. Poseía una larga cola aplanada lateralmente y en su gran cabeza tenía un amplio pico semiesférico similar al de un pato en su boca llena de pequeños dientes con los que trituraba su alimento.

Las proporciones de los Edmontosaurus varían considerablemente según la especie e individuo, presentando tamaños en adultos que van desde los 9 metros con especímenes de 13 metros,[6] hasta los 14 metros de largo.[7] Esto sumado a un peso estimado de aproximadamente 4 toneladas, por lo que exhibían una de las mayores dimensiones entre los hadrosáuridos.[8]
Tradicionalmente, E. regalis ha sido considerada la especie más grande, aunque esto fue desafiado por la hipótesis de que el hadrosáurido más grande, Anatotitan copei, es sinónimo de Edmontosaurus annectens, tal como lo plantearon Jack Horner et al.,[8] y apoyado en estudios de Campione y Evens en 2009 y 2011.[9] El espécimen tipo de E. regalis, NMC 2288, se estima entre 9 a 12 metros de largo.[5] E. annectens a menudo se considera más pequeño. Los dos esqueletos bien conocidos montados el USNM 2414 y el YPM 2182, miden 8 y 9 metros respectivamentes.[5][10] Sin embargo, estos son probablemente individuos subadultos.[9] Hay al menos un informe de un espécimen potencial de E. annectens mucho más grande, casi 12 metros de largo.[11] Dos especímenes aún en estudio en la colección del Museo de las Rocosas, una cola de 7,6 metros etiquetada como MOR 1142 y otra etiquetada como MOR 1609, indican que E. annectens podría haber crecido a tamaños más grandes, posiblemente rivalizando con Shantungosaurus en tamaño. Según los paleontólogos, las muestras de estos individuos indican una longitud de hasta 15 metros.[12] Tales individuos grandes de Edmontosaurus probablemente habrían sido muy raros debido a factores tales como el estrés ambiental, la enfermedad y la depredación. El holotipo de E. saskatchewanensis, hoy considerado un juvenil de E. annectens, es más pequeño aún, con un largo total estimado de 7 a 7,3 metros.[5][4]
Cráneo

El cráneo de un ejemplar completamente desarrollado de Edmontosaurus media alrededor de 1 metro de largo.[5] Un cráneo de E. annectens, anteriormente Anatotitan, mide 1,18 metros de largo.[13] El cráneo era triangular visto de perfil,[2] sin ninguna cresta ósea.[5] Visto desde arriba el frente y la parte trasera del cráneo estaban expandida, con la parte frontal formando una especie de pico de pato. El pico no poseía dientes, y tanto la parte superior y la inferior poseían un pico córneo de queratina,[8] de la cual buena parte de la valva superior se hallaron en la momia de Trachodon que se encuentra en el Museo Senckenberg.[6] En este espécimen, el pico preservado se proyecta hacia adelante y abajo por lo menos 8 centímetros más allá del hueso.[5] Las narinas eran elongadas y fueron contenidos en las depresiones profundas rodeadas por los distintos bordes óseos por arriba, detrás, y abajo.[14] En por lo menos un caso, el espécimen de Senckenberg, se preservaron los anillos escleróticos encontrados en la órbita ocularque raramente se preservan.[5] Otro hueso raramente visto, el estribo del oído, también se ha encontrado en un espécimen de Edmontosaurus.[8]
Los dientes solo se encuentran en las mejillas, sobre el hueso maxilar en la parte superior y en el dentario en la inferior. Los dientes se sustituyeron continuamente, llevando alrededor de medio año en formarse.[15] Estaban compuestos por seis tipos de tejidos, que rivalizaban con la complejidad de los dientes de mamíferos.[16] Estos dientes crecían en columnas, con hasta por lo menos seis dientes en una columna, y el número de columnas varia según el tamaño del animal.[5] Los conteos de columnas conocidas para las dos especies son, 51 a 53 columnas por maxilar y 48 a 49 por dentario, los dientes de la mandíbula superior son ligeramente más estrechos que los de la mandíbula inferior, para E. regalis y 52 columnas por maxilar y 44 por dentario para E. annectens, una muestra de E. saskatchewanensis.[5]
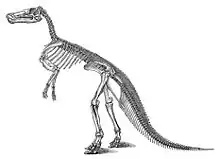
Esqueleto postcraneal

El número de vértebras difiere en número según la especie. E. regalis poseía 13 vértebras cervicales, 18 dorsales, 9 sacras, y un número desconocido de vértebras caudales.[5] El espécimen alguna vez identificado como Anatosaurus edmontoni, hoy considerado como E. annectens, es reportado con un vértebra dorsal adicional y 85 caudales, con una cantidad sin revelar en restauración .[5] Otros hadrosáuridos se informan teniendo solamente 50 a 70 vértebras en la cola,[8] por lo que parece haber sido una sobrestimación. La porción anterior de la espalda se curvaba hacia el suelo, con el cuello doblado hacia arriba y el resto de la espalda y de la cola sostenidas horizontalmente.[8] La mayor parte del lomo y la cola estaba reforzado por un enrejado de tendones osificados entre las espinas de las vértebras. Esta condición se ha descrito como parte de la espalda y por lo menos de la base de la cola rígida .[17][18] Estos tendones osificados son interpretados como un refuerzo de la columna vertebral para soportar el peso del cuerpo, soportado solo por las caderas y patas[17]
La escápula era un hueso plano con forma de espada, paralelo a la columna vertebral. La pelvis se componía por tres elementos a cada lado, un alargado ilion por encima de la articulación de la pata trasera, el isquion por debajo y por detrás como una varilla larga y delgada y el pubis en el frente que se dilataba en una estructura similar a una placa y perforado. La estructura de la cadera evitaba que el animal se colocara erguido, porque en tal posición el fémur no habría estado en contacto contra el sólido ilion, sino contra la articulación del ilion y del pubis. Las nueve vértebras sacras se fusionaron para ayudar a la estabilidad de la cadera.[5]
Las patas delanteras eran más cortas y estaban construidas más débilmente que las traseras. La parte superior del húmero tenía una gran cresta deltopectoral para la unión muscular, mientras que el cúbito y el radio eran delgados. La parte superior del brazo y el antebrazo eran similares en longitud. El carpo era simple, con solo dos huesos pequeños. Cada mano tenía cuatro dedos, sin el primer dedo. Los dedos segundo, tercero y cuarto tenían aproximadamente la misma longitud y estaban unidos en la vida dentro de una cubierta carnosa. Aunque el segundo y tercer dedo tenían unguinales parecidos a cascos, estos huesos también estaban dentro de la piel y no eran evidentes desde el exterior. El quinto dedo divergía de los otros tres y era mucho más corto. El hueso del muslo era robusto y recto, con un cuarto trocánter prominente aproximadamente a la mitad del lado posterior. Esta cresta era para la unión de poderosos músculos unidos a las caderas y la cola que tiraban de los muslos y por lo tanto de las patas traseras, hacia atrás y ayudaban a mantener el uso de la cola como órgano de equilibrio. Cada pie tenía solo tres dedos de los pies, sin dedo primero y el quinto. Los dedos tenían puntas parecidas a cascos.[5]
Piel
Múltiples ejemplares de Edmontosaurus annectens se han encontrado con impresiones de piel conservadas . Varios han sido bien publicitados, como la " momia de Trachodon " de principios del siglo XX,[19][20] y el espécimen apodado "Dakota",[21][22][23] este último aparentemente incluye compuestos orgánicos remanentes de la piel.[23] Debido a estos hallazgos, la escamación de Edmontosaurus annectens es conocida para la mayoría de las áreas del cuerpo. Las impresiones de la piel son menos conocidas para E. regalis, pero se han estudiado algunos ejemplos bien conservados, incluido uno que conserva una cresta de tejido blando o una barba en la cabeza. Se desconoce si dicha cresta estaba presente en E. annectens o y si era un indicador de dimorfismo sexual.[24]

En 1908 se descubrió la llamada "Momia de Trachodon" en Wyoming que es especialmente notable para la paleontología ya que se recuperaron impresiones fosilizadas de piel de Edmontosaurus. La impresión debe haber quedado por la deshidratación superficial muy rápida y la fijación su forma en el limo. Por estas impresiones sabemos que la piel era escamosa y coriácea, y el músculo del muslo estaba debajo de la piel del cuerpo. Esto habría dado la impresión que la pierna salía del cuerpo en la rodilla, y el muslo entero estaba debajo de la piel. Esto también contribuye a su semejanza a un pato. También tenía un número de nódulos a lo largo de su cuello, espalda y cola.[19][20]

Un espécimen juvenil de E. annectens fue recuperado con impresiones fosilizadas de la piel incluyendo los nódulos así como texturas previamente no vistas de la piel. Estas impresiones incluyeron escamas traslapadas elípticas, escamas acanaladas, y una estructura de 9 por 10 cm trapezoidal con forma de cuerno. Las impresiones fueron recuperadas de la cola del espécimen.[25]
Una rhamphotheca preservada presente en el espécimen de E. annectens LACM 23502, ubicado en el Museo de Historia Natural del Condado de Los Ángeles, indica que el pico de Edmontosaurus era más ganchudo y extenso de lo que muchas ilustraciones en medios científicos y públicos han descrito anteriormente. No se sabe si el espécimen en cuestión conservó la verdadera rhamphotheca o solo un molde de la estructura interna unida al hueso.[26][27][28]
Descubrimiento e investigación
Edmontosaurus ha tenido una historia larga y complicada dentro de la paleontología, después de haber pasado décadas con varias especies clasificadas en otros géneros. Su historia taxonómica se entrelaza en varios puntos con los géneros Agathaumas, Anatosaurus, Anatotitan, Claosaurus, Hadrosaurus, Thespesius y Trachodon[8][29] y las referencias anteriores a la década de 1980 típicamente usan Anatosaurus, Claosaurus, Thespesius o Trachodon para los fósiles de Edmontosaurus excluyendo los asignados a E. regalis, según el autor y la fecha. Aunque Edmontosaurus recién fue nombrado en 1917, su especie más antigua bien sustentada, E. annectens fue nombrada en 1892 como una especie de Claosaurus.
Siglo XIX-1917
La primera especie bien soportada de Edmontosaurus fue nombrada en 1892 como Claosaurus annectens por Othniel Charles Marsh. Esta especie se basa en USNM 2414, un techo de cráneo parcial y esqueleto, con un segundo cráneo y esqueleto, YPM 2182, designado como paratipo. Ambos fueron recolectados en 1891 por John Bell Hatcher de la formación Lance del Cretácico Superior en el Maastrichtiense tardío, del condado de Niobrara, entonces parte del condado de Converse, Wyoming, EE.UU..[30] Esta especie tiene algunas notas al pie de página de la historia de la paleontología. Es uno de los primeros dinosaurios en recibir una restauración esquelética, y es los primeros hadrosáuridos restaurados.[29][31] YPM 2182 y UNSM 2414 son, respectivamente, los primeros y segundos esqueletos de dinosaurios montados esencialmente completos en los Estados Unidos.[10] YPM 2182 se puso en exhibición en 1901[29] y USNM 2414 en 1904.[10]
Debido a la comprensión incompleta de los hadrosáuridos en el momento, después de la muerte de Marsh en 1897, Claosaurus annectens se clasificó de diversas maneras, como una especie de Claosaurus, Thespesius o Trachodon. Las opiniones varían mucho, los libros de texto y las enciclopedias establecían una distinción entre el Claosaurus annectens "tipo Iguanodon" y el Hadrosaurus "pico de pato", basado en restos ahora conocidos para un Edmontosaurus annectens adulto, mientras que Hatcher identificaba explícitamente C. annectens como sinónimo con el hadrosáurido representado por el mismo cráneo de pico de pato.[29] La revisión de Hatcher, publicada en 1902, fue radical, consideró casi todos los géneros de hadrosáuridos conocidos entonces como sinónimos de Trachodon. Esto incluía a Cionodon, Diclonius, Hadrosaurus, Ornithotarsus, Pteropelyx y Thespesius, así como a Claorhynchus y Polyonax, géneros fragmentarios que ahora se piensa que son dinosaurios con cuernos.[32] El trabajo de Hatcher condujo a un breve consenso, hasta que después de 1910 el nuevo material de Canadá y Montana mostró una mayor diversidad de hadrosáuridos de lo que se sospechaba anteriormente.[29] Charles W. Gilmore en 1915 revaluó hadrosáuridos y recomendó que Thespesius fuera reintroducido para los hadrosáuridos de la Formación Lance y las unidades rocosas de edad equivalente, y que Trachodon, basado en material inadecuado, debería restringirse a un hadrosáurido de la antigua Formación Judith River y sus equivalentes. Con respecto a Claosaurus annectens, recomendó que se considere lo mismo que Thespesius occidentalis.[33] Su restitución de Thespesius para los hadrosáuridos lanciano tendría otras consecuencias para la taxonomía de Edmontosaurus en las décadas siguientes.
Los primeros restos fósiles conocidos que pueden pertenecer a Edmontosaurus regalis fueron nombrados Trachodon cavatus en 1871 por Edward Drinker Cope.[34] El nombre se deletrea en fuentes más recientes como Trachodon avatus o incluso Trachodon atavus.[6][8][35] Esta especie fue evaluada sin comentarios como sinónimo de Edmontosaurus regalis en dos revisiones,[8][35] aunque el atavus es anterior a regalis por varias décadas. En 1874, Cope nombró, pero no describió, a Agathaumas milo por un sacro y fragmentos de vértebras y piernas de finales del Maastrichtiano en la formación Laramie de Colorado.[6][36][37] Más tarde, ese mismo año, describió estos huesos con el nombre de Hadrosaurus occidentalis.[6][37][38] Los huesos ahora están perdidos.[37] Al igual que con Trachodon atavus, Agathaumas milo ha sido asignado sin comentarios a Edmontosaurus regalis en dos revisiones,[8][35] aunque preceden a regalis por varias décadas. Ninguna de las especies ha atraído mucha atención, ambos están ausentes de la monografía de 1942 de Lull y Wright, por ejemplo. Una tercera especie poco conocida, Trachodon selwyni, descrita por Lawrence Lambe en 1902 para una mandíbula inferior de lo que ahora se conoce como la Formación del Parque de los Dinosaurios de Alberta, [19] fue erróneamente descrita por Glut en 1997 como asignada a Edmontosaurus regalis.[39] Fue, en cambio por Lull y Wright, designado "de muy dudosa validez".[40] Opiniones más recientes de hadrosáuridos han coincidido.[8][35]
Durante este marco de tiempo, 1902-1915, se recuperaron dos especímenes adicionales importantes de C. annectens. La primera, la "momia de Trachodon", AMNH 5060, fue descubierta en 1908 por Charles Hazelius Sternberg y sus hijos en las rocas de la Formación Lance cerca de Lusk, Wyoming. Sternberg trabajaba para el Museo Británico de Historia Natural, pero Henry Fairfield Osborn, del Museo Americano de Historia Natural, pudo comprar el espécimen por $ 2000.[41] Los Sternbergs recuperaron un segundo espécimen similar de la misma área en 1910,[42] no tan bien conservado pero también encontrado con impresiones de piel. Vendieron este espécimen, SM 4036, al Museo Senckenberg en Alemania.[41]
Como nota al margen, Trachodon selwyni, descrito por Lawrence Lambe en 1902 por una mandíbula inferior de lo que ahora se formación Parque de Dinosaurios de Alberta,[39] fue erróneamente descrito por Glut en 1997 como asignado a Edmontosaurus regalis por Lull y Wright.[6] No fue así, en cambio lo designaron "de muy dudosa validez".[5] Las Opiniones más recientes sobre los hadrosáuridos han coincidido.[8][35]
1917-1942
Edmontosaurus en sí fue acuñado en 1917 por Lawrence Lambe para dos esqueletos parciales encontrados en la formación Cañón Herradura, anteriormente la Formación Edmonton inferior, a lo largo del Río Red Deer en el sur de Alberta, Canadá.[43] Estas rocas son más antiguas que las rocas en las que se encontró Claosaurus annectens.[9] La Formación Edmonton le da a Edmontosaurus su nombre.[29] La especie tipo, E. regalis, cuyo nombre específico proviene de "regal", o aproximadamente, "tamaño de rey",[29] se basa en NMC 2288, que consiste en un cráneo, vértebras articuladas hasta la sexta vértebra de la cola, costillas, caderas parciales, un hueso del brazo superior y la mayor parte de una extremidad posterior. Fue descubierto en 1912 por Levi Sternberg. El segundo espécimen, el paratipo NMC 2289, consiste en una calavera y un esqueleto que carecen del pico, la mayor parte de la cola y parte de los pies. Fue descubierto en 1916 por George F. Sternberg . Lambe descubrió que su nuevo dinosaurio se comparaba mejor con Diclonius mirabilis, especímenes ahora asignados a Edmontosaurus annectens, y llamó la atención sobre el tamaño y la robustez de Edmontosaurus.[43] Inicialmente, Lambe solo describió los cráneos de los dos esqueletos, pero regresó al género en 1920 para describir el esqueleto de NMC 2289.[2] El postcraneo del espécimen tipo permanece sin describir, aún en sus bochones de yeso.[6]

Dos especímenes más que llegarían a incluirse con Edmontosaurus se nombraron a partir de restos canadienses en la década de 1920, pero ambas inicialmente se asignarían a Thespesius. Gilmore nombró al primero, Thespesius edmontoni, en 1924. T. edmontoni también vino de la formación Cañón Herradura, estaba basado en NMC 8399, otro esqueleto casi completo que carecía de la cola. NMC 8399 fue descubierto en el río Red Deer en 1912 por una partida de Sternberg.[3] Lambe describió brevemente sus extremidades anteriores, tendones osificados e impresiones cutáneas en 1913 y 1914, quien al principio pensó que era un ejemplo de una especie que había llamado Trachodon marginatus,[44] pero luego cambió de opinión.[45] El espécimen se convirtió en el primer esqueleto de dinosaurio que se montará para su exhibición en un museo canadiense. Gilmore descubrió que su nueva especie se comparaba estrechamente con lo que él llamaba Thespesius annectens, pero los dejó aparte por los detalles de los brazos y las manos. También notó que su especie tenía más vértebras que la de Marsh en la espalda y cuello, pero propuso que Marsh se equivocó al suponer que los especímenes de C. annectens estaban completos en esas regiones.[3]
En 1926, Charles Mortram Sternberg nombró a Thespesius saskatchewanensis basado en NMC 8509, una calavera y un esqueleto parcial de la meseta de Wood Mountain en el sur de Saskatchewan. Él había recogido este espécimen en 1921, de rocas que fueron asignadas a la formación Lance,[4] ahora la formación Frenchman.[8] El ejemplar NMC 8509 incluía un cráneo casi completo, numerosas vértebras, cinturas parciales de hombro y cadera, y extremidades traseras parciales, que representan el primer espécimen importante de dinosaurio recuperado de Saskatchewan. Sternberg optó por asignarlo a Thespesius porque ese era el único género hadrosáurido conocido de la formación Lance en ese momento.[4] En ese momento, T. saskatchewanensis era inusual debido a su pequeño tamaño, estimado en 7 a 7,3 metros de longitud.[5]
1942-2011
En 1942, Lull y Wright intentaron resolver la complicada taxonomía de los hadrosáuridos sin cresta al nombrar un nuevo género, Anatosaurus, para incluir varias especies que no encajaban bien en sus géneros anteriores. Anatosaurus, que significa "lagarto pato", debido a su ancho pico parecido a un pato, anas del latín, "pato" y saurus del griego por "lagarto", tenía como especie tipo el antiguo Claosaurus annectens de Marsh. También se asignaron a este género Thespesius edmontoni, T. saskatchewanensis, una gran mandíbula inferior que Marsh había llamado Trachodon longiceps en 1890, y una nueva especie, Anatosaurus copei, para dos esqueletos en exhibición en el Museo Americano de Historia Natural que durante mucho tiempo se había conocido como Diclonius mirabilis o variaciones de los mismos. Por lo tanto, las diversas especies se convirtieron en A. annectens, A. copei, A. edmontoni, A. longiceps y A. saskatchewanensis.[5] Anatosaurus llegaría a llamarse el "clásico dinosaurio de pico de pato".[46]

Este estado de cosas persistió durante varias décadas, hasta que Michael K. Brett-Surman reexaminó el material pertinente para sus estudios de posgrado en los años setenta y ochenta. Concluyó que la especie tipo de Anatosaurus, A. annectens, en realidad era una especie de Edmontosaurus y que A. copei era lo suficientemente diferente como para garantizar su propio género.[47][48][49] Aunque las tesis y disertaciones no son consideradas como publicaciones oficiales por la Comisión Internacional de Nomenclatura Zoológica, que regula el nombramiento de organismos, sus conclusiones fueron conocidas por otros paleontólogos, y fueron adoptadas por varias publicaciones de divulgación por ese tiempo.[50][51] Brett-Surman y Ralph Chapman designaron un nuevo género para A. copei, Anatotitan, en 1990.[52] De las especies restantes, A. saskatchewanensis y A. edmontoni fueron asignados a Edmontosaurus también[35] y A. longiceps fue a Anatotitan, ya sea como una segunda especie[53] o como un sinónimo de A. copei.[35] Debido a que la especie tipo de Anatosaurus, A. annectens se hundió en Edmontosaurus, el nombre Anatosaurus es abandonado como un sinónimo más moderno de Edmontosaurus.
La concepción de Edmontosaurus que surgió incluía tres especies válidas: el tipo E. regalis, E. annectens, incluido Anatosaurus edmontoni, modificado a edmontonensis y E. saskatchewanensis.[35] El debate sobre la taxonomía adecuada de los especímenes de A. copei continúa hasta el presente, volviendo al argumento de Hatcher de 1902, Jack Horner, David B. Weishampel y Catherine Forster consideraron a Anatotitan copei como representantes de ejemplares de Edmontosaurus annectens con cráneos aplastados.[8] En 2007, se anunció otra "momia"; apodado "Dakota", fue descubierto en 1999 por Tyler Lyson en la formación Hell Creek de Dakota del Norte.[21][22]
Últimos estudio
En un estudio de 2011 realizado por Nicolás Campione y David Evans, los autores realizaron el primer análisis morfométrico para comparar los distintos especímenes asignados a Edmontosaurus. Concluyeron que solo dos especies son válidas, E. regalis, del Campaniano tardío, y E. annectens, del Maastrichtiense tardío. Su estudio proporcionó evidencia adicional de que Anatotitan copei es un sinónimo de E. annectens, específicamente, que el cráneo largo y bajo de A. copei es el resultado de un cambio ontogenético y representa individuos maduros de E. annectens.[9]
Especies y distribución
En la actualidad, se considera que Edmontosaurus tiene dos especies válidas, la especie tipo E. regalis y E. annectens.[8][9] E. regalis solo se conoce de la formación Cañón Herradura de Alberta, que data de finales del Campaniano a finales del Cretácico. Se conocen al menos una docena de individuos,[9] incluidos siete cráneos con material postcranial asociado, y otros cinco a siete cráneos.[8][9] La especie antes conocida como Thespesius edmontoni o Anatosaurus edmontoni representa individuos inmaduros.[9][14][54]

Se conoce a E. annectens de la formación Frenchman de Saskatchewan, la formación Hell Creek de Montana y la formación Lance de Dakota del Sur y Wyoming. Está limitado a las últimas rocas de Maastrichtiense, y está representado por al menos veinte cráneos, algunos con restos postcraneanos.[9] Un autor, Kraig Derstler, describió E. annectens como "quizás el dinosaurio más conocido hasta la fecha en 1994".[55] Ahora se piensa que Anatotitan (Anatosaurus) copei y E. saskatchewanensis son etapas de crecimiento de E. annectens, A. copei como adultos, y E. saskatchewanensis como juveniles.[9] Trachodon longiceps puede ser un sinónimo de E. annectens'" también.[8] "'Anatosaurus edmontoni fue catalogado erróneamente como sinónimo de E. annectens'" en ambas revisiones de Dinosauria,[8][35] pero este no parece ser el caso.[9][54]

E. annectens difiere de E. regalis por tener un cráneo más largo, más bajo, menos robusto.[6][9] Aunque Brett-Surman consideraba que E. regalis y E. annectens representaban potencialmente a machos y hembras de la misma especie,[47] todos los especímenes de E. regalis provienen de formaciones más antiguas que las de E. annectens.[54]
Especímenes de edmontosaurinos de la formación Prince Creek de Alaska anteriormente asignada a Edmontosaurus sp. se les ha dado su propio nombre de género y especie, Ugrunaaluk kuukpikensis.[56] Sin embargo, la identificación de Ugrunaaluk como un género separado fue cuestionada por un estudio de 2017 de Hai Xing y sus colegas, quienes lo consideraron un nomen dubium indistinguible de otros Edmontosaurus.[1] En 2020, Ryuji Takasaki y sus colegas acordaron que los restos de Prince Creek deberían clasificarse como Edmontosaurus, aunque la designación de especies no está clara ya que los especímenes son juveniles. También se informó de Edmontosaurus en la formación Javelina del parque nacional Big Bend, en el oeste de Texas, basado en TMM 41442-1,[57] pero luego se lo refirió a Kritosaurus cf. navajovius por Wagner en 2001, antes de ser asignado a Kritosaurus sp. por Lehman et al. en 2016.[58][59][60]
Clasificación
Edmontosaurus es un hadrosáurido, un miembro de la familia de los "dinosaurios de pico de pato" que proliferaron solamente a finales del período Cretácico. Es usualmente clasificado dentro de la subfamilia Saurolophinae alternativamente llamada Hadrosaurinae, un grupo de hadrosáuridos sin cresta ósea. Otros miembros de este clado incluyen a Gryposaurus, Maiasaura, Brachylophosaurus, Naashoibitosaurus, Saurolophus, Lophorhothon, y Prosaurolophus.[8] Estuvo estrechamente relacionado con[35] o incluye la especie Anatosaurus annectens, hoy conocido como Edmontosaurus annectens,[8] un gran hadrosáurido de varias de las últimas formaciones del Cretácico del oeste de América del Norte. El hadrosáurido chino gigante Shantungosaurus giganteus también es anatómicamente similar a Edmontosaurus. MK Brett-Surman encontró que los dos difieren solo en los detalles relacionados con el mayor tamaño de Shantungosaurus, con base en lo que se había descrito de este último género.[47]
Si bien el estado de Edmontosaurus dentro de Saurolophinae/Hadrosaurinae según el autor no se ha cuestionado, su ubicación exacta dentro del clado es incierta. Las filogenias tempranas, como la presentada en R. S. Lull y la influyente monografía de Nelly Wright de 1942, tenían Edmontosaurus y varias especies de Anatosaurus, la mayoría de las cuales serían consideradas más tarde como especies adicionales o especímenes de Edmontosaurus, como un linaje entre varios linajes de "hadrosáuridos de cabeza plana".[5] Uno de los primeros análisis que usaba métodos cladísticos descubrió que estaba relacionado con Anatosaurus, también llamado Anatotitan y Shantungosaurus en un clado informal de "edmontosaurinos", que fue emparejado con el "saurolophinos" con cresta de espinas y más lejanamente relacionado con los "brachylophosaurinos" y los "gryposaurinos" de hocico en arco.[35] Un estudio de 2007 realizado por Terry Gates y Scott Sampson encontró resultados ampliamente similares, en el sentido de que Edmontosaurus permaneció cerca de -Saurolophus y Prosaurolophus y distante de Gryposaurus, Brachylophosaurus y Maiasaura.[61] Sin embargo, la revisión más reciente de Hadrosauridae, por Jack Horner y colegas de 2004, llegó a un resultado notablemente diferente. Edmontosaurus estaba anidado entre Gryposaurus y los "brachylophosaurinos", y distante de Saurolophus.[8] Las discrepancias se complican por la relativa falta de trabajo en las relaciones evolutivas en Hadrosauridae.
El análisis de Godefroit et al. de 2012 incluye a las especies de Edmontosaurus junto a Kerberosaurus y Kundurosaurus en una tribu llamada Edmontosaurini. Esta última es hermana de la tribu Saurolophini, a su vez el clado formado por estas dos tribus esta más cerca de Gryposaurus y Kritosaurus que de Brachylophosaurini, todos dentro de Saurolophinae[62] La clasificación sigue el criterio del paleontólogo Albert Prieto-Márquez que emitió una publicación donde modificó a gran escala la filogenia de los hadrosáuridos, donde Hadrosaurinae fue reasigando, y solo contendrá al género Hadrosaurus y Lophorhothon y en cuestión a los "hadrosáuridos no crestados" serán incluidos en la subfamilia Saurolophinae.[63]
Filogenia

Cladograma según Horner et al. en 2004, con Edmontosaurus anidado entre Gryposaurus y lo "brachylophosauinos"[8]
| Hadrosaurinae |
| |||||||||||||||||||||||||||||||||||||||||||||

Cladograma por Gates y Sampson de 2007, con Edmontosaurus cerca a los "saurolophinos".[61]
| Hadrosaurinae |
| ||||||||||||||||||||||||||||||||||||||||||
El cladograma a continuación sigue el análisis de Godefroit et al. de 2012.[62]

| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiología
Crecimiento

En un estudio de 2011, Campione y Evans registraron datos de todos los cráneos de Edmontosaurus conocidos de Campanian y Maastrichtian y los usaron para trazar un gráfico morfométrico, comparando las características variables del cráneo con el tamaño del cráneo. Sus resultados mostraron que dentro de ambas especies de Edmontosaurus reconocidas, muchas características previamente utilizadas para clasificar especies o géneros adicionales se correlacionaron directamente con el tamaño del cráneo. Campione y Evans interpretaron estos resultados como fuertemente sugiriendo que la forma de los cráneos de Edmontosaurus cambió drásticamente a medida que crecían. Esto ha llevado a varios errores aparentes en la clasificación en el pasado. La especie del Campaniano Thespesius edmontoni, anteriormente considerada un sinónimo de E. annectens debido a su pequeño tamaño y forma de cráneo, es más probable que sea un espécimen subadulto del E. regalis contemporáneo. De manera similar, las tres especies de Edmontosaurus del Maastrichtiense previamente reconocidas probablemente representan etapas de crecimiento de una sola especie, con E. saskatchewanensis representando juveniles, subadultos de E. annectens y adultos completamente maduros de Anatotitan copei. Las calaveras se hicieron más largas y más planas a medida que los animales crecían.[9]
Cerebro y sistema nervioso

El cerebro de Edmontosaurus se ha descrito en varios documentos y resúmenes mediante el uso de moldes de la cavidad donde había estado el cerebro tanto en E. annectens[64][65] como en E. regalis,[2] así como especímenes de especies no identificados,[66][5][67] han sido estudiados de esta manera. El cerebro no era particularmente grande para un animal del tamaño de Edmontosaurus. El espacio que lo contenía era solo aproximadamente una cuarta parte de la longitud del cráneo,[2] y se han medido el volumen de estos moldes de 374 mililitros[67] a 450 mililitros,[5] que no tiene en cuenta que el cerebro puede haber ocupado tan poco como el 50% del espacio del molde, el resto del espacio ocupado por la duramadre que rodea el cerebro.[5][67] Por ejemplo, se estima que el cerebro del espécimen con el molde de 374 mililitros tuvo un volumen de 268 mililitros.[67] El cerebro era una estructura alargada[5] y cno habría habido neocorteza.[67] Al igual que el Stegosaurus, el canal neural se expandió en las caderas, pero no en el mismo grado, el espacio endosacro del Stegosaurus tenía 20 veces el volumen de su molde endocraneal, mientras que el espacio endosacro de Edmontosaurus era solo 2,59 veces mayor en volumen.[5]
Defensa e interacciones con otros dinosaurios

Algunas especies de Edmontosaurus coexistieron en el mismo lugar y tiempo con el Tyrannosaurus y un ejemplar adulto de E. annectens expuesto en el Museo de Naturaleza y Ciencia de Denver posee evidencia de una mordida de un terópodo en la cola. Contando desde la cadera hacia atrás, desde la decimotercera a la decimoséptima vértebras posee daño en las espinas vertebrales consistente con un ataque desde atrás y derecha del animal. Una espina vertebral dorsal tiene una porción arrancada, las otras están dañadas, tres tienen marcas evidentes del paso del diente. Puesto que la parte superior de la cola estaba por lo menos a 2,9 metros del suelo, el atacante tendría que haber tenido un tamaño comparable, y la única especie de terópodo conocida de la misma formación que era lo bastante alta como para acometer tal ataque era el T. rex. Los huesos están parcialmente curados, pero el Edmontosaurus murió antes de que las heridas sanaran completamente. El hueso muestra evidencias de daño por una infección. Ken Carpenter, quien estudiara el espécimen, observó que también parece haber una fractura curada en la cadera izquierda, anterior al ataque porque estaba completamente curada. Sugirió que el Edmontosaurus fuera un blanco fácil porque pudo haber cojeado debido a esta lesión anterior. Ya que sobrevivió al ataque, Carpenter sugirió que pudo esquivar o superó en velocidad a su atacante, o que el daño a su cola fue contraído por el hadrosáurido al usarla como arma contra un Tyrannosaurus,[68] Otro espécimen de E. annectens, de alrededor de 7,6 metros de Dakota del Sur, muestra marcas de dientes de pequeños terópodos en sus mandíbulas inferiores. Algunas de las marcas se curaron parcialmente. Michael Triebold, quien describiera al espécimen, sugirió un panorama donde los pequeños terópodos atacaron la garganta del edmontosaurio, el animal sobrevivió el ataque inicial pero sucumbió a sus lesiones poco después de eso.[69] Algunos lechos óseos de Edmontosaurus eran sitios de carroña. Las marcas de los dientes de Albertosaurus y Saurornitholestes son comunes en un lecho óseo de Alberta,[70] y un Daspletosaurus se alimentó de un Edmontosaurus y su compañero hadrosáurido Saurolophus en otro sitio de Alberta.[71]
Entre herbívoros contemporáneos de Cañón de la Herradura, Edmontosaurus es encontrado cerca de sedimento marino, mientras que Hypacrosaurus y Saurolophus se encuentran en sedimentos más continentales, tierra adentro.[72] Edmontosaurus y Saurolophus no se encuentran juntos comúnmente.[71] E. regalis se conoce de la más baja de cinco unidades dentro de la Formación Cañón de la Herradura, pero está ausente de por lo menos la segunda a la superior.[73] Aunque esté presentada a veces en fecha del principio del Maastrichtiense,[58] la Formación Cañón de la Herradura es más extensa, con su base aproximadamente hace 73 millones de años atrás, en finales del Campaniense.[74]
Comportamiento social
Grandes camas de huesos se conocen de Edmontosaurus, y tales agrupaciones de hadrosáuridos se utilizan para sugerir que eran gregarias, viviendo en grupos.[8] Cuatro canteras contienen restos de edmontosaurios fueron identificados en la base de datos del 2007 cama de huesos, de Alaska en la formación Prince Creek, Alberta en la formación Cañón Horseshoe, Dakota del Sur en la formación Hell Creek y Wyoming en la eormación Lance).[75]
Una cama de huesos de Edmontosaurus de la arcilla y lutolita de la formación Lance en el este de Wyoming, cubre alrededor de un kilómetro cuadrado, aunque los huesos de Edmontosaurus están concentrados en una subsección de alrededor de 40 hectáreas. Se estima que hay aproximadamente entre 10 000 a 25 000 Edmontosaurus desarticulados. Los autores sugirieron que los animales muertos primero se acumularon en un ambiente de costa de agua dulce, donde se desarticularon, y después los huesos se volvieron a mover y fueron depositados en un agua más profundas.[76]
A diferencia de otros hadrosáuridos, Edmontosaurus no tenía cresta ósea en la cabeza. Más probablemente tenía bolsas de tejido suave en el rostro, los huesos alrededor de las narinas tenían muescas profundas que rodeaban las aberturas, y este par de hendiduras se postula como para haber sostenido sacos de aire inflables, quizás teniendo en cuenta la señalización visual y auditiva.[14] Edmontosaurus debe haber presentado dimorfismo sexual, con formas más robustas y más gráciles, pero no se ha establecido si esto se relaciona con dimorfismo sexual.[77]
Debido a su distribución amplia, que cubre un territorio de Alaska a Colorado e incluye adaptaciones a las regiones polares que habrían tenido poca luz durante partes significativas del año, Edmontosaurus se lo ha considerado como un animal migratorio. Una revisión de 2008 de los estudios de la migración del dinosaurio de Phil R. Bell y de Eric Snively propuso que E. regalis era capaz de un desplazamiento anual 2600 kilómetros del viaje de ida y vuelta, con tal que tuviera el fuentes para su metabolismo y depósitos de grasa. Tal viaje tendría velocidades requeridas de cerca de 2 a 10 kilómetros por la hora y habrían podido traerlo de Alaska a Alberta. La naturaleza migratoria posible de Edmontosaurus contrasta con muchos dinosaurios, por ejemplo terópodos, saurópodos, y anquilosaurianos que Bell y Snively encontraron que eran más probables animales que hibernaran.[78][79]
Salud y patologías
En 2003, evidencias de tumores, incluyendo hemangiomas, fibromas, Metástasis, y osteoblastomas fueron descubiertos en esqueletos de Edmontosaurus. Rothschild et al. Buscaron en vértebras por tumores usando tomografía y fluoroscopios. Otros hadrosáuridos, incluyendo Brachylophosaurus, Gilmoreosaurus, y Bactrosaurus, dieron positivos. Con más de 10 000 fósiles analizados de esta manera, los tumores se limitan a Edmontosaurus y géneros cercanos. Los tumores deben haber sido causados por factores ambientales y predisposición genética.[80]
Osteocondrosis, o lesiones en los cartílagos articulares de Edmontosaurus, similares a artritis. Esta condición resulta cuando el cartílago falla y es remplazado por hueso durante el crecimiento y estuvo presente en 2,2% de 224 huesos de los dedos de Edmontosaurio. La causa subyacente de la condición es desconocida. La predisposición genética, la intensidad del trauma, la alimentación, las alteraciones en médula ósea, el exceso de hormonas tiroides, y las deficiencias en varios factores de crecimiento se han sugerido. Entre los dinosaurios, la osteocondrosis así como con tumores se encuentra más comúnmente en hadrosáuridos que en otros.[81]
Estudio de isótopos
La dieta y la fisiología de Edmontosaurus han sido estudiando usando isótopos estables de carbono y oxígeno según lo registrado en el esmalte dental. Los animales, al alimentar, beber, y respirar, absorben el carbono y el oxígeno, que se incorporan en el hueso. Los isótopos del carbono y oxígeno son determinados por varios factores internos y externos, incluyendo el tipo de plantas que son comidas, la fisiología del animal, salinidad, y el clima. Para los fósiles, si los cocientes del isótopo no son alterados por la fosilización y posterior diagénesis, pueden ser estudiados para obtener la información sobre los factores originales. Los animales de sangre caliente tendrán ciertas composiciones isotópicas dependientes de sus alrededores, los animales que comen ciertos tipos de plantas o ciertos procesos digestivos usadas tendrán composiciones isotópicas distintas, y así sucesivamente. El esmalte se utiliza típicamente porque la estructura del mineral que el esmalte lo hace el material más resistente al cambio químico en el esqueleto.[15]
Un estudio de 2004 de Kathryn Thomas y Sandra Carlson donde usaron dientes maxilares de tres individuos interpretados como un joven, un subadulto, y un adulto, recuperado de a cama de huesos en la formación Hell Creek en el condado de Corson, Dakota del Sur. En este estudio, dientes sucesivos en columnas de las baterías dentales de Edmontosaurus fueron muestreadas de localizaciones múltiples a lo largo de cada diente usando un sistema de microtaladro. Este método de muestreo se aprovecha de la organización de baterías dentales del hadrosáurido para encontrar la variación en isótopos del diente durante tiempo. Los dientes de los tres individuos aparecían demostrar la variación en los cocientes del isótopo de oxígeno que podrían corresponder a climas caliente y seco y fresco y húmedo. Thomas y Carlson consideraban la posibilidad que eran los animales hayan emigrado de un lugar a otro, solamente las variaciones estacionales locales favorecidas porque la migración habría llevado más probablemente a la homogeneización de cocientes, tantos animales emigran para permanecer dentro de gamas de temperaturas específicas o para acercar a fuentes particulares del alimento. Los dientes habrían tomado menos de 0,65 años para formarse, levemente más rápida en Edmontosaurus más jóvenes.[15]
Los Edmontosaurus también mostraron valores enriquecidos del isótopo de carbono, que para los mamíferos modernos serían interpretados como dieta mezclada de plantas C3, la mayoría de las plantas y plantas C4, las gramíneas. Sin embargo, las plantas C4 eran extremadamente raras en el Cretácico. Thomas y Carlson propusieron varios factores que pudieron haber funcionado, y encontrar el más probable incluir una dieta de muchas gimnospermas, consumiendo las plantas saladas de zonas costeras adyacente la vía marítima interior occidental, y diferencia fisiológica entre los dinosaurios y los mamíferos que hicieron dinosaurios formar el tejido con distintos cocientes del carbono que sería el esperado para los mamíferos. Una combinación de factores es también posible.[15]
Adaptaciones de alimentación
Como la mayoría de los hadrosáuridos, Edmontosaurus tenía una cabeza que hace recordar a la de los modernos patos. El frente de la cabeza era plano y redondeado, extendiéndose en un pico córneo que era ideal para recortar las ramitas y hojas de los bosques de Norteamérica. En Edmontosaurus sus dientes fueron reemplazados continuamente y empacados en baterías dentales que contenían cientos de dientes, de los cuales solo un puñado relativo estaba en uso en al mismo tiempo.[8] Utilizó su pico ancho para cortar alimentos a granel, tal vez mediante como cosechando,[8] o al cerrar las mandíbulas en forma de concha sobre ramitas y ramas y luego quitar las hojas y brotes más nutritivos.[17] Debido a que las filas dentales están profundamente colocadas desde el exterior de las mandíbulas, y debido a otros detalles anatómicos, se deduce que Edmontosaurus y la mayoría de los otros ornitisquios tenían estructuras parecidas a las mejillas, musculares o no musculares. La función de las mejillas era retener la comida en la boca.[82][83] El rango de alimentación del animal habría sido desde el nivel del suelo a alrededor de 4 metros por encima del suelo.[8]
Antes de los años 1960 y 1970, la interpretación predominante de hadrosáuridos como Edmontosaurus era que eran acuáticos y se alimentaban de plantas acuáticas.[84] Un ejemplo de esto es la interpretación de William Morris de 1970 de un cráneo Edmontosaurus con restos de pico no óseo. Propuso que el animal tenía una dieta muy similar a la de algunos patos modernos, plantas de filtración e invertebrados acuáticos como moluscos y crustáceos del agua y descargando agua a través de surcos en forma de V a lo largo de la cara interna del pico superior.[11] Esta interpretación del pico ha sido rechazada, ya que los surcos y crestas son más parecidos a los de los picos de las tortugas herbívoras que las estructuras flexibles que se ven en las aves que se alimentan por filtración.[84]
Entre mediados de los años ochenta y la primera década de la década de 2000, la interpretación prevaleciente de cómo los hadrosáuridos procesaban sus alimentos siguió el modelo presentado en 1984 por Propuso que la estructura del cráneo permitiera el movimiento entre los huesos de la mandíbula inferior que iban de atrás hacia adelante y la inclinación hacia fuera de las baterías de dientes de la superior cuando la boca era cerrada. Los dientes de la parte superior molerían contra los dientes de la inferior como escofinas, procesando material vegetal atrapado entre ellos..[85][8] Este movimiento en análogo a la masticación de los mamíferos, aunque se produzca por una vía diferente.[86] Un trabajo en la década de 2000 desafió el modelo de Weishampel. Un estudio publicado en 2008 por Casey Holliday y Lawrence Witmer descubrió que los ornitópodos como Edmontosaurus carecían de los tipos de cráneos que se ven en los animales modernos que se sabe que tienen cráneos cinéticos, cráneos que permiten el movimiento entre los huesos constituyentes, como los escamosos y las aves. Propusieron que las articulaciones que se habían interpretado como que permitían el movimiento en los cráneos de los dinosaurios eran en realidad zonas de crecimiento cartilaginosos.[87] Una importante pieza de evidencia para el modelo de Weishampel es la orientación en las raspaduras en los dientes, mostrando la dirección de la acción de la mandíbula. Sin embargo, otros movimientos podrían producir arañazos similares, como el movimiento de los huesos de las dos mitades de la mandíbula inferior. No todos los modelos han sido examinados bajo las técnicas actuales.[87] Vincent Williams y sus colegas en 2009 publicaron trabajos adicionales sobre el micro desgaste en los dientes de hadrosáuridos. Encontraron cuatro clases de rasguños en dientes de Edmontosaurus. La clase más común se interpretó como resultado de un movimiento oblicuo, no de un simple movimiento hacia arriba o hacia atrás, que es consistente con el modelo de Weishampel. Se cree que este movimiento fue el movimiento principal para moler alimentos. Se interpretaron otras dos clases de rasguños como resultado del movimiento hacia delante o hacia atrás de las mandíbulas. La otra clase fue variable y probablemente resultó de abrir las mandíbulas. La combinación de movimientos es más compleja de lo que se había predicho previamente.[88]

Weishampel desarrollo su modelo con la asistencia de la animación por computadora. Natalia Rybczynski y colegas han actualizado el modelo con el uso de imágenes de tres dimensiones en el, escaseando un cráneo de E. regalis con láser. Podían replicar el movimiento propuesto con su modelo, aunque encontraron que los movimientos secundarios adicionales entre otros huesos fueron requeridos, con separaciones máximas de 1,3 a 1,4 centímetros entre algunos huesos durante el ciclo de masticación. Rybczynski y los colegas no quedaron convencidos de que el modelo de Weishampel es viable, pero observaron que tienen varias mejoras a ejecutar a su animación. Las mejoras previstas incluyen la incorporación de tejido blando y las marcas y rasguños del desgaste de dientes, lo que debería limitar los movimientos. Apuntan que hay varias otras hipótesis a probar también.[86] La investigación adicional publicada en 2012 por Robin Cuthbertson y sus colegas encontraron que los movimientos requeridos para el modelo de Weishampel eran poco probables, y favorecían un modelo en el que los movimientos de la mandíbula inferior producían una acción de molienda. La articulación de la mandíbula inferior con la mandíbula superior permitiría un movimiento anteroposterior junto con la rotación habitual, y la articulación anterior de las dos mitades de la mandíbula inferior también permitiría el movimiento, en combinación, las dos mitades de la mandíbula inferior podrían moverse ligeramente hacia adelante y hacia atrás, así como girar ligeramente a lo largo de sus ejes largos. Estos movimientos explicarían el desgaste dental observado y un cráneo más sólido que el modelado por Weishampel.[89] Debido a que los arañazos dominan la textura del micro desgaste de los dientes, Williams et al. sugirieron que Edmontosaurus era un animal de pastoreo en lugar de un ramoreador, que se podría predecir que tendría menos arañazos debido a que comería menos materiales abrasivos. Los candidatos para los alimentos abrasivos ingeridos incluyen plantas ricas en sílice como las cola de caballo y el suelo que se ingirió accidentalmente debido a la alimentación a nivel del suelo.[88] La estructura del diente indica capacidades combinadas de corte y molienda.[16]
Los informes de gastrolitos o cálculos estomacales en el hadrosáurido Claosaurus en realidad se basan en una probable identificación errónea. Primero, el espécimen es en realidad un Edmontosaurus annectens. Barnum Brown, que descubrió el espécimen en 1900, se refirió a él como Claosaurus porque se creía que E. annectens era una especie de Claosaurus en ese momento. Además, es más probable que los supuestos gastrolitos representen grava lavada durante el entierro.[29]
Contenido intestinal
Se informó que los dos especímenes "momia" recolectados por los Sternberg tenían posibles contenidos intestinales. Charles H. Sternberg informó la presencia de contenido de intestino carbonizado en el espécimen del Museo Americano de Historia Natural,[90] pero este material no ha sido descrito.[91] Los restos de plantas en el espécimen del Museo Senckenberg han sido descritos, pero han demostrado ser difíciles de interpretar. Las plantas que se encuentran en el cadáver incluyen agujas de la conífera Cunninghamites elegans, ramitas de coníferas y árboles de hoja ancha y numerosas semillas pequeñas o frutas.[92] Tras su descripción en 1922, fueron objeto de un debate en la revista en lengua alemana Paläontologische Zeitschrift. Kräusel, quien describió el material, lo interpretó como el contenido del intestino del animal,[92] mientras que Abel no pudo descartar que las plantas hubieran sido arrastradas al cadáver después de la muerte.[93]
En ese momento, se pensaba que los hadrosáuridos eran animales acuáticos, y Kräusel insistió en que el espécimen no descartaba que los hadrosáuridos consumieran plantas acuáticas.[17][92] El descubrimiento de posibles contenidos intestinales tuvo poco impacto en los círculos de habla inglesa, a excepción de otra breve mención de la dicotomía acuático-terrestre,[94] hasta que fue mencionado por John Ostrom en el curso de un artículo revaluar la antigua interpretación de los hadrosáuridos como acuáticos. En lugar de intentar adaptar el descubrimiento al modelo acuático, lo usó como una evidencia de que los hadrosáuridos eran herbívoros terrestres.[17] Si bien su interpretación de los hadrosáuridos como animales terrestres ha sido generalmente aceptada,[8] los fósiles de plantas de Senckenberg siguen siendo equívocos. Kenneth Carpenter ha sugerido que pueden representar el contenido intestinal de un animal muerto de hambre, en lugar de una dieta típica.[95][96] Otros autores han notado que debido a que los fósiles de plantas fueron removidos de su contexto original en la muestra y estaban fuertemente preparados, ya no es posible hacer un seguimiento de la obra original, dejando abierta la posibilidad de que las plantas fuesen restos arrastrados.[91][97]
Locomoción
Edmontosaurus era bípedo pero con certeza pudo caminar en cuatro patas. Sus brazos aunque más cortos que sus patas, no lo eran tanto como para impedir que los usara para desplazarse. Las manos tenían los dedos formando pezuñas en dos dedos, y cojines. Los pies tenían tres dedos del pie funcionales y todos eran ungulados. La estructura del hueso en los miembros traseros sugiere que en las piernas y los pies se insertaban músculos muy grandes. La espina dorsal se curvaba hacia abajo en los hombros, tanto que Edmontosaurus habría tenido una postura baja y se habría alimentado cerca de la tierra. El trabajo reciente conducido por investigadores de computación, en 2007, sugiere que Edmontosaurus podía correr a altas velocidades, quizás hasta 12,5 m/s, alrededor de 45 km/h.[21]
Al igual que otros hadrosáuridos, se cree que Edmontosaurus fue un bípedo facultativo, lo que significa que se movió principalmente en cuatro patas, pero podría adoptar una postura bípeda cuando sea necesario. Probablemente se ponía a cuatro patas cuando se detenía o se movía lentamente, y cambiaba a usar solo las patas traseras cuando se movía más rápido.[8] La investigación realizada por modelado por computadora en 2007 sugiere que Edmontosaurus podría correr a altas velocidades, quizás hasta 45 kilómetros por hora.[21] Simulaciones adicionales usando un espécimen de subadulto estimado como un peso de 715 kilogramos cuando estaba vivo produjeron un modelo que podía correr o saltar como bípedo, usar un trote, paso, o modo cuadrúpedo simétrico de pie único, o moverse al galope. Los investigadores descubrieron con sorpresa que el modo de andar más rápido era el salto tipo canguro, con una velocidad máxima simulada de 17,3 m/s, unos 62 km/h, que consideraron improbable en función del tamaño del animal y la falta de huellas saltando en el registro fósil, y en su lugar se interpretó el resultado como indicativo de una inexactitud en su simulación. Los modos más rápidos de no saltos era el galope con una velocidad máxima simulada de 15,7 m/s, unos 57 km/h y el que se ejecutaba en forma bípeda, velocidad máxima simulada de 14,0 m/s, unos 50 km/h. Encontraron un soporte débil para la carrera bípeda como la opción más probable para el movimiento a alta velocidad, pero no descartaron el movimiento cuadrúpedo de alta velocidad.[98] Aunque durante mucho tiempo se pensó que eran acuáticos o semiacuáticos, los hadrosáuridos no eran tan aptos para nadar como otros dinosaurios, especialmente los terópodos, que alguna vez se pensó que no podían perseguir a los hadrosáuridos en el agua. Los hadrosáuridos tenían manos delgadas con dedos cortos, haciendo que sus extremidades anteriores fueran ineficaces para la propulsión, y la cola tampoco era útil para la propulsión debido a los tendones osificados que aumentaban su rigidez y los puntos de unión poco desarrollados para los músculos que habrían movido la cola desde el costado a un lado.[99][100]
Paleoecología
Edmontosaurus es un género de amplia distribución en tiempo y espacio. Las unidades de rocas de las que se la conoce se pueden dividir en dos grupos por edad, las más antiguas formaciones Cañón Herradura y St. Mary River, y las formaciones más jóvenes de Frenchman, Hell Creek y Lance. El lapso de tiempo cubierto por la Formación Cañón Herradura y equivalentes también se conoce como Edmontoniano, y el lapso de tiempo cubierto por las unidades más jóvenes también se conoce como Lanciano. Los intervalos de tiempo de Edmontoniano y Lanciano tenían distintas faunas de dinosaurios.[101]

La edad Edmontoniana de los vertebrados terrestres se define por la primera aparición de Edmontosaurus regalis en el registro fósil.[102] Aunque a veces se informa como exclusivamente del Maastrichtiano temprano, [60] la Formación Horseshoe Canyon fue de una duración algo mayor. La deposición comenzó hace aproximadamente 73 millones de años, a fines del Campaniano, y terminó entre 68.0 y 67.6 millones de años atrás.[74] Edmontosaurus regalis es conocido desde la más baja de las cinco unidades dentro de la Formación Horseshoe Canyon, pero está ausente al menos desde el segundo hasta la cima.[73] Hasta tres cuartos de los ejemplares de dinosaurios de las tierras baldías cerca de Drumheller, Alberta puede pertenecer a Edmontosaurus.[103]
El intervalo de tiempo Lanciano fue el último intervalo antes del evento de extinción Cretácico-Paleógeno que eliminó los dinosaurios no aviares. Edmontosaurus fue uno de los dinosaurios más comunes del intervalo. Robert Bakker informa que constituía un séptimo de la gran muestra de dinosaurio, con la mayor parte del resto, cinco sextos, formado por el dinosaurio cornudo Triceratops.[104] La llanura costera de Triceratops en asociación con Edmontosaurus, dominada por Triceratops, se extendió desde Colorado a Saskatchewan.[105]

La formación Lance, como se tipifica por las exposiciones a aproximadamente 100 kilómetros al norte de Fort Laramie en el este de Wyoming, se ha interpretado como un entorno de pantano similar a la llanura costera de Luisiana. Estaba más cerca de un gran delta que la configuración de depósito de la formación Hell Creek al norte y recibió mucho más sedimento. Coníferas, palmeras y araucarias tropicales salpican los bosques de frondosas, diferenciando la flora de la llanura costera del norte.[106] El clima era húmedo y subtropical, con coníferas, palmitos y helechos en los pantanos y coníferas, fresnos, robles y arbustos en los bosques.[55] Los peces de agua dulce, las salamandras, las tortugas, diversos lagartos, serpientes, aves playeras y pequeños mamíferos vivían junto a los dinosaurios. Los pequeños dinosaurios no son tan abundantes aquí como en las rocas de Hell Creek, pero Thescelosaurus una vez más parece haber sido relativamente común. Triceratops es conocido por muchos cráneos, que tienden a ser algo más pequeños que los de individuos más del norte. La Formación Lance es el escenario de dos "momias" de Edmontosaurus.[106]
En la cultura popular
Gracias a la enorme abundancia de fósiles encontrados, Edmontosaurus es uno de los dinosaurios más conocidos por los paleontólogos que, en consecuencia, han podido reconstruir toda su ecología, desde la forma en que migraban hasta el lugar donde se alimentaban. Por esta razón, fue el protagonista de varios documentales, series de televisión y especiales sobre dinosaurios. La especie E. annectens, llamada también Anatotitan, aparece en el último capítulo de la serie de documentales de la BBC Walking with Dinosaurs, " "Muerte de una Dinastía" (Death of a Dinasty) ".[107] El director y productor Jasper James decidió representar a E. annectens como el último género de dinosaurio pico de pato cazado por el terópodo Tyrannosaurus rex. Edmontosaurus también aparece en el documental When Dinosaurs Roamed America como un pacífico dinosaurio herbívoro. E. regalis, por su parte, aparece en el documental March of the Dinosaurs de 2011, donde el protagonista es un joven ejemplar llamado Cicatriz, quien debe sobrevivir a la dura migración de sus compañeros; haciendo igualmente su debut en solitario en los medios.
Edmontosaurus también ha aparecido en series y películas, siendo las más destacadas el anime Dino Rey, tanto la especie tipo E. regalis como E. annectens aparecen en el juego del cual se basa la serie, siendo este el único medio que mostró ambas especies de Edmontosaurus en los medios. E. annectens está presente como una de las especies representadas en Dino Dan. También en la película Walking with Dinosaurs del 2013, aparecen de forma breve una manada de E. annectens, lo cual es incorrecto, ya que este último vivió desde Saskatchewan (al sur de Canadá) hasta Colorado (EE. UU.), mientras que E. regalis vivió en Alaska y en norte de Canadá, dónde se ubican los eventos del filme. Una imagen promocional de E. regalis (otra vez con el error anatómico sin cresta) aparece en el sitio promocional de Jurassic World, pero el animal no apareció en la película final, excepto con un breve cameo en una escena eliminada. Un esqueleto de E. annectens aparece dentro de la mansión de Benjamin Lockwood en Jurassic World: El reino caído. Edmontosaurus ha aparecido también en videojuegos; teniendo participación aparte de Dino Rey, también está presente por ejemplo en los juegos de Jurassic Park, de los más recientes, E. regalis es una de las especies representadas en Jurassic World Evolution. Por otro lado, E. annectens ha aparecido recientemente en Saurian y en Archotek Project, bajo el sinónimo Anatosaurus.
Referencias
- Xing, H.; Mallon, J.C.; Currie, M.L. (2017). «Supplementary cranial description of the types of Edmontosaurus regalis (Ornithischia: Hadrosauridae), with comments on the phylogenetics and biogeography of Hadrosaurinae». PLoS ONE 12 (4): e0175253. Bibcode:2017PLoSO..1275253X. doi:10.1371/journal.pone.0175253.
- Lambe, Lawrence M. (1920). The hadrosaur Edmontosaurus from the Upper Cretaceous of Alberta. Memoir 120. Department of Mines, Geological Survey of Canada. pp. 1-79.
- Gilmore, Charles W. (1924). A new species of hadrosaurian dinosaur from the Edmonton Formation (Cretaceous) of Alberta. Bulletin 38. Department of Mines, Geological Survey of Canada. pp. 13-26.
- Sternberg, Charles M. (1926). A new species of Thespesius from the Lance Formation of Saskatchewan. Bulletin 44. Department of Mines, Geological Survey of Canada. pp. 77-84.
- Lull, Richard Swann; Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. Geological Society of America Special Paper 40. Geological Society of America.
- Glut, Donald F. (1997). «Edmontosaurus». Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co. pp. 389–396. ISBN 0-89950-917-7.
- Lambert, David; and the Diagram Group (1990). The Dinosaur Data Book. Nueva York: Avon Books. p. 60. ISBN 0-380-75896-2.
- Horner, John R.; Weishampel, David B.; Forster, Catherine A (2004). «Hadrosauridae». En Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.), ed. The Dinosauria (2nd edición). Berkeley: University of California Press. pp. 438-463. ISBN 0-520-24209-2.
- Campione, N.E.; Evans, D.C. (2011). «Cranial Growth and Variation in Edmontosaurs (Dinosauria: Hadrosauridae): Implications for Latest Cretaceous Megaherbivore Diversity in North America». PLoS ONE 6 (9): e25186. Bibcode:2011PLoSO...625186C. PMC 3182183. PMID 21969872. doi:10.1371/journal.pone.0025186.
- Lucas, Frederic A. (2004). «The dinosaur Trachodon annectens». Smithsonian Miscellaneous Collections 45: 317-320.
- Morris, William J. (1970). «Hadrosaurian dinosaur bills — morphology and function». Contributions in Science (Los Angeles County Museum of Natural History) 193: 1-14.
- Horner, John R.; Goodwin, Mark B.; Myhrvold, Nathan (2011). «Dinosaur Census Reveals Abundant Tyrannosaurus and Rare Ontogenetic Stages in the Upper Cretaceous Hell Creek Formation (Maastrichtian), Montana, USA.». PLoS ONE 6 (2): e16574. Bibcode:2011PLoSO...616574H. PMC 3036655. PMID 21347420. doi:10.1371/journal.pone.0016574.
- Cope, Edward D. (1883). «On the characters of the skull in the Hadrosauridae». Proceedings of the Philadelphia Academy of Natural Sciences 35: 97-107.
- Hopson, James A. (1975). «The evolution of cranial display structures in hadrosaurian dinosaurs». Paleobiology 1 (1): 21-43.
- Stanton Thomas, Kathryn J.; Carlson, Sandra J. (2004). «Microscale δ18O and δ13C isotopic analysis of an ontogenetic series of the hadrosaurid dinosaur Edmontosaurus: implications for physiology and ecology». Palaeogeography, Palaeoclimatology, and Palaeoecology 206 (2004): 257-287. doi:10.1016/j.palaeo.2004.01.007.
- Erickson, Gregory M.; Krick, Brandon A.; Hamilton, Matthew; Bourne, Gerald R.; Norell, Mark A.; Lilleodden, Erica; Sawyer, W. Gregory. (2012). «Complex dental structure and wear biomechanics in hadrosaurid dinosaurs» (Submitted manuscript). Science 338 (6103): 98-101. Bibcode:2012Sci...338...98E. PMID 23042891. doi:10.1126/science.1224495.
- Ostrom, John H.. «A reconsideration of the paleoecology of the hadrosaurian dinosaurs». American Journal of Science 262: 975-997.
- Galton, Peter M. (1970). «The posture of hadrosaurian dinosaurs». Journal of Paleontology 44 (3): 464-473.
- Osborn, Henry Fairfield (1909). «The epidermis of an iguanodont dinosaur». Science 29 (750): 793-795. Bibcode:1909Sci....29..793F. PMID 17787819. doi:10.1126/science.29.750.793.
- Osborn, Henry Fairfield (1912). «Integument of the iguanodont dinosaur Trachodon» (pdf (very large; 76,048 kb)). Memoirs of the American Museum of Natural History 1: 33-54. Consultado el 8 de marzo de 2009.
- Mummified Dinosaur Unveiled. National Geographic News. 2007-12-03. Consultado el 3 de diciembre de 2007.
- Lee, Christopher (3 de diciembre de 2007). «Scientists Get Rare Look at Dinosaur Soft Tissue». Washington Post. Consultado el 3 de diciembre de 2007.
- Manning, Phillip L.; Morris, Peter M.; McMahon, Adam; Jones, Emrys; Gize, Andy; Macquaker, Joe H. S.; Wolff, G.; Thompson, Anu; Marshall, Jim; Taylor, Kevin G.; Lyson, Tyler; Gaskell, Simon; Reamtong, Onrapak; Sellers, William I.; van Dongen, Bart E.; Buckley, Mike; and Wogelius, Roy A. (2009). «Mineralized soft-tissue structure and chemistry in a mummified hadrosaur from the Hell Creek Formation, North Dakota (USA)». Proceedings of the Royal Society B 276 (1672): 3429-3437. PMC 2817188. PMID 19570788. doi:10.1098/rspb.2009.0812.
- Bell, P. R.; Fanti, F.; Currie, P. J.; Arbour, V.M. (2013). «A Mummified Duck-Billed Dinosaur with a Soft-Tissue Cock's Comb». Current Biology 24 (1): 70-75. PMID 24332547. doi:10.1016/j.cub.2013.11.008.
- Lyson, Tyler R., Hanks, H. Douglas, and Tremain, Emily S. (2003) "New skin structures from a juvenile Edmontosaurus from the Late Cretaceous of North Dakota Archivado el 24 de febrero de 2016 en Wayback Machine.". Geological Society of America. Last accessed 2008-08-02.
- https://blogs.scientificamerican.com/tetrapod-zoology/enough-with-the-duckbilled-dinosaurs/
- https://www.smithsonianmag.com/science-nature/shovel-beaked-not-duck-billed-122016600/
- J, Morris, William (1970). «Hadrosaurian dinosaur bills — morphology and function». Contributions in Science (en inglés) 193. ISSN 0459-8113.
- Creisler, Benjamin S. (2007). «Deciphering duckbills: a history in nomenclature». En Carpenter, Kenneth (ed.), ed. Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs. Bloomington and Indianapolis: Indiana University Press. pp. 185-210. ISBN 0-253-34817-X.
- Marsh, Othniel Charles (1892). «Notice of new reptiles from the Laramie Formation». American Journal of Science 43 (257): 449-453. Bibcode:1892AmJS...43..449M. doi:10.2475/ajs.s3-43.257.449.
- Marsh, Othniel Charles (1892). «Restorations of Claosaurus and Ceratosaurus» (Submitted manuscript). American Journal of Science 44 (262): 343-349. doi:10.2475/ajs.s3-44.262.343.
- Hatcher, John B. (1902). «The genera and species of the Trachodontidae (Hadrosauridae, Claosauridae) Marsh». Annals of the Carnegie Museum 1 (14): 377-386.
- Gilmore, Charles W. (1915). «On the genus Trachodon». Science 41 (1061): 658-660. Bibcode:1915Sci....41..658G. PMID 17747979. doi:10.1126/science.41.1061.658.
- Cope, Edward Drinker (1871). «Supplement to the synopsis of the extinct Batrachia and Reptilia of North America». American Philosophical Society, Proceedings 12 (86): 41-52.
- Horner, John R.; Weishampel, David B.; Forster, Catherine A (2004). «Hadrosauridae». En Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.), ed. The Dinosauria (2nd edición). Berkeley: University of California Press. pp. 438–463. ISBN 0-520-24209-2.
- Cope, Edward Drinker (1874). «Report on the stratigraphy and Pliocene vertebrate paleontology of northern Colorado». U.S. Geological and Geographical Survey of the Territories Annual Report 1: 9-28.
- Carpenter, Kenneth; Young, D. Bruce (2002). «Late Cretaceous dinosaurs from the Denver Basin, Colorado». Rocky Mountain Geology 37 (2): 237-254. doi:10.2113/11.
- Cope, Edward Drinker (1874). «Report on the vertebrate paleontology of Colorado». U.S. Geological and Geographical Survey of the Territories Annual Report 2: 429-454.
- Lambe, Lawrence M. (1902). «On Vertebrata of the mid-Cretaceous of the Northwest Territory. 2. New genera and species from the Belly River Series (mid-Cretaceous)». Contributions to Canadian Paleontology 3: 25-81.
- Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. pp. 220–221.
- Norell, M. A.; Gaffney, E. S.; Dingus, L. (1995). Discovering Dinosaurs in the American Museum of Natural History. New York: Knopf. pp. 154–155. ISBN 978-0-679-43386-6.
- Dal Sasso, Cristiano; Brillante, Giuseppe (2004). Dinosaurs of Italy. Bloomington and Indianapolis: Indiana University Press. p. 112. ISBN 978-0-253-34514-1.
- Lambe, Lawrence M. (1917). «A new genus and species of crestless hadrosaur from the Edmonton Formation of Alberta» (pdf (entire volume, 18 mb)). The Ottawa Naturalist 31 (7): 65-73. Consultado el 8 de marzo de 2009.
- Lambe, Lawrence M. (1913). «The manus in a specimen of Trachodon from the Edmonton Formation of Alberta». The Ottawa Naturalist 27: 21-25.
- Lambe, Lawrence M. (1914). «On the fore-limb of a carnivorous dinosaur from the Belly River Formation of Alberta, and a new genus of Ceratopsia from the same horizon, with remarks on the integument of some Cretaceous herbivorous dinosaurs». The Ottawa Naturalist 27: 129-135.
- Glut, Donald F. (1982). The New Dinosaur Dictionary. Secaucus, NJ: Citadel Press. p. 57. ISBN 978-0-8065-0782-8.
- Brett-Surman, Michael K. (1989). A revision of the Hadrosauridae (Reptilia: Ornithischia) and their evolution during the Campanian and Maastrichtian. Ph.D. dissertation. Washington, D.C.: George Washington University.
- Brett-Surman, Michael K. (1975). The appendicular anatomy of hadrosaurian dinosaurs. M.A. thesis. Berkeley: University of California.
- Brett-Surman, Michael K. (1979). «Phylogeny and paleobiogeography of hadrosaurian dinosaurs» (Submitted manuscript). Nature 277 (5697): 560-562. Bibcode:1979Natur.277..560B. doi:10.1038/277560a0.
- Glut, Donald F. (1982). The New Dinosaur Dictionary. Secaucus, NJ: Citadel Press. pp. 49, 53. ISBN 978-0-8065-0782-8.
- Lambert, David; the Diagram Group (1983). A Field Guide to Dinosaurs. New York: Avon Books. pp. 156–161. ISBN 978-0-380-83519-5.
- Chapman, Ralph E.; Brett-Surman, Michael K. (1990). «Morphometric observations on hadrosaurid ornithopods». En Carpenter, Kenneth; Currie Philip J. (eds.), eds. Dinosaur Systematics: Perspectives and Approaches. Cambridge: Cambridge University Press. pp. 163–177. ISBN 978-0-521-43810-0.
- Olshevsky, George. (1991). A Revision of the Parainfraclass Archosauria Cope, 1869, Excluding the Advanced Crocodylia. Mesozoic Meanderings No. 2. San Diego: Publications Requiring Research.
- Campione, N.E. (2009). "Cranial variation in Edmontosaurus (Hadrosauridae) from the Late Cretaceous of North America." North American Paleontological Convention (NAPC 2009): Abstracts, p. 95a. PDF link
- Derstler, Kraig (1994). «Dinosaurs of the Lance Formation in eastern Wyoming». En Nelson Gerald E. (ed.), ed. The Dinosaurs of Wyoming. Wyoming Geological Association Guidebook, 44th Annual Field Conference. Wyoming Geological Association. pp. 127-146.
- Mori, Hirotsugu; Druckenmiller, Patrick S.; Erickson, Gregory M. (2015). «A new Arctic hadrosaurid from the Prince Creek Formation (lower Maastrichtian) of northern Alaska». Acta Palaeontologica Polonica 61: . doi:10.4202/app.00152.2015.
- Davies, K.L., 1983, Hadrosaurian dinosaurs of Big Bend National Park [M.A. thesis]: Austin, University of Texas, 231 p.
- Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loueff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth M.P. et al. (2004). «Dinosaur distribution». En Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.), ed. The Dinosauria (2nd edición). Berkeley: University of California Press. pp. 517–606. ISBN 0-520-24209-2.
- Wagner, J.R. 2001. The hadrosaurian dinosaurs (Ornithischia: Hadrosauria) of Big Bend National Park, Brewster County, Texas, with implications for Late Cretaceous Paleozoogeography. M.Sc. thesis, Texas Tech University, Austin, Tex.
- Lehman, Thomas M.; Wick, Steven L.; Wagner, Jonathan R. (2016). «Hadrosaurian dinosaurs from the Maastrichtian Javelina Formation, Big Bend National Park, Texas». Journal of Paleontology 90 (2): 1-24. doi:10.1017/jpa.2016.48.
- Gates, Terry A.; Sampson, Scott D. (2007). «A new species of Gryposaurus (Dinosauria: Hadrosauridae) from the late Campanian Kaiparowits Formation, southern Utah, USA». Zoological Journal of the Linnean Society 151 (2): 351-376. doi:10.1111/j.1096-3642.2007.00349.x.
- Godefroit, P.; Bolotsky, Y. L.; Lauters, P. (2012). «A New Saurolophine Dinosaur from the Latest Cretaceous of Far Eastern Russia». PLoS ONE 7 (5): e36849. PMC 3364265. PMID 22666331. doi:10.1371/journal.pone.0036849.
- Prieto-Márquez, A. (2010). «Global phylogeny of Hadrosauridae (Dinosauria: Ornithopoda) using parsimony and Bayesian methods». Zoological Journal of the Linnean Society 159 (2): 435-502. doi:10.1111/j.1096-3642.2009.00617.x.
- Marsh, Othniel Charles (1893). «The skull and brain of Claosaurus». American Journal of Science 45 (265): 83-86. Bibcode:1893AmJS...45...83M. doi:10.2475/ajs.s3-45.265.83.
- Marsh, Othniel Charles (1896). «The dinosaurs of North America». Sixteenth Annual report of the United States Geological Survey to the Secretary of the Interior, 1894–1895: Part 1. Washington, D.C.: U.S. Geological Survey. pp. 133-244. Archivado desde el original el 9 de mayo de 2009. Consultado el 8 de marzo de 2009.
- Brown, Barnum (1914). «Anchiceratops, a new genus of horned dinosaurs from the Edmonton Cretaceous of Alberta, with discussion of the ceratopsian crest and the brain casts of Anchiceratops and Trachodon». Bulletin of the American Museum of Natural History 33: 539-548.
- Jerison, Harry J.; Horner, John R.; Horner, Celeste C (2001). «Dinosaur forebrains». Journal of Vertebrate Paleontology 21 (3, Suppl.): 64A. doi:10.1080/02724634.2001.10010852.
- Carpenter, Kenneth (1998). «Evidence of predatory behavior by theropod dinosaurs.». Gaia 15: 135-144. Archivado desde el original el 17 de noviembre de 2007. [not printed until 2000]
- Campagna, Tony (2000). «The PT interview: Michael Triebold». Prehistoric Times 40: 18-19.
- Jacobsen, Aase Roland; Ryan, Michael J. (2000). «Taphonomic aspects of theropod tooth-marked bones from an Edmontosaurus bone bed (Lower Maastrichtian), Alberta, Canada». Journal of Vertebrate Paleontology 19 (3, suppl.): 55A. doi:10.1080/02724634.1999.10011202.
- «City Site Was Dinosaur Dining Room». ScienceDaily. ScienceDaily. 3 de julio de 2007. Consultado el 7 de diciembre de 2008.
- Russell, Dale A.; Chamney, T. P. (1967). «Notes on the biostratigraphy of dinosaurian and microfossil faunas in the Edmonton Formation (Cretaceous), Alberta». National Museum of Canada Natural History Papers 35: 1-35.
- Eberth, David A. (2002). «Review and comparison of Belly River Group and Edmonton Group stratigraphy and stratigraphic architecture in the southern Alberta Plains» (pdf). Canadian Society of Petroleum Geology Diamond Jubilee Convention, Programs and Abstracts 117: (cd). Archivado desde el original el 13 de noviembre de 2007. Consultado el 8 de enero de 2009.
- Wu, X-C.; Brinkman, D.B.; Eberth, D.A.; Braman, D.R. (2007). «A new ceratopsid dinosaur (Ornithischia) from the uppermost Horseshoe Canyon Formation (upper Maastrichtian), Alberta, Canada». Canadian Journal of Earth Science 44 (9): 1243-1265.
- Behrensmeyer, Anna K. (2007). «Evolution of Terrestrial Ecosystems Bonebed Database» (Excel spreadsheet). Bonebeds: Genesis, Analysis, and Paleobiological Significance. University of Chicago Press. Consultado el 7 de diciembre de 2008.
- Chadwick, Arthur; Spencer, Lee, and Turner, Larry (2006). «Preliminary depositional model for an Upper Cretaceous Edmontosaurus bonebed». Journal of Vertebrate Paleontology 26 (3, suppl.): 49A.
- Gould, Rebecca; Larson, Robb; Nellermoe, Ron (2003). «An allometric study comparing metatarsal IIs in Edmontosaurus from a low-diversity hadrosaur bone bed in Corson Co., SD». Journal of Vertebrate Paleontology 23 (3, suppl.): 56A-57A.
- Bell, Phil R.; Snively, E. (2008). «Polar dinosaurs on parade: a review of dinosaur migration». Alcheringa 32 (3): 271-284. doi:10.1080/03115510802096101.
- Lloyd, Robin (4 de diciembre de 2008). «Polar Dinosaurs Endured Cold Dark Winters». LiveScience.com. Imaginova. Consultado el 11 de diciembre de 2008.
- Rothschild, B.M.; Tanke, D.H., Helbling II, M. and Martin, L.D. (2003). «Epidemiologic study of tumors in dinosaurs». Naturwissenschaften 90 (11): 495-500. doi:10.1007/s00114-003-0473-9. Consultado el 25 de julio de 2008.
- Rothschild, Bruce; Tanke, Darren H. (2007). «Osteochondrosis is Late Cretaceous Hadrosauria». En Carpenter, Kenneth (ed.), ed. Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs. Bloomington and Indianapolis: Indiana University Press. pp. 171-183. ISBN 0-253-34817-X.
- Galton, Peter M. (1973). «The cheeks of ornithischian dinosaurs». Lethaia 6: 67-89. doi:10.1111/j.1502-3931.1973.tb00873.x.
- Fastovsky, D.E., and Smith, J.B. (2004). "Dinosaur paleoecology." The Dinosauria. pp. 614–626.
- Barrett, Paul M. (2005). «The diet of ostrich dinosaurs (Theropoda: Ornithomimosauria)». Palaeontology 48 (2): 347-358. doi:10.1111/j.1475-4983.2005.00448.x.
- Weishampel, David B. (1984). Evolution in jaw mechanics in ornithopod dinosaurs. Advances in Anatomy, Embryology, and Cell Biology 87. Berlin; New York: Springer-Verlag. ISBN 0387131140. ISSN 0301-5556. PMID 6464809.
- Rybczynski, Natalia; Tirabasso, Alex; Bloskie, Paul; Cuthbertson, Robin; Holliday, Casey (2008). «A three-dimensional animation model of Edmontosaurus (Hadrosauridae) for testing chewing hypotheses». Palaeontologia Electronica 11 (2): online publication. Consultado el 10 de agosto de 2008.
- Holliday, Casey M.; Witmer, Lawrence M. (2008). «Cranial kinesis in dinosaurs: intracranial joints, protractor muscles, and their significance for cranial evolution and function in diapsids». Journal of Vertebrate Paleontology 28 (4): 1073-1088.
- Williams, Vincent S.; Barrett, Paul M.; Purnell, Mark A. (2009). «Quantitative analysis of dental microwear in hadrosaurid dinosaurs, and the implications for hypotheses of jaw mechanics and feeding». Proceedings of the National Academy of Sciences 106 (27): 11194-11199. Bibcode:2009PNAS..10611194W. PMC 2708679. PMID 19564603. doi:10.1073/pnas.0812631106.
- Cuthbertson, Robin S.; Tirabasso, Alex; Rybczynski, Natalia; Holmes, Robert B. (2012). «Kinetic limitations of intracranial joints in Brachylophosaurus canadensis and Edmontosaurus regalis (Dinosauria: Hadrosauridae), and their implications for the chewing mechanics of hadrosaurids». The Anatomical Record 295 (6): 968-979. PMID 22488827. doi:10.1002/ar.22458.
- Sternberg, Charles H. (1909). «A new Trachodon from the Laramie Beds of Converse County, Wyoming». Science 29 (749): 753-54. Bibcode:1909Sci....29..747G. doi:10.1126/science.29.749.747.
- Currie, Philip J.; Koppelhus, Eva B.; Muhammad, A. Fazal (1995). «"Stomach" contents of a hadrosaurid from the Dinosaur Park Formation (Campanian, Upper Cretaceous) of Alberta, Canada». En Sun Ailing and Wang Yuangqing (editors), ed. Sixth Symposium on Mesozoic Terrestrial Ecosystems and Biota, Short Papers. Beijing: China Ocean Press. pp. 111-114. ISBN 978-7-5027-3898-3.
- Kräusel, R. (1922). «Die Nahrung von Trachodon». Paläontologische Zeitschrift (en alemán) 4 (2–3): 80. doi:10.1007/BF03041547.
- Abel, O. (1922). «Diskussion zu den Vorträgen R. Kräusel and F. Versluys». Paläontologische Zeitschrift (en alemán) 4 (2–3): 87. doi:10.1007/BF03041547.
- Wieland, G. R. (1925). «Dinosaur feed». Science 61 (1589): 601-603. Bibcode:1925Sci....61..601W. PMID 17792714. doi:10.1126/science.61.1589.601.
- Carpenter, Kenneth (1987). «Paleoecological significance of droughts during the Late Cretaceous of the Western Interior». En Currie, Philip J. and Koster, Emlyn H. (editors), ed. Fourth Symposium on Mesozoic Terrestrial Ecosystems, Drumheller, August 10–14, 1987. Occasional Paper of the Tyrrell Museum of Palaeontology 3. Drumheller, Alberta: Royal Tyrrell Museum of Palaeontology. pp. 42-47. ISBN 978-0-7732-0047-0.
- Carpenter, Kenneth (2007). «How to make a fossil: part 2 – Dinosaur mummies and other soft tissue» (PDF). The Journal of Paleontological Sciences. online. Archivado desde el original el 23 de agosto de 2009. Consultado el 8 de marzo de 2009.
- Tweet, Justin S.; Chin, Karen; Braman, Dennis R.; Murphy, Nate L. (2008). «Probable gut contents within a specimen of Brachylophosaurus canadensis (Dinosauria: Hadrosauridae) from the Upper Cretaceous Judith River Formation of Montana». PALAIOS 23 (9): 624-635. Bibcode:2008Palai..23..624T. doi:10.2110/palo.2007.p07-044r.
- Sellers, W. I.; Manning, P. L.; Lyson, T.; Stevens, K.; Margetts, L. (2009). «Virtual palaeontology: gait reconstruction of extinct vertebrates using high performance computing». Palaeontologia Electronica 12 (3): unpaginated. Consultado el 13 de diciembre de 2009.
- Bakker, Robert T. (1986). «The case of the duckbill's hand». The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction. New York: William Morrow. pp. 146–159. ISBN 978-0-8217-2859-8.
- Brett-Surman, M. K. (1997). «Ornithopods». En James O. Farlow; M. K. Brett-Surman (eds.), eds. The Complete Dinosaur. Bloomington: Indiana University Press. pp. 330–346. ISBN 978-0-253-33349-0.
- Dodson, Peter (1996). The Horned Dinosaurs: A Natural History. Princeton: Princeton University Press. pp. 14–15. ISBN 978-0-691-05900-6.
- Sullivan, Robert M.; Lucas, Spencer G. (2006). «The Kirtlandian land-vertebrate "age" – faunal composition, temporal position and biostratigraphic correlation in the nonmarine Upper Cretaceous of western North America» (pdf). En Lucas, Spencer G.; and Sullivan, Robert M. (eds.), ed. Late Cretaceous Vertebrates from the Western Interior. New Mexico Museum of Natural History and Science Bulletin 35. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 7-29.
- Russell, Dale A. (1989). An Odyssey in Time: Dinosaurs of North America. Minocqua, Wisconsin: NorthWord Press, Inc. pp. 170-171. ISBN 978-1-55971-038-1.
- Bakker, Robert T. (1986). The Dinosaur Heresies. p. 438.
- Lehman, Thomas M. (2001). «Late Cretaceous dinosaur provinciality». En Tanke, Darren; and Carpenter, Kenneth (eds.), ed. Mesozoic Vertebrate Life. Bloomington and Indianapolis: Indiana University Press. pp. 310–328. ISBN 978-0-253-33907-2.
- Russell, Dale A. (1989). An Odyssey in Time: Dinosaurs of North America. pp. 180–181.
- Walking with Dinosaurs: Celluloid (and Digital) Dinosaurs History of Dinosaur Movies (and Television Documentaries). 2000. Retrieved 2007-06-06.
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
- Edmontosaurus en Dinodata (en inglés, se requiere registrarse en forma gratuita)
- Edmontosaurus dentro de Hadrosaurinae en The Thescelsosaurus (en inglés)
- Significado y pronunciación del nombre (en inglés)
- Ficha e imagen de Edmontosaurus en Dinosaurier-web (en alemán)
- Edmontosaurus en The DINOSAUR ENCYCLOPAEDIA Version 4.0 Archivado el 9 de julio de 2008 en Wayback Machine. (en inglés)
| Control de autoridades |
|
|---|
 Datos: Q309466
Datos: Q309466 Multimedia: Edmontosaurus / Q309466
Multimedia: Edmontosaurus / Q309466 Especies: Edmontosaurus
Especies: Edmontosaurus