Triceratops
Triceratops (del griego τρι tri, 'tres', κέρας kéras, 'cuerno' y -ωψ -ops, 'cara'[1]) es un género con dos especies conocidas de dinosaurios ceratopsianos ceratópsidos, que vivieron a finales del período Cretácico, hace aproximadamente entre 68 y 66 millones de años, durante el Maastrichtiense, en lo que hoy es Norteamérica. Es uno de los últimos géneros en aparecer antes del gran evento de extinción masiva del Cretácico-Terciario.[2] Poseía una gran gola ósea que le cubria dorsal y lateral del cuello y tres cuernos, un cuerpo grande sobre cuatro fornidas patas, y varias similitudes con respecto al moderno rinoceronte y los bovinos. Triceratops es uno de los más reconocidos de todos los dinosaurios. También fue uno de los más grandes, de hasta 8 a 9 metros de largo y de 5 a 9 toneladas métricas de masa corporal. Aunque compartiera el territorio con el temible Tyrannosaurus y fuera cazado por él,[3] no está claro si los dos lucharon de la manera representada a menudo en exhibiciones de museo o en imágenes populares.
| Triceratops | ||
|---|---|---|
| Rango temporal: 68 Ma - 66 Ma Maastrichtiense (Cretácico Superior) | ||
 Reconstrucción de un Triceratops. | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Ornithischia | |
| Suborden: | Neornithischia | |
| Infraorden: | Ceratopsia | |
| Familia: | Ceratopsidae | |
| Subfamilia: | Chasmosaurinae | |
| Género: |
Triceratops Marsh, 1889 | |
| Especie tipo | ||
|
Triceratops horridus Marsh, 1889 | ||
| Otras especies | ||
| ||
| Sinonimia | ||
| ||
Aún no se ha encontrado un esqueleto completo de Triceratops,[4] aunque la criatura está bien documentada por numerosos restos parciales recogidos desde la introducción del género en 1887. Se han encontrado especímenes que representan etapas de vida desde la cría hasta el adulto. La función de la gola o volante y de los tres cuernos faciales distintivos ha inspirado largas discusiones. Aunque se vean tradicionalmente como armas defensivas contra depredadores, las últimas teorías proponen que es más probable que estas características fueron utilizadas durante el cortejo sexual y exhibiciones de dominancia, como la cornamenta de los modernos reno, la cabra blanca, o el escarabajo rinoceronte.[5]
Triceratops es el ceratópsido del que los científicos han adquirido más conocimientos, pero la colocación exacta del género dentro del grupo ha sido, sin embargo, un punto de discusión entre los paleontólogos. Tradicionalmente dentro de los Centrosaurinae, "con volantes cortos", pero los estudios cladísticos modernos muestran que es un miembro de Chasmosaurinae que generalmente tiene "volantes largos". Dos especies, Triceratops horridus y Triceratops prorsus, son actualmente consideradas válidas, aunque se han descrito muchas otras. Recientes investigaciones parecen apuntar a que el conocido ceratópsido, Torosaurus, representa una forma completamente madura de Triceratops más que un género aparte.[6] Esta opinión ha sido cuestionada y lentamente se la esta dejando de lado. Como el ceratópsido arquetípico, Triceratops es uno de los dinosaurios más populares y ha aparecido en películas, sellos postales y muchos otros tipos de medios.[7]
Descripción
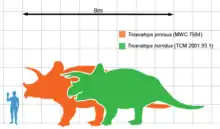
Era un animal muy grande, el tamaño de los individuos de Triceratops se estima entre los 7,8 a 9 metros de largo y 2,9 a 3 de altura[8][9] y 5-9 toneladas de masa corporal.[10][11][12][13][14] Lo más distintivo era su gran cabeza, proporcionalmente una de las más grandes de todos los animales terrestres. Pudo haber medido alrededor de 2 metros de largo,[5] y casi podía alcanzar un tercio de la longitud total del animal.[4] Tenía un único cuerno en el hocico, sobre las narinas, y un par de cuernos de aproximadamente 1 metro de largo, sobre cada ojo. En la parte trasera del cráneo tenía una gola ósea (expansión nucal o plataforma craneal posterior) relativamente corta y sin fenestras, a diferencia de la mayoría de los demás ceratópsidos, que tenían grandes aberturas en esta estructura. Un espécimen de T. horridus llamado Kelsey medía de 6,7 a 7,3 metros de largo con un cráneo de 2 metros, medía unos 2,3 metros de altura, y el Black Hills Institute estimó que pesaba aproximadamente 5,4 toneladas.[15][16]
Cráneo
Como todos los casmosaurinos, Triceratops tenía un cráneo grande en relación con el tamaño de su cuerpo, entre los más grandes de todos los animales terrestres. Se estima que el cráneo más grande conocido, el espécimen MWC 7584, anteriormente BYU 12183, tenía 2,5 metros de largo cuando estaba completo,[17] y podría alcanzar casi un tercio de la longitud del animal completo.[4]

La parte delantera de la cabeza estaba equipada con un gran pico delante de los dientes. El núcleo del pico superior estaba formado por un hueso rostral especial. Detrás de él, se ubicaron los huesos premaxilares, embalados por detrás por orificios nasales circulares muy grandes. En los casmosaurinos, los premaxilares se reunían en su línea media en una placa ósea compleja, cuyo borde posterior estaba reforzado por el "puntal narial". Desde la base de este puntal, un proceso triangular sobresalía hacia la fosa nasal. Triceratops se diferencia de la mayoría de los parientes en que este proceso fue ahuecado en el lado exterior. Detrás del premaxilar desdentado, el maxilar tenía de treinta y seis a cuarenta posiciones de dientes, en las que se apilaban verticalmente de tres a cinco dientes por posición. Los dientes estaban estrechamente adheridos, formando una "batería dental" curvada hacia el interior. El rostro tenía un solo cuerno en el hocico, por encima de las fosas nasales. En Triceratops, el cuerno de la nariz a veces se reconoce como una osificación separada, la epinasal.[18]
El cráneo también presentaba un par de "cuernos de las cejas" o cuernos supraorbitales de aproximadamente 1 metro de largo, uno encima de cada ojo.[19][20] Los huesos yugulares apuntaban hacia abajo en los lados posteriores del cráneo y estaban cubiertos por epijugales separados. En Triceratops, estos no eran particularmente grandes y, a veces, tocaban los quadratojugales. Los huesos del techo del cráneo estaban fusionados. Mediante un plegamiento de los huesos frontales, se creó un techo de cráneo "doble". En Triceratops, algunos ejemplares muestran una fontanela, una abertura en la capa superior del techo. La cavidad entre las capas invadió los núcleos óseos de los cuernos de las cejas.[18]

En la parte posterior del cráneo, los huesos escamosos externos y los huesos parietales internos se convirtieron en un volante óseo relativamente corto, adornado con epoccipitales en especímenes jóvenes. Estos eran procesos triangulares bajos en el borde del volante, que representaban osificaciones de piel separadas u osteodermos. Por lo general, con especímenes de Triceratops hay dos epoccipitales presentes en cada hueso parietal, con un proceso central adicional en su borde. Cada hueso escamoso tenía cinco procesos. La mayoría de los otros ceratópsidos tenían grandes fenestras parietales, aberturas, en sus volantes, pero los de Triceratops eran notablemente sólidos,[21] a menos que el género Torosaurus representa individuos maduros de Triceratops. Debajo del volante, en la parte posterior del cráneo, un enorme cóndilo occipital, de hasta 106 milímetros de diámetro, conectaba la cabeza con el cuello.[18]
Las mandíbulas inferiores eran alargadas y se unían en sus puntas en un hueso epidentario compartido, el núcleo del pico inferior desdentado. En el hueso dentario, la batería de dientes se curvaba hacia el exterior para encontrarse con la batería del maxilar superior. En la parte posterior de la mandíbula inferior, el hueso articular era excepcionalmente ancho, coincidiendo con el ancho general de la articulación de la mandíbula.[18] T. horridus se puede distinguir de T. prorsus por tener un hocico menos profundo.[12]
Gola o volante óseo
Desde su nuca y extendiéndose sobre el cuello, Triceratops poseía una gola ósea, la plataforma craneal posterior, sobre el que se extendía una capa de piel, profusamente irrigada por numerosos vasos sanguíneos, de lo que se tiene cuenta dada las improntas que estos capilares dejaron en el hueso. Erróneamente se pensó en la antigüedad que esta gola cumplía una función defensiva para el animal, lo que ha sido descartado recientemente dada su fragilidad, pues no constituía una placa ósea lo suficientemente sólida y completa. Algunos paleontólogos incluso han llegado a sugerir que una lesión en dicha gola puede haber resultado en extremo nociva para el animal, dada su alta irrigación sanguínea. Entre las numerosas funciones que se han atribuido a esta formación ósea, se encuentran las de termorregulación y de exhibición tanto sexual como defensiva, fundadas en la circulación sanguínea del animal. En el primer caso, actuando como una vela similar al caso del Spinosaurus, y en las segundas alternativas, debido al enrojecimiento que supuestamente habría adoptado la gola al aumentar el flujo sanguíneo, como en las placas del Stegosaurus. La piel del animal era gruesa y presentaba unas pequeñas protuberancias que se diseminaban por su cuerpo de manera irregular.
Pico
La boca recuerda a un pico de loro, mientras que la dentadura revela que la alimentación de los Triceratops era eminentemente herbívora. Tras arrancar los brotes y hojas trituraba el alimento con hileras de dientes localizados en el fondo de la boca que le servían como un mecanismo para esquilar el alimento. Esas piezas dentales crecían de nuevo una vez se desgastaban.[22] Ya que la posición del cráneo es bastante baja, lo más probable es que su comportamiento alimentario fuese eminentemente ramoneador. Los dientes de Triceratops son uno de los fósiles más abundantes del final del Cretácico en el norte de Estados Unidos, sugiriendo que fueron los herbívoros dominantes de su tiempo
Los tres cuernos
El cráneo distintivo de Triceratops tenía un solo cuerno nasal, corto y grueso, localizado sobre las ventanas de la nariz, además de un par de cuernos de aproximadamente 1 metro de longitud, los cuales, estaban ubicados sobre las cuencas oculares. Las crías tenían los cuernos rectos, con una longitud de 5-12 centímetros,[23] antes de que maduren y comience a formarse una curvatura. La parte posterior del cráneo posee una pequeña concavidad ósea. Se han propuesto un diverso número de teorías sobre el uso de los cuernos que todavía están en discusión, aunque estas no son mutuamente excluyentes.
- Como defensa contra los terópodos.
- Como medio de comunicación entre miembros de su manada.
- Como medio de disputa de territorios y apareamiento.
- Como medio de cortejo.
- Como un símbolo de estatus que reflejaba o determinaba su posición en la manada.
- Como puntos de anclaje para los músculos de la mandíbula.
- Como un medio de termorregulación.
Esqueleto axial
Los casmosaurinos mostraron poca variación en su esqueleto poscraneal.[18] El esqueleto de Triceratops es notablemente robusto. Ambas especies de Triceratops poseían una constitución robusta, con extremidades fuertes, manos cortas con tres pezuñas cada una y pies cortos con cuatro pezuñas cada uno.[24] La columna vertebral constaba de 10 vértebras del cuello, 10 de la espalda, 10 sacras y unas 45 de la cola. Las vértebras frontales del cuello se fusionaron en una sincervical. Tradicionalmente, se suponía que esto había incorporado las tres primeras vértebras, lo que implicaría que el atlas más frontal era muy grande y lucía una columna neural. Interpretaciones posteriores revivieron una vieja hipótesis de John Bell Hatcher que muy al frente se puede observar un vestigio del atlas real, el sincervical entonces compuesto por cuatro vértebras. El conteo vertebral mencionado, se ajusta a esta visión. En Triceratops, las espinas neurales del cuello tienen una altura constante, en lugar de inclinarse gradualmente hacia arriba. Otra peculiaridad es que las costillas del cuello recién empiezan a alargarse con la novena vértebra cervical.[18]
Las vértebras más bien cortas y altas de la espalda estaban reforzadas en su región media por tendones osificados que discurrían a lo largo de la parte superior de los arcos neurales. El sacro recto era largo y los individuos adultos muestran una fusión de todas las vértebras sacras. En Triceratops, los primeros cuatro y los dos últimos sacros tenían procesos transversales, que conectaban la columna vertebral con la pelvis, que estaban fusionados en sus extremos distales. Los sacros 7 y 8 tenían procesos más largos, lo que hacía que el sacro tuviera un perfil ovalado en la vista superior. En la parte superior del sacro estaba presente una placa neural formada por una fusión de las espinas neurales de la segunda a la quinta vértebra.[18]
Extremidades
Como se dijo anteriormente Triceratops tenía un cuerpo robusto y poderoso, apoyado sobre sus cuatro extremidades que eran muy fuertes y con los dedos de las extremidades anteriores con cinco dedos y cuatro en las posteriores, todos formando un fuerte casco.[18] Se movían sobre sus cuatro gruesas patas; las delanteras destacaban por su fuerza, ya que debían soportar el peso de un cráneo de dimensiones desproporcionadas. Aunque ciertamente cuadrúpedo, la postura de estos dinosaurios ha sido objeto de largo debate. Originalmente se creyó que las extremidades anteriores del animal tuvieron que estar dispuestas en ángulos desde el tórax para soportar mejor el peso de la cabeza.[5] Esta posición se puede ver en las clásicas pinturas de Charles Knight y Rudolph Zallinger. Sin embargo, las icnitas (huellas de pisadas) atribuidas a Triceratops y las reconstrucciones recientes de esqueletos tanto reales como digitales, parecen demostrar que Triceratops mantuvo una postura vertical durante la locomoción normal, con los codos levemente arqueados hacia fuera, en una posición intermedia entre completamente vertical y completamente arqueado, como en el rinoceronte moderno.[25][26] Esta conclusión no imposibilita un paso arqueado para las confrontaciones o la alimentación.

Las manos y los antebrazos del Triceratops conservaron una estructura bastante primitiva en comparación con otros dinosaurios cuadrúpedos como los tireóforos y muchos saurópodos. En esos dos grupos, las extremidades anteriores de las especies cuadrúpedas generalmente se rotaban de modo que las manos miraban hacia adelante con las palmas hacia atrás, "pronadas", mientras los animales caminaban. Triceratops, al igual que otros ceratopsianos y los ornitópodos cuadrúpedos relacionados, que juntos forman Cerapoda, caminaban con la mayoría de sus dedos apuntando hacia afuera del cuerpo, la condición original de los dinosaurios, también retenida por formas bípedas como los terópodos. En Triceratops, el peso del cuerpo lo soportaban solo los primeros tres dedos de la mano, mientras que los dígitos 4 y 5 eran vestigiales y carecían de garras o pezuñas.[24] La fórmula de la falange de la mano es 2-3-4-3-1, lo que significa que el primer dedo o el más interno de la extremidad anterior tiene dos huesos, el siguiente tiene tres, así sucesivamente.[27] Triceratops tenía una pelvis grande con un ilion largo y el isquion estaba curvado hacia abajo. El pie era corto con cuatro dedos funcionales. La fórmula de la falange del pie es 2-3-4-5-0.[18]
Piel
Se conoce piel preservada de Triceratops a partir de un espécimen encontrado en Wyoming, aunque no se ha publicado formalmente en la literatura científica. Estos consisten en tubérculos hexagonales grandes de entre 50 a 60 milímetros de ancho, junto con tubérculos más grandes de alrededor de 100 milímetros de ancho con proyecciones cónicas.[28]
Descubrimiento e investigación
El primer espécimen nombrado hoy considerado un Triceratops es un par de cuernos orbitales unidos a un pedazo del techo del cráneo, encontrado cerca de Denver, Colorado en la primavera de 1887.[29] Este epécimen fue enviado a Othniel Charles Marsh, quien creyó que provenía de rocas de una formación datada en el Plioceno, y que los huesos pertenecían a un gran e inusual bisonte, al que denominó Bison alticornis.[29][30] El propio Marsh fue el primero en describir los dinosaurios con cuernos, un año después al presentar al Ceratops a partir de restos fragmentarios,[31] pero siguió pensando que B. alticornis era un mamífero del Plioceno. Hizo falta un tercer espécimen para que cambiara de posición.

Ese espécimen, el holotipo de Triceratops YPM 1820, obtenido en 1888 por John Bell Hatcher en la Formación Lance de Wyoming, fue inicialmente descrito como otra especie de Ceratops.[32] El vaquero Edmund B. Wilson se había sobresaltado al ver una calavera monstruosa que sobresalía del costado de un barranco. Intentó recuperarlo tirando un lazo alrededor de uno de los cuernos. Cuando se rompió, y el cráneo cayó al fondo de la hendidura, Wilson le llevó el cuerno a su jefe, el ranchero y ávido coleccionista de fósiles Charles Arthur Guernsey, quien casualmente se lo mostró a Hatcher. Posteriormente, Marsh ordenó a Hatcher que localizara y salvara el cráneo.[5] El holotipo primero se llamó Ceratops horridus, cuando durante la preparación posterior descubrió el tercer cuerno de la nariz, Marsh cambió de opinión y le dio a la pieza el nombre genérico Triceratops, literalmente, "cara de tres cuernos", aceptando a su Bison alticornis como otra especie de Ceratops,[33] que más tarde sería incluida en Triceratops.[34] La robusta naturaleza del cráneo de estos animales ha permitido que muchos ejemplares se hayan preservado como fósiles, lo que también ha ayudado a que puedan ser estudiadas variaciones entre diversas especies e individuos. Han sido encontrado restos de Triceratops en los estados de EE. UU. de Montana, Dakota del Sur, Colorado y Wyoming, y en las provincias canadienses de Saskatchewan y Alberta. Un espécimen anterior, también recuperado de la Formación Lance, fue nombrado Agathaumas sylvestris por Edward Drinker Cope en 1872. Originalmente identificado como un hadrosáurido, este espécimen consiste solo en restos poscraneales y solo se considera provisionalmente un ejemplo de Triceratops.[35]
Aunque no se pueden asignar de forma definitiva, los fósiles que posiblemente pertenecen a Triceratops fueron descritos como dos taxones, Agathaumas sylvestris y Polyonax mortuarius en 1872 y 1874 respectivamente, por el rival de Marsh, Edward Drinker Cope.[36][37] Agathaumas fue nombrado con base en una pelvis, varias vértebras y algunas costillas recolectadas por Fielding Bradford Meek y Henry Martyn Bannister cerca del río Green del sureste de Wyoming a partir de capas provenientes de la Formación Lance de Maastrichtiense.[38] Debido a la naturaleza fragmentaria de los restos, solo se puede asignar con seguridad a Ceratopsidae.[5][39] Polyonax mortuarius fue recolectado por el mismo Cope en 1873 del noreste de Colorado, posiblemente proveniente de la Formación Denver del Maastrichtiense.[40][37] Los fósiles solo consistían en núcleos fragmentarios de cuernos, 3 vértebras dorsales y elementos fragmentarios de extremidades.[37] Polyonax tiene el mismo problema que Agathaumas, con los restos fragmentarios no asignables más allá de Ceratopsidae.[41][5]
Especies
A lo largo de su historia taxonómica una gran cantidad de especies fueron asignadas al género. Hoy en día solo se reconocen dos de todas aquellas, Triceratops horridus, la especie tipo descrita por Marsh en 1889 y Triceratops prorsus también erigida por Marsh un año después en 1890.[42]
Después de que se describiera Triceratops, entre 1889 y 1891 Hatcher recolectó con gran esfuerzo otros treinta y uno de sus cráneos, que variaban en menor o mayor grado del Triceratops original, llamado T. horridus por Marsh. Este nombre específico proviene del latín horridus, rugoso, refiriéndose a la textura rugosa de aquellos huesos pertenecientes al espécimen holotipo, más tarde identificado como un individuo anciano. Esta variación no es de asombrar, dado que los cráneos de Triceratops son objetos tridimensionales grandes, que proceden de individuos de diversas edades y de ambos sexos, y que fueron sujetos a presiones de intensidad y dirección variadas durante la fosilización.[5] Los investigadores las describirían como especies distintas, como se ve más abajo, y establecieron varias filogenenias de cómo se relacionarían unas con otras.

En la primera tentativa de entender las muchas especies, Lull diferenció dos grupos, aunque no describió los caracteres en que se basó. Uno compuesto por T. horridus, T. prorsus y Triceratops brevicornus, y el otro por Triceratops elatus y Triceratops calicornis. Por otro lado dejó dos especies, Triceratops serratus y Triceratops flabellatus aparte de estos grupos.[34] En 1933, y en su revisión de la monografía fundamental de Hatcher, Marsh y Lull de 1907 de todos los ceratopsianos conocidos, conservó sus dos grupos y las dos especies no afiliadas, más un tercer linaje formado por Triceratops obtusus y Triceratops hatcheri, caracterizados por un cuerno nasal muy pequeño.[39] El linaje T. horridus-T. prorsus-T. brevicornus, probablemente el linaje más conservador, muestra un aumento de tamaño del cráneo y una disminución del tamaño del cuerno nasal, y el linaje T. elatus-T. calicornis fue definido por sus largos cuernos orbitales y pequeño cuerno nasal.[39] C. M. Sternberg hizo una modificación al agregar Triceratops eurycephalus y sugiriendo que ligara los segundos y tercer linajes más juntos que estos con el linaje de T. horridus.[43] Este patrón fue seguido hasta los grandes estudios de las décadas de 1980 y 1990.

Con tiempo, sin embargo, ganó apoyo la idea de que los variados cráneos pudieran ser representantes de la variación individual de solo una o dos especies. En 1986, Ostrom y Wellnhofer publicaron un trabajo en el cual propusieron que hubiera solamente una especie, Triceratops horridus.[44] La argumentación de su análisis era que generalmente hay solamente una o dos especies de cualquier animal grande en una región, como los modernos elefante y jirafa en el África de hoy. A sus resultados, Lehman agregó los viejos linajes del período de Lull y Sternberg combinados con madurez y dimorfismo sexual, sugiriendo que el supuesto T. horridus, T. prorsus y T. brevicornus estaba compuesto por hembras, el supuesto linaje T. calicornis y T. elatus por machos, y el de T. obtusus y T. hatcheri por viejos machos enfermos.[45] Su razonamiento era que los machos tenían cuernos más altos y erguidos y cráneos más grandes, y las hembras tenían cráneos más pequeños con cuernos más cortos, dirigidos hacia adelante.
Estos resultados, sin embargo, fueron discutidos algunos años más tarde por Catherine Forster, quien reanalizó el material de Triceratops más exhaustivamente y concluyó que los restos encajaban en dos especies, T. horridus y T. prorsus, aunque el cráneo distintivo de T. hatcheri se diferenciaba lo suficiente como para justificar un género separado, hoy Nedoceratops.[46] Esta autora encontró que T. horridus se agrupaba con otras especies, mientras que T. prorsus y T. brevicornus quedaban aparte, y puesto que había muchos más especímenes en el primer grupo, sugirió que esto significaba que los dos grupos representaban dos especies. Sin embargo, es todavía posible interpretar las diferencias de los dos grupos como la representación de una sola especie con dimorfismo sexual.[5][47] Scannella y Fowler en 2009 también apoyaron la separación de T. prorsus y T. horridus, y notaron que las dos especies estaban separadas estratigráficamente dentro de la Formación Hell Creek, indicando que nunca vivieron juntas al mismo tiempo.[48] En 2009, John Scannella agregó una nueva dimensión a la discusión, con la posible reinterpretación de Torosaurus, como ejemplares machos maduros de Triceratops, cosa que se tratara más adelante.[6]
Especies válidas

- Triceratops horridus (Marsh, 1889) (originalmente Ceratops) (especie tipo)
- Triceratops prorsus (Marsh, 1890)
Especies dudosas
Las siguientes son especies consideradas dudosas, y están basadas en restos muy incompletos o pobres para asignarlas a una especie conocida de Triceratops, no pudiendo ser clasificadas masallá de Triceratops sp..
Errores de asignación
Las siguientes especies son sinónimos, como se indica entre paréntesis ("= T. horridus " o "= T. prorsus ").
- Triceratops albertensis (C. M. Sternberg, 1949) (=T. horridus)
- Triceratops brevicornus (Hatcher, 1905) (=T. horridus)
- Triceratops calicornis (Marsh, 1898) (=T. horridus)
- Triceratops elatus (Marsh, 1891) (=T. horridus)
- Triceratops eurycephalus (Schlaikjer, 1935) (=T. horridus)
- Triceratops flabellatus (Marsh, 1889) (=T. horridus)
- Triceratops hatcheri (Lull, 1907) (=Nedoceratops hatcheri)
- Triceratops "ingens" (Marsh vide Lull, 1915) (=T. horridus)
- Triceratops mortuarius (Cope, 1874) (nomen dubium; originalmente Polyonax; =Polyonax mortuarius)
- Triceratops obtusus (Marsh, 1898) (=T. horridus)
- Triceratops serratus (Marsh, 1890) (=T. horridus)
- Triceratops sylvestris (Cope, 1872) (nomen dubium; originalmente Agathaumas sylvestris)
Controversia conTorosaurus

Torosaurus es un ceratópsido que fue identificado dos años después de Triceratops en 1891, a partir de dos cráneos. El género Torosaurus coincide con Triceratops en edad geológica, distribución, anatomía y tamaño, siendo reconocido como su pariente más cercano.[49] Se diferencia por un cráneo más largo con dos grandes ventanas en su gola. Paleontólogos investigadores de la ontogenia de los dinosaurios de la Formación Hell Creek de Montana, han presentado pruebas de que ambos son en realidad un solo género.
John Scannella, en su trabajo presentado en Bristol, Reino Unido a la conferencia de la Sociedad para la Paleontología de Vertebrados (25 de septiembre de 2009) reclasificó especímenes de Torosaurus como ejemplares completamente desarrollados de Triceratops, quizás de un solo sexo. Jack Horner, el mentor de Scannella en la Universidad de Montana, observó que los cráneos de ceratopsianos consisten de hueso metaplástico. Una característica del hueso metaplástico es que se alarga y se acorta en el tiempo, extendiéndose y reabsorbiéndose, adquiriendo nuevas formas. Horner observó una variedad significativa de formas incluso en esos cráneos identificados ya como Triceratops, «en los que los cuernos se orientan hacia atrás en los jóvenes y hacia delante en los adultos». Aproximadamente el 50 % de todos los cráneos de subadultos de Triceratops tienen dos áreas de hueso más fino en la plataforma craneal posterior, que se corresponden con la ubicación de las fenestras en las plataformas de los cráneos de Torosaurus, indicando que las aberturas se desarrollaron para compensar el peso que habría ido apareciendo con la maduración de los individuos de Triceratops al desarrollar volantes más largos.[6][50] Scannella y Horner publicaron en julio de 2010 un artículo que describe estos hallazgos en detalle. Argumenta formalmente que Torosaurus y el contemporáneo Nedoceratops son sinónimos de Triceratops.[17]
Esta idea fue discutida en 2011 por el especialista en ceratopsios Andrew Farke, quien publicó una redescripción de Nedoceratops hatcheri, una especie problemática que en diferentes ocasiones se ha considerado como representante de su propio género, como un sinónimo de alguna especie de Triceratops o bien una especie distinta del mismo, o, según la hipótesis de Scannella y Horner, un ejemplo de un estadio de crecimiento intermedio entre las formas conocidas como Triceratops y Torosaurus. Farke concluyó que Nedoceratops hatcheri corresponde a individuos adultos de su propio género, relacionado estrechamente con Triceratops. Este autor también concluyó que los cambios requeridos para «convertir» un Triceratops en un Torosaurus serían un proceso sin precedente entre los ceratópsidos, pues requeriría la adición de huesos epoccipitales, la reversión de la textura del hueso de adulto a un estado inmaduro y de vuelta al adulto, además de un tardío desarrollo de las aberturas en la gola.[51]
En un artículo ulterior, publicado en 2011, el paleontólogo Nicholas Longrich afirmó que la sinonimización de Triceratops y Torosaurus no podía ser apoyada sin formas intermedias mejores que las provistas por Scannella y Horner. Sin embargo, Longrich estuvo de acuerdo en que Nedoceratops es un sinónimo de Triceratops[52]
La afirmación encendió el debate. Andrew Farke había subrayado en 2006 que, aparte del volante, no se podían encontrar diferencias sistemáticas entre Torosaurus y Triceratops.[49] Sin embargo, cuestionó la conclusión de Scannella al argumentar en 2011 que los cambios morfológicos propuestos necesarios para "envejecer" un Triceratops en un Torosaurus no tendrían precedentes entre los ceratópsidos. Dichos cambios incluirían el crecimiento de epoccipitales adicionales, la reversión de la textura ósea de un adulto a un tipo inmaduro y nuevamente a un adulto, y el crecimiento de orificios de volantes en una etapa posterior a la habitual.[51] Un estudio de Nicholas Longrich y Daniel Field analizó 35 especímenes de ambos, Triceratops y Torosaurus. Los autores concluyeron que los individuos de Triceratops demasiado viejos para ser considerados formas inmaduras están representados en el registro fósil, al igual que los individuos de Torosaurus demasiado jóvenes para ser considerados adultos completamente maduros. La sinonimia de Triceratops y Torosaurus no puede sustentarse, dijeron, sin formas intermedias más convincentes que las que inicialmente produjeron Scannella y Horner. El espécimen de Triceratops de Scannella con un agujero en su volante, argumentaron, podría representar a un individuo enfermo o malformado en lugar de una etapa de transición entre un Triceratops inmaduro y una forma de Torosaurus maduro.[53][54]
Sinonimia con Nedoceratops, Ojoceratops y Tatankaceratops
En su artículo de 2011, Longrich sugirió que el recientemente descrito Ojoceratops era indistinguible de los especímenes de T. horridus previamente atribuidos a la especie obsoleta T. serratus. Longrich señaló asimismo que otro nuevo género, Tatankaceratops, poseía una extraña mezcla de características de los ejemplares juveniles y adultos de Triceratops, indicando que podría representar una especie enana de Triceratops o simplemente un ejemplar de Triceratops con un desorden en su desarrollo que detuvo su crecimiento prematuramente.[52]
La opinión ha variado sobre la validez de un género separado para Nedoceratops. Scannella y Horner lo consideraron como una etapa de crecimiento intermedia entre Triceratops y Torosaurus.[17][55] Farke, en su redescripción de 2011 del único cráneo conocido, concluyó que se trataba de un individuo anciano de su propio taxón válido, Nedoceratops hatcheri.[51] Longrich y Fields tampoco lo consideraron una transición entre Torosaurus y Triceratops, sugiriendo que los agujeros de los volantes eran patológicos.[54]
Como se describió anteriormente, Scannella había argumentado en 2010 que Nedoceratops debería considerarse un sinónimo de Triceratops.[17] Farke en 2011 sostuvo que representa un género distinto válido.[51] Longrich estuvo de acuerdo con Scannella sobre Nedoceratops e hizo una sugerencia adicional, que Ojoceratops recientemente descrito también era un sinónimo. Los fósiles, argumentó, son indistinguibles de los especímenes de T. horridus que anteriormente se atribuían a la especie desaparecida T. serratus.
Longrich observó que otro género recientemente descrito, Tatankaceratops, mostraba una extraña mezcla de características que ya se encuentran en Triceratops adultos y juveniles. En lugar de representar un género distinto, Tatankaceratops podría representar fácilmente un Triceratops enano o un individuo Triceratops con un trastorno del desarrollo que hizo que dejara de crecer prematuramente.[52]
Clasificación

Triceratops es el dinosaurio de la familia Ceratopsidae del que se tiene más conocimientos, una familia dentro de Ceratopsia, los dinosaurios con cuernos, en su mayoría norteamericanos. La localización exacta de Triceratops entre los ceratopsianos se ha discutido durante muchos años. La confusión provenía principalmente de la combinación de golas cortas y sólidas, similares a los de Centrosaurinae, y de los cuernos largos de la frente, más relacionados con los Chasmosaurinae, antes llamados Ceratopsinae. En la primera descripción de dinosaurios con cuernos, R. S. Lull propuso dos linajes, uno para Monoclonius y Centrosaurus incluyendo a Triceratops, con el otro incluyendo a Ceratops y Torosaurus, haciendo a Triceratops un centrosaurino tal como se entiende hoy al grupo.[34] Revisiones posteriores apoyaron esta visión, describieron formalmente los primeros, el grupo de golas cortas como Centrosaurinae, incluyendo Triceratops, y el segundo, el de las golas largas como Chasmosaurinae.[56][39] En 1949, C. M. Sternberg fue el primero que cuestionó esta posición y propuso que Triceratops estaba más relacionado con Arrhinoceratops y Chasmosaurus de acuerdo con características del cráneo y cuernos, haciendo a Triceratops un ceratopsino según su antigua denominación, hoy un casmosaurino.[43] Sin embargo, esto fue ignorado por John Ostrom,[57] posteriormente por David Norman, ambos incluyendo a Triceratops dentro de Centrosaurinae.[58]

Posteriores descubrimientos y estudios apoyaron la visión de Sternberg sobre la posición de Triceratops, con Lehman definiendo las subfamilias en 1990 y diagnosticando a Triceratops como ceratopsino, casmosaurino según el uso actual, en base de varias características morfológicas. De hecho, entra bien en la subfamilia del Ceratopsinae, hoy Chasmosaurinae, aparte de su una característica, la de una gola corta.[45] Posteriores estudios de Peter Dodson, incluyen un análisis cladistico de 1990[59] y un estudio de 1993 usando RFTRA, análisis de resistencia de rho y theta ajustado,[60] una técnicas morfometricas qué mide sistemáticamente semejanzas en la forma del cráneo, refuerza la posición de Triceratops en la subfamilia Chasmosaurinae.
Los orígenes evolutivos de Triceratops fueron desconocidos durante muchos años después de su descubrimiento. En 1922, Henry Fairfield Osborn interpretó al recientemente descubierto Protoceratops como su antepasado,[61] pero pasaron muchas décadas antes de que nueva información saliera a luz. Los últimos años el siglo XX y primeros del XXI han sido fructíferos en el descubrimiento de varios dinosaurios relacionados con los antepasados de Triceratops. Zuniceratops, el ceratopsiano más antiguo conocido con cuernos en la frente, fue descrito a finales de la década de 1990, y Yinlong, el primer ceratopsiano del Jurásico, en 2005. Estos nuevos hallazgos han sido cruciales en el entendimiento de los orígenes de los dinosaurios con cuernos, sugiriendo un origen en Asia, durante el Jurásico, y la aparición de los verdaderos dinosaurios con cuernos, los ceratópsidos, a principios del Cretácico Superior en Norteamérica.[18] Como Triceratops se ve cada vez más como un miembro de la subfamilia de gola larga, Ceratopsinae, un probable antepasado pudo haberse parecido a Chasmosaurus, que prosperó unos 5 millones de años antes.
Filogenia
En la sistemática filogenética, el género se ha utilizado como punto de referencia en la definición de Dinosaurio. Los dinosaurios se han definido como todos los descendientes del antepasado común más reciente de Triceratops y Neornithes, es decir los pájaros modernos.[62] Además los dinosaurios con cadera de aves, Ornithischia, han sido definido como todos los dinosaurios con un antepasado común más reciente al Triceratops que con las aves modernas.[63] El siguiente cladograma muestra la posición de según Sampson et al., en 2010.[64]

| Ceratopsidae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
El cladograma a continuación sigue a Longrich de 2014, donde se nombró una nueva especie de Pentaceratops e incluyó casi todas las especies de casmosaurinos.[65]
| Chasmosaurinae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiología

Aunque comúnmente se retrate a los Triceratops en manadas, como animales gregarios, realmente hay pocas pruebas. Mientras que se conocen de yacimientos que contienen restos desde dos a centenares o miles de individuos de varios géneros de dinosaurios con cuernos, hasta la fecha hay solamente un yacimiento documentado con varios ejemplares de Triceratops: un lugar en el sudeste de Montana con restos de tres jóvenes. Puede ser significativo que solamente se encontraran restos de individuos juveniles. [66]
En 2012, cerca de Newcastle, Wyoming, se encontró un grupo de tres Triceratops en condiciones relativamente completas, cada uno de diferentes tamaños, desde un adulto hasta un pequeño juvenil. Los restos están siendo excavados actualmente por el paleontólogo Peter Larson y un equipo del Instituto Black Hills. Se cree que los animales viajaban como una unidad familiar, pero se desconoce si el grupo está formado por una pareja apareada y sus crías, o dos hembras y un joven al que cuidan. Los restos también muestran signos de depredación o carroña por parte de Tyrannosaurus, particularmente en el espécimen más grande, con los huesos de las extremidades delanteras que muestran roturas y heridas punzantes de los dientes del depredador.[67] En 2020, Illies y Fowler describieron las vértebras caudales distales coosificadas de Triceratops. Según ellos, esta patología podría haber surgido después de un Triceratops accidentalmente pisara la cola de otro miembro de la manada.[68][69]

Por muchos años solo se encontraron individuos aislados de Triceratops.[66] Sin embargo, estos restos son muy comunes, por ejemplo, Bruce Erickson, un paleontólogo del Museo de Ciencias de Minnesota, ha reportado que se conocen 200 especímenes de T. prorsus en la Formación Hell Creek de Montana.[70] Igualmente, Barnum Brown comentó haber visto más de 500 cráneos en el campo.[71] Debido a que los dientes, fragmentos de los cuernos, de la gola, y otros fragmentos del cráneo de Triceratops son los fósiles más abundantes en la Formación Lance, de finales del Maastrichtiense durante el Cretácico Superior, hace 68 a 65 millones de años en Norteamérica occidental, Triceratops está considerado como uno de los herbívoros dominantes de su tiempo, si no el más dominante. En 1986, Robert Bakker estimó que componía 5/6 de la fauna de grandes dinosaurios a finales del Cretácico.[72] A diferencia de lo que sucede con otros animales, los restos del cráneo son más comunes que los postcraneales, lo que sugiriere que el cráneo tuviera un potencial inusualmente alto de preservación.[73] Triceratops fue uno de los últimos géneros de ceratópsidos en aparecer antes de la extinción masiva del Cretácico-Terciario. Los relacionados Nedoceratops y Torosaurus, y el más distante y diminuto Leptoceratops, estaban también presentes, aunque sus restos son escasos.[5]
El análisis de la anatomía endocraneal de Triceratops sugiere que su sentido del olfato era pobre en comparación con el de otros dinosaurios. Sus oídos estaban en sintonía con los sonidos de baja frecuencia dadas las longitudes cocleares cortas registradas en un análisis realizado por Sakagami et al. Este mismo estudio también sugiere que Triceratops mantuvo su cabeza a unos 45 grados con respecto del suelo, un ángulo que exhibiría los cuernos y el volante de la manera más efectiva y al mismo tiempo permitiría que el animal aprovechara la comida a través del pastoreo.[74]
Un estudio de 2022 realizado por Wiemann y sus colegas de varios géneros de dinosaurios, incluido el Triceratops, sugiere que tenía un metabolismo ectotérmico, de sangre fría, o gigantotérmico, a la par con el de los reptiles modernos. Esto se descubrió utilizando la espectroscopia de las señales de lipoxidación, que son subproductos de la fosforilación oxidativa y se correlacionan con las tasas metabólicas. Sugirieron que tales metabolismos pueden haber sido comunes para los dinosaurios ornitisquios en general y que el grupo evolucionó hacia la ectotermia a partir de un antepasado con un metabolismo endotérmico, de sangre caliente.[75]
Dentición y dieta

Los Triceratops eran herbívoros y, debido a su cabeza baja, su alimento principal era probablemente plantas de crecimiento bajo, aunque pudieron haber derribado árboles más altos con sus cuernos, pico, y cuerpo.[18][76] Las mandíbulas terminaban en un pico profundo y estrecho, del que se piensa que estaba más adaptado para arrancar y ramonear que para morder.[77]
Los dientes del Triceratops estaban agrupados en baterías, de 36 a 40 hileras de dientes a cada lado de cada mandíbula, con tres a cinco dientes apilados por hileras, dependiendo del tamaño del animal.[18] Esto le da un total de 432 a 800 dientes, de los cuales solamente una fracción estaba funcionando en un momento dado; el reemplazo de los dientes era continuo y ocurría a lo largo de la vida del animal.[18] Estos actuaban cortando en una posición vertical o casi vertical[18] El gran tamaño y número de los dientes de un Triceratops sugiere que comían volúmenes grandes de material vegetal fibroso,[18] por lo que se ha sugerido que palmeras y cycas formaban la mayor parte de su dieta,[78][79] o helechos, que crecerían en praderas.[80]
Funciones de los cuernos y del volante o gola ósea
Ha habido mucha especulación sobre las funciones de las características principales de Triceratops. Las dos teorías principales han girado alrededor de uso en combate, o exhibición en el cortejo, con esta última idea considerada como la función primaria más probable.[18] En los primeros estudios sobre el género, Lull propuso que la gola o volante craneal posterior pudiera servir de anclaje a los músculos de la masticación, permitiéndole una mayor potencia muscular.[81] Esto fue aceptado por otros autores durante muchos años, pero estudios posteriores no encontraron pruebas de grandes zonas de inserción muscular en los huesos de la gola.[82]
Combate contra depredadores

Otra importante corriente de pensamiento vio la gola y los cuernos como armas en la lucha contra depredadores como Tyrannosaurus, esta idea fue propuesta por primera vez por C. H. Sternberg en 1917 y 70 años después por Robert Bakker.[83][84] Hay pruebas de que un Tyrannosaurus rex ha tenido un encuentro agresivo contra un Triceratops, basándose en las marcas de dientes parcialmente cicatrizadas en un cuerno supraorbital y hueso escamoso de un Triceratops. El cuerno mordido está también quebrado, con nuevo crecimiento óseo después de la rotura. Qué animal fue el agresor es desconocido.[85] Lo que sí se sabe es que los Tyrannosaurus se alimentaban de diferentes especies de dinosaurios, y una ellas era Triceratops. Entre las pruebas que lo demuestran se incluye un ilion y un sacro de Triceratops con grandes marcas que solo podían haber sido dejadas por los grandes dientes de un tiranosaurio.[3]
En 2005, un documental de la BBC, La verdad sobre los dinosaurios asesinos, mostraba cómo un Triceratops pudo haberse defendido contra grandes depredadores como el Tyrannosaurus. Para ver si un Triceratops habría podido cargar contra otros dinosaurios como un rinoceronte moderno, se construyó un cráneo artificial de Triceratops y se propulsó sobre la piel simulada de un Tyrannosaurus a 24 kilómetros por hora. Los cuernos de la frente penetraron la piel, pero el cuerno de la nariz y el pico no pudieron, rompiéndose el frente del cráneo. La conclusión extraída fue que habría sido imposible que un Triceratops se defendiera de esta manera; en lugar de esto, probablemente se afirmaban contra el suelo cuando eran atacados por depredadores grandes, usando sus cuernos para cornear si se acercaban demasiado. Sin embargo, hay defectos en la metodología. El cráneo fue fijado a un marco de acero y no tenía la gama de movimiento o de compresión dada por una espina dorsal flexible y un cuerpo muscular. Un Triceratops sería incapaz de funcionar y de golpear algo de una manera tan artificial.
Combates con otros Triceratops

Las ilustraciones clásicas de Triceratops, además representarlos usando los cuernos en combate con depredadores, los muestran enfrentándose en combate entre sí con los cuernos trabados. Aunque los estudios demuestran que tal actividad sería factible, al igual que se observa en los animales con cuernos actuales,[86] sin embargo, no hay acuerdo sobre si lo hicieron realmente. Además, aunque aparezcan hoyos, agujeros, lesiones, y otros daños en los cráneos de Triceratops y en cráneos de otros ceratópsidos, que se atribuyen a menudo al daño por cuernos en combate, un estudio reciente señala que no se han encontrado restos de lesiones por el empuje de cuerno que causen estas formas de daño, por ejemplo no hay pruebas de infección o cura. En su lugar se sugieren como causas la resorción ósea no patológica, u otras enfermedades de los huesos desconocidas.[87] Sin embargo, un estudio más reciente comparó índices de incidencia de lesiones del cráneo en Triceratops y Centrosaurus, demostrado que estos eran constantes con Triceratops usando sus cuernos en combate y la gola adaptada como estructura protectora, mientras que un índice de patologías más baja en Centrosaurus puede indicar la representación visual más que el uso físico de la ornamentación craneal, o una forma de combate centrada en el cuerpo algo más que en la cabeza.[88] Se encontró que la frecuencia de lesiones era del 14 % en Triceratops.[89] Los investigadores también concluyeron que el daño encontrado en las muestras en el estudio a menudo estaba demasiado localizado para ser causado por una enfermedad ósea.[90] El examen histológico revela que el volante de Triceratops está compuesto de hueso fibrolamelar[91] que contiene fibroblastos que desempeñan un papel fundamental en la cicatrización de heridas y son capaces de depositar hueso rápidamente durante la remodelación.[92][93]
Se encontró un cráneo con un agujero en el hueso yugal, aparentemente una herida punzante sufrida mientras el animal estaba vivo, como lo indican los signos de curación. El agujero tiene un diámetro cercano al del extremo distal de un cuerno de Triceratops. Esta y otras heridas aparentemente curadas en los cráneos de los ceratopsianos se han citado como evidencia de una competencia intraespecífica no fatal en estos dinosaurios.[94][95] Otro espécimen, conocido como "Big John" tiene una fenestra similar a la escamosa causada por lo que parece ser otro cuerno de Triceratops y el hueso escamoso muestra signos de curación significativa, lo que reivindica aún más la hipótesis de que este ceratopsiano usó sus cuernos para el combate intraespecífico.[96]

Termorregulación y exhibición
La gola pudo haber ayudado aumentando el área expuesta y favoreciendo la termorregulación.[97] Una teoría similar fue propuesta para las placas del estegosaurio,[98] aunque este uso solamente no explicara la variación extraña y extravagante vista en diversos miembros de Ceratopsidae.[18] Esta observación es altamente sugestiva de lo que se cree ahora para ser la función primaria, la exhibición.
La teoría sobre el uso en la exhibición sexual fue propuesta por Davitashvili en 1961 y ha ganado muchos adeptos desde entonces.[45][82][99] Se ha sugerido que la exhibición visual era importante, en cortejo o en otro comportamiento social, es visto en el hecho de que los dinosaurios con cuernos tenían diferencias marcadas en sus adornos, haciendo de cada especie altamente distintiva. También, las criaturas vivas modernas con tales exhibiciones de cuernos y los adornos los utilizan en un comportamiento similar.[100] Un estudio reciente del más pequeño cráneo de un Triceratops, perteneciente fehacientemente a un joven, muestra que la gola y los cuernos se desarrollaron a una edad muy temprana, antes del desarrollo sexual y así probablemente importante de para el reconocimiento individual y de la especie en general.[101]
Crecimiento y ontogenia

En 2006, se publicó el primer estudio ontogenético extenso de Triceratops en la revista Proceedings of the Royal Society. El estudio, realizado por John R. Horner y Mark Goodwin, encontró que los individuos de Triceratops podría dividirse en cuatro grupos ontogenéticos generales, bebés, juveniles, subadultos y adultos. Con un número total de 28 cráneos estudiados, el más joven medía solo 38 centímetros de largo. Diez de los 28 cráneos podrían colocarse en orden en una serie de crecimiento con uno que represente cada edad. Se encontró que cada una de las cuatro etapas de crecimiento tenía características de identificación. Se descubrieron múltiples tendencias ontogenéticas, incluida la reducción del tamaño de los epoccipitales, el desarrollo y la reorientación de los cuernos postorbitales y el ahuecamiento de los cuernos.[102] Los ojos grandes y las características acortadas, un sello de "lindo" en los mamíferos bebé, también sugieren que los padres Triceratops pueden haber cuidado a sus jóvenes.
Paleoecología
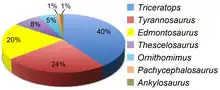
Triceratops vivió durante el Cretácico superior de América del Norte, sus fósiles provienen de la Formación Evanston, Formación Scollard, Formación Laramie, Formación Lance, Formación Denver y Formación Hell Creek.[103] Estas formaciones fósiles se remontan a la época de la extinción del Cretácico-Paleógeno, que data de hace 66 ± 0,07 millones de años.[104] Se han encontrado muchos animales y plantas en estas formaciones, pero principalmente de la Formación Lance y la Formación Hell Creek.[103] Triceratops fue uno de los últimos géneros de ceratopsianos en aparecer antes del final del Mesozoico. El relacionado Torosaurus, y el diminuto Leptoceratops, emparentado más distantemente, también estaban presentes, aunque sus restos son mucho más escasos.[5]

Los terópodos de estas formaciones incluyen géneros de dromeosáuridos, tiranosáuridos, ornitomímidos, troodóntidos,[103] avialanos,[105] y cenagnátidos.[106]
Los dromeosáuridos de la Formación Hell Creek son Acheroraptor y Dakotaraptor. Se conocen dromeosaurios indeterminados de otras formaciones fósiles. Los dientes comunes anteriormente referidos a Dromaeosaurus y Saurornitholestes más tarde se consideraron Acheroraptor.[107] Los tiranosáuridos de la formación son Nanotyrannus y Tyrannosaurus, aunque el primero podría ser un sinónimo menor del segundo. Entre los ornitomimidos se encuentran los géneros Struthiomimus y Ornithomimus;[103] un animal no descrito llamado "Orcomimus" podría ser de la formación.[108] Los troodóntidos sólo están representados por Pectinodon y Paronychodon en la Formación Hell Creek, con una posible especie de Troodon de la Formación Lance. Se conoce una especie de celurosaurio de Hell Creek y formaciones similares por una sola especie, Richardoestesia. Solo tres oviraptorosaurios son conocidos, dos de la Formación Hell Creek, Anzu, Leptorhynchos[106] y una especie gigante de cenagnátido, muy similar a Gigantoraptor, de Dakota del Sur. Sin embargo, solo se descubrieron huellas fosilizadas.[109] Los avialanos conocidos de la formación son Avisaurus,[103] múltiples especies de Brodavis,[110] y varias otras especies de hesperornitoformes, así como varias especies de aves verdaderas, incluyendo Cimolopteryx.[105]

Los ornitisquios abundan en las formaciones Scollard, Laramie, Lance, Denver y Hell Creek. Los principales grupos de ornitisquios son los anquilosaurianos, los ornitópodos, los ceratópsidos y los paquicefalosáuridos. Se conocen tres anquilosaurianos, Ankylosaurus, Denversaurus y posiblemente una especie de Edmontonia o un género no descrito relacionado. Se conocen múltiples géneros de ceratópsidos de la formación que no sean Triceratops, el leptoceratópsido Leptoceratops y los ceratópsidos casmosaurinos Torosaurus,[103] Nedoceratops y Tatankaceratops.[111] Los ornitópodos son comunes en la Formación Hell Creek, y se conocen de varias especies del ornitópodo Thescelosaurus y el hadrosáurido Edmontosaurus.[103][112] Se han encontrado varios paquicefalosaurios en la Formación Hell Creek y en formaciones similares. Entre ellos se encuentran los paquicefalosáuridos derivados Stygimoloch,[103] Dracorex,[113] Pachycephalosaurus,[103] Sphaerotholus, y un espécimen no descrito de Dakota del Norte. Los dos primeros podrían ser sinónimos menores de Pachycephalosaurus.
Los mamíferos abundan en la Formación Hell Creek. Los grupos representados incluyen multituberculados, metaterios y euterios. Los multituberculados representados incluyen Paracimexomys;[114] los cimolomiidos Paressonodon,[115] Meniscoessus, Essonodon, Cimolomys, Cimolodon y Cimexomys y los neoplagiaulacidos Mesodma y Neoplagiaulax. Los metatherianos están representados por los alfadóntido Alphadon, Protalphodon y Turgidodon; the pediomyids Pediomys,[114] Protolambda, and Leptalestes;[116] el estagodontido Didelphodon;[114] the deltateridiido Nanocuris; el herpetotheriido Nortedelphys;[115] y el glasbiido Glasbius. Se conocen unos pocos euterios, representados por Alostera,[114] Protungulatum,[116] los cimolestidos Cimolestes y Batodon, el gypsonictopsido Gypsonictops y el posible nyctiteriido Paranyctoides.[114]
En la cultura popular

El aspecto distintivo del Triceratops ha llevado a que sean representados con frecuencia en películas, juegos de computadoras y documentales. Aparecen en el film Parque Jurásico, donde uno es mostrado enfermo y tratado por humanos. Triceratops más activos se ven en las 4 secuelas del filme. También se han visto en tres documentales importantes sobre dinosaurios: Walking with Dinosaurs, The Truth About Killer Dinosaurs y Prehistoric Park. Aparecen en dos clásicos de Disney: Fantasía y Dinosaurio. Son generalmenrte llamados "tres-cuernos" y así son llamados en la película The Land Before Time y sus numerosas secuelas debido a los prominentes cuernos en la frente y nariz, los cuales han llegado a ser casi sinónimos con los dinosaurios. Los triceratops también aparecen en la palícula Ice Age: Dawn of the Dinosaurs. El diminutivo "Trike" es otro nombre informal de los personajes Triceratops en la literatura infantil y en series de televisión. En la serie Power Rangers, el Triceratops fue el dinosaurio del Blue Ranger durante las primeras tres temporadas[117] y luego en Power Rangers Dino Thunder. Aparece en la animada Harry and His Bucket Full of Dinosaurs. Baby Bop, personaje de la serie de televisión infantil Barney y sus amigos, es otro ejemplo de Triceratops en la cultura popular.

Una escena recurrente, en especial en los libros infantiles sobre dinosaurios, es la pelea entre un Triceratops y un T. rex.[118][119][120][121] Como tales estos dos dinosaurios se representan y se piensan a menudo como enemigos naturales. Una batalla memorable, pero anacrónica con un ceratosaurio sustituye al T. rex en la película 1966 One Million Years B.C.; otro ejemplo es en la serie japonesa Dino Rey, donde aparece Gabu, un triceratops con elemento de electricidad y que forma parte de los dinosaurios protagonistas del anime. Aparece en PDFC, pelea contra un Tyrannosaurus, Suchomimus y Velociraptor.
El Triceratops aparece en los videojuegos derivados directamente de la serie de Jurassic Park o similares, a saber los juegos 1997 para PC Jurassic Park: Chaos Island, Turok: Dinosaur Hunter, el juego del 2000 para PC y PS Dino Crisis 2, el juego de 2003 Jurassic Park: Operation Genesis, en 2015 en Ark: Survival Evolved, y en 2018 en Jurassic World:Evolution. También se incluye en la expansión para la franquicia Zoo Tycoon. Es una criatura popular usada en los juegos diseñados por Rareware, incluyendo Diddy Kong Racing y Star Fox Adventures. Triceratops, sin especificar la especie, es el fósil estatal de Dakota del Sur,[122] y dinosaurio oficial de Wyoming.[123]
El Triceratops también aparece representado en la franquicia Digimon, con la criatura conocida como Triceramon[124], quien es un Digimon de nivel Perfecto (o Mega Campeón, en el doblaje español) y una de las posibles evoluciones de Greymon[125] o Tyranomon[126], entre otros monstruos de la franquicia.
Véase también
Referencias
- Liddell y Scott (1980). Greek-English Lexicon, Abridged Edition. Oxford (Reino Unido): Oxford University Press. ISBN 0-19-910207-4.
- Lehman T. M. (1987). «Late Maastrichtian paleoenvironments and dinosaur biogeography in the Western Interior of North America». Paleogeography, Paleoclimatology and Paleoecology 60: 290. doi:10.2307/2406631.
- Erickson, G. M.; Olson, K. H. (1996). «Bite marks attributable to Tyrannosaurus rex: preliminary description and implications». Journal of Vertebrate Paleontology 16 (1): 175-178.
- Lambert, D. (1993). The ultimate dinosaur book. Dorling Kindersley, New York. pp. 152–167. ISBN 1-56458-304-X.
- Dodson, P. (1996). The horned dinosaurs. Princeton, New Jersey: Princeton University Press. ISBN 0-691-02882-6.
- «New analyses of dinosaur growth may wipe out one-third of species». Science News. ScienceDaily.com. 31 de octubre de 2009. Consultado el 3 de noviembre de 2009.
- «Melbourne Museum acquires world's most complete triceratops skeleton in 'immense' dinosaur deal». the Guardian (en inglés). 2 de diciembre de 2020. Archivado desde el original el 18 de febrero de 2021. Consultado el 10 de febrero de 2021.
- «DinoDictionary.com : T Dinosaurs Page 2». Archivado desde el original el 11 de mayo de 2020. Consultado el 16 de enero de 2008.
- Triceratops in The Natural History Museum's Dino Directory Archivado el 17 de febrero de 2009 en Wayback Machine.
- Alexander, R.M. (1985). «Mechanics of posture and gait of some large dinosaurs». Zoological Journal of the Linnean Society 83: 1-25. doi:10.1111/j.1096-3642.1985.tb00871.x.
- Seebacher, F. (2001). «A new method to calculate allometric length-mass relationships of dinosaurs». Journal of Vertebrate Paleontology 21 (1): 51-60. JSTOR 4524171. S2CID 53446536. doi:10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2.
- Paul, G. S. (2010). The Princeton Field Guide to Dinosaurs. Princeton University Press. pp. 265–267. ISBN 978-0-691-13720-9.
- Holtz, Thomas R. Jr. (2012). Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. «Winter 2011 Appendix ».
- Dinosaur facts
- «A Triceratops Named 'Kelsey'». www.bhigr.com. Archivado desde el original el 23 de diciembre de 2017. Consultado el 22 de diciembre de 2017.
- «Kesley the Triceratops». www.bhigr.com.
- Scannella, J.; Horner, J.R. (2010). «Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae): synonymy through ontogeny». Journal of Vertebrate Paleontology 30 (4): 1157-1168. S2CID 86767957. doi:10.1080/02724634.2010.483632.
- Dodson, P.; Forster, C. A. y Sampson, S. D. (2004) «Ceratopsidae». En: Weishampel, D. B.; Dodson, P. y Osmólska, H. (eds.) The Dinosauria (2.ª ed.). Berkeley: University of California Press. Págs. 494–513. ISBN 0-520-24209-2.
- «Denver museum unveils 7-foot-long, 1,000-pound Triceratops skull». The Daily Courier. 18 de noviembre de 2003. Archivado desde el original el 19 de mayo de 2021. Consultado el 26 de diciembre de 2013.
- Scannella, John B.; Fowler, Denver W.; Goodwin, Mark B.; Horner, John R. (15 de julio de 2014). «Evolutionary trends in Triceratops from the Hell Creek Formation, Montana». Proceedings of the National Academy of Sciences (en inglés) 111 (28): 10245-10250. Bibcode:2014PNAS..11110245S. ISSN 0027-8424. PMC 4104892. PMID 24982159. doi:10.1073/pnas.1313334111.
- «Making A Triceratops. Science Supplies Missing Part! Of Skeleton». Boston Evening Transcript. 24 de octubre de 1901. Archivado desde el original el 19 de mayo de 2021. Consultado el 26 de diciembre de 2013.
- Dodson, P.; Forster, C. A. y Sampson, S. D. (2004) «Ceratopsidae». En: Weishampel, D. B.; Dodson, P. y Osmólska, H. (eds.) The Dinosauria (2.ª ed.). Berkeley: University of California Press. pp. 494–513. ISBN 0-520-24209-2.
- Brown, B. y Schlaikjer, E. M. (1940). «The origin of ceratopsian horn-cores». American Museum Novitates, 1065: 1–7
- Fujiwara, Shin-Ichi (12 de diciembre de 2009). «A reevaluation of the manus structure in Triceratops (Ceratopsia: Ceratopsidae)». Journal of Vertebrate Paleontology 29 (4): 1136-1147. ISSN 0272-4634. S2CID 86519018. doi:10.1671/039.029.0406.
- Christiansen, P., and Paul, G. S. (2001). «Limb bone scaling, limb proportions, and bone strength in neoceratopsian dinosaurs». Gaia 16:1 3–29.
- Chapman, R.E.; Snyder, R.A.; Jabo, S. y Andersen, A. (2001). «On a new posture for the horned dinosaur Triceratops». Journal of Vertebrate Paleontology 21 (suplemento al número 3), Abstracts of Papers, 61st Annual Meeting: 39A–40A.
- Martin, Anthony J. (2006). Introduction to the study of dinosaurs (2nd edición). Malden, MA: Blackwell Pub. ISBN 978-1405134132. OCLC 61130756.
- Witton, Mark (24 de diciembre de 2015). «Mark Witton.com Blog: Dinosaur scales: some thoughts for artists». Mark Witton.com Blog. Archivado desde el original el 18 de junio de 2021. Consultado el 25 de junio de 2021.
- Carpenter, K. (2006). «"Bison" alticornis and O.C. Marsh's early views on ceratopsians.» En: Carpenter, K. (ed.). Horns and beaks: ceratopsian and ornithopod dinosaurs Bloomington e Indianapolis: Indiana University Press. Págs. 349–364. ISBN 0-253-34817-X.
- Marsh, O.C. (1887). «Notice of new fossil mammals». American Journal of Science 34: 323–331.
- Marsh, O.C. (1888). «A new family of horned Dinosauria, from the Cretaceous». American Journal of Science 36: 477–478.
- Marsh, O.C. (1889a). «Notice of new American Dinosauria». American Journal of Science 37 (220): 331-336. Bibcode:1889AmJS...37..331M. S2CID 131729220. doi:10.2475/ajs.s3-37.220.331. Archivado desde el original el 29 de septiembre de 2018. Consultado el 19 de octubre de 2021.
- Marsh, O.C. (1889b). «Notice of gigantic horned Dinosauria from the Cretaceous». American Journal of Science 38: 173–175
- Hatcher, J. B., Marsh, O. C. and Lull, R. S. (1907) The Ceratopsia. Government Printing Office, Washington, D.C. ISBN 0-405-12713-8.
- Breithaupt, B. H. (1999). «First Discovery of Dinosaurs in the American West». En Gillette, D. D., ed. Vertebrate Paleontology In Utah. Utah Geological Survey. pp. 59–65. ISBN 978-1-55791-634-1.
- Cope, E.D. (1872). "On the existence of Dinosauria in the Transition Beds of Wyoming". Proceedings of the American Philosophical Society. 12: 481–483.
- Cope, E.D. (1874). Report on the stratigraphy and Pliocene vertebrate paleontology of northern Colorado. Bulletin of the U.S. Geological and Geographical Survey of the Territories. 9:9-28.
- Lull, R. S., & Wright, N. E. (1942). Hadrosaurian dinosaurs of North America(Vol. 40). Geological Society of America.
- Lull, R. S. (1933) «A revision of the Ceratopsia or horned dinosaurs». Memoirs of the Peabody Museum of Natural History, 3(3): 1–175.
- «Division of Paleontology». research.amnh.org. Consultado el 12 de abril de 2022.
- Dodson, P., Forster, C.A., and Sampson, S.D. (2004). Ceratopsidae. In: Weishampel, D.B., Dodson, P., and Osmólska, H. (eds.). The Dinosauria (second edition). University of California Press:Berkeley, 494-513. ISBN 0-520-24209-2.
- Forster, C. A. (1996). «Species resolution in Triceratops: cladistic and morphometric approaches». Journal of Vertebrate Paleontology, 16: 259–270
- Sternberg, C. M. (1949). The Edmonton fauna and description of a new Triceratops from the Upper Edmonton member; phylogeny of the Ceratopsidae. National Museum of Canada Bulletin 113:33–46.
- Ostrom, J. H. y Wellnhofer, P. (1986). «The Munich specimen of Triceratops with a revision of the genus». Zitteliana, 14: 111–158
- Lehman, T. M. (1990). The ceratopsian subfamily Chasmosaurinae: sexual dimorphism and systematics. in: Carpenter, K., and Currie, P. J. (eds.). Dinosaur Systematics: Perspectives and Approaches, Cambridge University Press, Cambridge, pp. 211–229. ISBN 0-521-36672-0.
- Forster, C. A. (1996). «Species resolution in Triceratops: cladistic and morphometric approaches». Journal of Vertebrate Paleontology, 16(2): 259–270
- Lehman, T. M. (1998). «A gigantic skull and skeleton of the horned dinosaur Pentaceratops sternbergi from New Mexico». Journal of Paleontology, 72(5): 894–906
- Scannella, J.B. y Fowler, D.W. (2009). «Anagenesis in Triceratops: evidence from a newly resolved stratigraphic framework for the Hell Creek Formation». En: 9th North American Paleontological Convention Abstracts. Cincinnati Museum Center Scientific Contributions, 3: 148–149
- Farke, A. A. (2006). «Cranial osteology and phylogenetic relationships of the chasmosaurine ceratopsid Torosaurus latus». En Carpenter, K., ed. Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs. Bloomington: Indiana University Press. pp. 235-257. ISBN 978-0-253-34817-3.
- «New analyses of dinosaur growth may wipe out one-third of species».
- Farke, Andrew A. (2011). «Anatomy and taxonomic status of the chasmosaurine ceratopsid Nedoceratops hatcheri from the Upper Cretaceous Lance Formation of Wyoming, U.S.A». En Claessens, Leon, ed. PLOS ONE 6 (1): e16196. Bibcode:2011PLoSO...616196F. PMC 3024410. PMID 21283763. doi:10.1371/journal.pone.0016196.
- Nicholas R. Longrich (2011). «Titanoceratops ouranous, a giant horned dinosaur from the Late Campanian of New Mexico». Cretaceous Research 32 (3): 264-276. doi:10.1016/j.cretres.2010.12.007. Archivado desde el original el 16 de agosto de 2016. Consultado el 6 de noviembre de 2011.
- Longrich, Nicholas R.; Field, Daniel J. (29 de febrero de 2012). «Torosaurus Is Not Triceratops: Ontogeny in Chasmosaurine Ceratopsids as a Case Study in Dinosaur Taxonomy». PLOS ONE 7 (2): e32623. Bibcode:2012PLoSO...732623L. ISSN 1932-6203. PMC 3290593. PMID 22393425. doi:10.1371/journal.pone.0032623.
- Bowdler, Neil (1 de marzo de 2012). «Triceratops and Torosaurus dinosaurs 'two species, not one'». BBC News (en inglés británico). Archivado desde el original el 15 de marzo de 2013. Consultado el 29 de julio de 2013.
- Scannella, J. B.; Horner, J. R. (2011). «'Nedoceratops': An Example of a Transitional Morphology». En Claessens, Leon, ed. PLOS ONE 6 (12): e28705. Bibcode:2011PLoSO...628705S. PMC 3241274. PMID 22194891. doi:10.1371/journal.pone.0028705.
- Lambe, L.M. (1915). On Eoceratops canadensis, gen. nov., with remarks on other genera of Cretaceous horned dinosaurs. Canada Department of Mines Geological Survey Museum Bulletin 12:1–49.
- Ostrom, J. H. (1966). Functional morphology and evolution of the ceratopsian dinosaurs. Evolution 20:220–227.
- Norman, David (1985). The Illustrated Encyclopaedia of Dinosaurs. London: Salamander Books. ISBN 0-517-468905.
- Dodson, P., and Currie, P. J. (1990). Neoceratopsia. 593–618. in Weishampel, D. B., Dodson, P., & Osmólska, H. (eds.). The Dinosauria University of California Press:Berkeley, pp. 593–618. ISBN 0-520-06727-4.
- Dodson, P. (1993). Comparative Craniology of the Ceratopsia. In: American Journal of Science 293, pp. 200–234.
- Dodson, P. (1996). The Horned Dinosaurs. Princeton University Press:Princeton, New Jersey, p. 244. ISBN 0-691-02882-6.
- Gauthier, J. A. (1986). Saurischian monophyly and the origin of birds. The Origin of Birds and the Evolution of Flight, K. Padian (ed.), Memoirs of the California Academy of Sciences 8:1–55.
- Sereno, P. C. (1998). A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 210(1):41–83.
- Scott D. Sampson, Mark A. Loewen, Andrew A. Farke, Eric M. Roberts, Catherine A. Forster, Joshua A. Smith, and Alan A. Titus (2010). «New Horned Dinosaurs from Utah Provide Evidence for Intracontinental Dinosaur Endimism». PLoS ONE. 5 5 (9): e12292. PMC 2929175. PMID 20877459. doi:10.1371/journal.pone.0012292.
- Longrich, N. R. (2014). «The horned dinosaurs Pentaceratops and Kosmoceratops from the upper Campanian of Alberta and implications for dinosaur biogeography». Cretaceous Research 51: 292-308. doi:10.1016/j.cretres.2014.06.011.
- Mathews, Joshua C.; Brusatte, Stephen L.; Williams, Scott A.; Henderson, Michael D. (2009). «The first Triceratops bonebed and its implications for gregarious behavior». Journal of Vertebrate Paleontology 29 (1): 286-290.
- Smith, Matt (4 de junio de 2013). «Triceratops trio unearthed in Wyoming – CNN». CNN. Archivado desde el original el 1 de agosto de 2017. Consultado el 22 de diciembre de 2017.
- Illies, M. M. Canoy; Fowler, D. W. (2020). «Triceratops with a kink: Co-ossification of five distal caudal vertebrae from the Hell Creek Formation of North Dakota». Cretaceous Research 108: 104355. S2CID 214215413. doi:10.1016/j.cretres.2019.104355.
- Barrera, Nathanial A. (9 de enero de 2020). «More than old bones: New study sheds light on Triceratops behavior and living habits». The Dickinson Press (en inglés). Archivado desde el original el 27 de mayo de 2020. Consultado el 31 de marzo de 2020.
- Erickson, B.R. (1966). «Mounted skeleton of Triceratops prorsus in the Science Museum». Scientific Publications of the Science Museum, 1: 1–16
- Dodson, P. (1996). The Horned Dinosaurs. Princeton, New Jersey: Princeton University Press, p. 79. ISBN 0-691-02882-6.
- Bakker, R.T. (1986). The dinosaur heresies: new theories unlocking the mystery of the dinosaurs and their extinction. Nueva York: William Morrow, p. 438. ISBN 0-14-010055-5.
- Derstler, Kraig (1994). «Dinosaurs of the Lance Formation in eastern Wyoming». En Nelson, Gerald E. (ed.), ed. The Dinosaurs of Wyoming. Wyoming Geological Association Guidebook, 44th Annual Field Conference. Wyoming Geological Association. pp. 127-146.
- Sakagami, Rina; Kawabe, Soichiro (2020). «Endocranial anatomy of the ceratopsid dinosaur Triceratops and interpretations of sensory and motor function». PeerJ 8: e9888. PMC 7505063. PMID 32999761. doi:10.7717/peerj.9888. Archivado desde el original el 19 de mayo de 2021. Consultado el 18 de septiembre de 2020.
- Wiemann, J.; Menéndez, I.; Crawford, J.M.; Fabbri, M.; Gauthier, J.A.; Hull, P.M.; Norell, M.A.; Briggs, D.E.G. (2022). «Fossil biomolecules reveal an avian metabolism in the ancestral dinosaur». Nature 606 (7914): 522-526. PMID 35614213. S2CID 249064466. doi:10.1038/s41586-022-04770-6.
- Tait J., and Brown, B. (1928). How the Ceratopsia carried and used their head. Transactions of the Royal Society of Canada. 22:13–23.
- Ostrom, J. H. (1966). «Functional morphology and evolution of the ceratopsian dinosaurs». Evolution 20 (3): 290. doi:10.2307/2406631.
- Ostrom, J. H. (1964). «A functional analysis of jaw mechanics in the dinosaur Triceratops». Postilla, Yale Peabody Museum 88: 1–35.
- Weishampel, D. B. (1984). «Evolution of jaw mechanisms in ornithopod dinosaurs». Advances in Anatomy, Embryology, and Cell Biology, 87: 1–110.
- Coe, M. J.; Dilcher, D. L.; Farlow, J. O.; Jarzen, D. M. y Russell, D. A. (1987). «Dinosaurs and land plants». En: Friis, E. M., Chaloner, W. G. y Crane, P. R. (eds.) The Origins of Angiosperms and their Biological Consequences Cambridge University Press, pp. 225–258. ISBN 0-521-32357-6.
- Lull, R. S. (1908). «The cranial musculature and the origin of the frill in the ceratopsian dinosaurs». American Journal of Science, 4(25): 387–399
- Forster, C. A. (1990). The cranial morphology and systematics of Triceratops, with a preliminary analysis of ceratopsian phylogeny. Tesis doctoral. Universidad de Pennsylvania, Philadelphia. 227 pp.
- Sternberg, C. H. (1917). Hunting Dinosaurs in the Badlands of the Red Deer River, Alberta, Canada. Published by the author, San Diego, California, 261 pp.
- Bakker, R. T. (1986). The dinosaur heresies: new theories unlocking the mystery of the dinosaurs and their extinction. Nueva York: William Morrow. ISBN 0-14-010055-5.
- Happ, John; Carpenter, Kenneth (2008). «An analysis of predator-prey behavior in a head-to-head encounter between Tyrannosaurus rex and Triceratops». En Carpenter, Kenneth y Larson, Peter E., ed. Tyrannosaurus rex, the tyrant king (Life of the Past). Bloomington: Indiana University Press. pp. 355–368. ISBN 0-253-35087-5.
- Farke, A. A. (2004). Horn use in Triceratops (Dinosauria: Ceratopsidae): Testing behavioral hypotheses using scale models. Palaeontologia electronica, 7(1): 10 pp.
- Tanke, D. H, y Farke, A. A. (2006). «Bone resorption, bone lesions, and extracranial fenestrae in ceratopsid dinosaurs: a preliminary assessment». En: Carpenter, K. (ed.). Horns and beaks: ceratopsian and ornithopod dinosaurs Bloomington: Indiana University Press. Págs. 319–347. ISBN 0-253-34817-X.
- Farke, A. A.; Wolff, E.D.S. y Tanke, D.H. (2009). «Evidence of combat in Triceratops». PLoS ONE, 4(1): e4252. doi 10.1371/journal.pone.0004252
- Peterson, JE; Dischler, C; Longrich, NR (2013). «Distributions of Cranial Pathologies Provide Evidence for Head-Butting in Dome-Headed Dinosaurs (Pachycephalosauridae)». PLOS ONE 8 (7): e68620. Bibcode:2013PLoSO...868620P. PMC 3712952. PMID 23874691. doi:10.1371/journal.pone.0068620.
- Wall, Michael (27 de enero de 2009). «Scars Reveal How Triceratops Fought –». Wired. Archivado desde el original el 12 de enero de 2014. Consultado el 3 de agosto de 2010.
- Reid, R.E.H. (1997). «Histology of bones and teeth». En Currie, P. J.; Padian, K., eds. Encyclopedia of Dinosaurs. San Diego, CA.: Academic Press. pp. 329-339.
- Horner, JR; Goodwin, MB (2009). «Extreme Cranial Ontogeny in the Upper Cretaceous Dinosaur Pachycephalosaurus». PLOS ONE 4 (10): e7626. Bibcode:2009PLoSO...4.7626H. PMC 2762616. PMID 19859556. doi:10.1371/journal.pone.0007626.
- Horner, JR; Lamm, E (2011). «Ontogeny of the parietal frill of Triceratops: a preliminary histological analysis». Comptes Rendus Palevol 10 (5–6): 439-452. doi:10.1016/j.crpv.2011.04.006.
- Farlow, J. O.; Dodson, P. (1975). «The behavioral significance of frill and horn morphology in ceratopsian dinosaurs». Evolution 29 (2): 353-361. JSTOR 2407222. PMID 28555861. doi:10.2307/2407222.
- Martin, A. J. (2006). Introduction to the Study of Dinosaurs (Second edición). Oxford: Blackwell Publishing. pp. 299-300. ISBN 978-1-4051-3413-2.
- D’Anastasio, Ruggero; Cilli, Jacopo; Bacchia, Flavio; Fanti, Federico; Gobbo, Giacomo; Capasso, Luigi (7 de abril de 2022). «Histological and chemical diagnosis of a combat lesion in Triceratops». Scientific Reports (en inglés) 12 (1): 3941. Bibcode:2022NatSR..12.3941D. ISSN 2045-2322. PMC 8990019. PMID 35393445. doi:10.1038/s41598-022-08033-2.
- Wheeler, P.E. (1978). «Elaborate CNS cooling structures in large dinosaurs». Nature 275 (275): 441-443. doi:10.1038/275441a0.
- Farlow, J. O., Thompson, C. V., and Rosner, D. E. (1976). «Plates of the dinosaur Stegosaurus: Forced convection heat loss fins?». Science 192: 1123. PMID 17748675. doi:10.1126/science.192.4244.1123.
- Davitashvili L (1961). The Theory of sexual selection. Izdatel'stvo Akademia nauk SSSR. p. 538.
- Farlow, J. O., and Dodson, P. (1975). «The behavioral significance of frill and horn morphology in ceratopsian dinosaurs». Evolution 29 (2): 353. doi:10.2307/2407222.
- Goodwin, M. B., Clemens, W. A., Horner, J. R., and Padian, K. (2006). «The smallest known Triceratops skull: new observations on ceratopsid cranial anatomy and ontogeny.» (PDF). Journal of Vertebrate Paleontology 26 (1): 103. doi:10.1671/0272-4634(2006)26[103:TSKTSN]2.0.CO;2. Archivado desde el original el 5 de noviembre de 2015. Consultado el 28 de noviembre de 2009.
- Horner, J.R.; Goodwin, M.B. (2006). «Major cranial changes during Triceratops ontogeny». Proceedings of the Royal Society B: Biological Sciences 273 (1602): 2757-2761. PMC 1635501. PMID 17015322. doi:10.1098/rspb.2006.3643.
- Weishampel, D.B.; Dodson, Peter; Osmólska, H. (2004). The Dinosauria (Second edición). Berkeley: University of California Press. p. 861. ISBN 978-0-520-24209-8.
- Husson, D. E.; Galbrun, B.; Laskar, J.; Hinnov, L. A.; Thibault, N.; Gardin, S.; Locklair, R. E. (2011). «Astronomical calibration of the Maastrichtian (Late Cretaceous)». Earth and Planetary Science Letters 305 (3–4): 328-340. Bibcode:2011E&PSL.305..328H. doi:10.1016/j.epsl.2011.03.008.
- Longrich, N. R.; Tokaryk, T.; Field, D. J. (2011). «Mass extinction of birds at the Cretaceous-Paleogene (K-Pg) boundary». Proceedings of the National Academy of Sciences 108 (37): 15253-15257. Bibcode:2011PNAS..10815253L. PMC 3174646. PMID 21914849. doi:10.1073/pnas.1110395108.
- Lamanna, M. C.; Sues, H. D.; Schachner, E. R.; Lyson, T. R. (2014). «A New Large-Bodied Oviraptorosaurian Theropod Dinosaur from the Latest Cretaceous of Western North America». PLOS ONE 9 (3): e92022. Bibcode:2014PLoSO...992022L. PMC 3960162. PMID 24647078. doi:10.1371/journal.pone.0092022.
- Evans, D. C.; Larson, D. W.; Currie, P. J. (2013). «A new dromaeosaurid (Dinosauria: Theropoda) with Asian affinities from the latest Cretaceous of North America». Naturwissenschaften 100 (11): 1041-1049. Bibcode:2013NW....100.1041E. PMID 24248432. S2CID 14978813. doi:10.1007/s00114-013-1107-5.
- Triebold, M. (1997). «The Sandy site: Small dinosaurs from the Hell Creek Formation of South Dakota». En Wolberg, D.; Stump, E.; Rosenberg, G., eds. Dinofest International: Proceedings of a Symposium: 245-248.
- Maltese, Anthony (17 de diciembre de 2013). «Giant Oviraptor Tracks from the Hell Creek». RMDRC paleo lab. Archivado desde el original el 12 de noviembre de 2020. Consultado el 17 de diciembre de 2013.
- Martin, L. D.; Kurochkin, E. N.; Tokaryk, T. T. (2012). «A new evolutionary lineage of diving birds from the Late Cretaceous of North America and Asia». Palaeoworld 21: 59-63. doi:10.1016/j.palwor.2012.02.005.
- Ott, C.J.; Larson, P.L. (2010). «A New, Small Ceratopsian Dinosaur from the Latest Cretaceous Hell Creek Formation, Northwest South Dakota, United States: A Preliminary Description». En Ryan, M.J.; Chinnery-Allgeier, B.J.; Eberth, D.A., eds. New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium. Bloomington: Indiana University Press. p. 656.
- Campione, N. S. E.; Evans, D. C. (2011). «Cranial Growth and Variation in Edmontosaurs (Dinosauria: Hadrosauridae): Implications for Latest Cretaceous Megaherbivore Diversity in North America». PLOS ONE 6 (9): e25186. Bibcode:2011PLoSO...625186C. PMC 3182183. PMID 21969872. doi:10.1371/journal.pone.0025186.
- Bakker, R.T.; Sullivan, R.M.; Porter, V.; Larson, P.; Saulsbury, S.J. (2006). «Dracorex hogwartsia, n. gen., n. sp., a spiked, flat-headed pachycephalosaurid dinosaur from the Upper Cretaceous Hell Creek Formation of South Dakota». En Lucas, S.G.; Sullivan, R.M., eds. Late Cretaceous Vertebrates from the Western Interior. New Mexico Museum of Natural History and Science Bulletin 35: 331-345. Archivado desde el original el 19 de noviembre de 2018. Consultado el 6 de enero de 2015.
- Kielan-Jaworowska, Zofia; Cifelli, Richard L.; Luo, Zhe-Xi (2004). Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure. New York: Columbia University Press. pp. 98-99. ISBN 978-0-231-11918-4.
- Wilson, G. P. (2013). «Mammals across the K/Pg boundary in northeastern Montana, U.S.A.: Dental morphology and body-size patterns reveal extinction selectivity and immigrant-fueled ecospace filling». Paleobiology 39 (3): 429-469. S2CID 36025237. doi:10.1666/12041.
- Archibald, J. D.; Zhang, Y.; Harper, T.; Cifelli, R. L. (2011). «Protungulatum, Confirmed Cretaceous Occurrence of an Otherwise Paleocene Eutherian (Placental?) Mammal». Journal of Mammalian Evolution 18 (3): 153-161. S2CID 16724836. doi:10.1007/s10914-011-9162-1.
- «Ranger Central». Archivado desde el original el 9 de septiembre de 2010.
- Geis, D. (1982). How and Why Book of Dinosaurs. Allan Publishers. ISBN 0-8241-5001-5.
- Watson, J. W. (1981). The Giant Golden Book of Dinosaurs. Random House Children's Books. ISBN 0-307-13764-3.
- Pistorius A. (1958) What dinosaur is it? Follett Pub. Co., Chicago.
- Wise, W. (1963) In The Time of The Dinosaurs. Scholastic. ISBN 0-590-32351-2.
- State of South Dakota. «Signs and Symbols of South Dakota.....». Archivado desde el original el 14 de enero de 2007. Consultado el 20 de enero de 2007.
- State of Wyoming. «State of Wyoming – General Information». Archivado desde el original el 10 de febrero de 2007. Consultado el 20 de enero de 2007.
- «Triceramon | Digimon Encyclopedia». Digimon Web | Digimon Official General Site (en inglés). Consultado el 29 de abril de 2023.
- «グレイモン | デジモン図鑑». デジモンウェブ | デジモン公式総合サイト (en japonés). Consultado el 29 de abril de 2023.
- «Tyrannomon | Digimon Encyclopedia». Digimon Web | Digimon Official General Site (en inglés). Consultado el 29 de abril de 2023.
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikispecies tiene un artículo sobre Triceratops.
Wikispecies tiene un artículo sobre Triceratops. Wikimedia Commons alberga una categoría multimedia sobre Triceratops.
Wikimedia Commons alberga una categoría multimedia sobre Triceratops.
| Control de autoridades |
|
|---|
 Datos: Q14384
Datos: Q14384- Multimedia: Triceratops / Q14384
- Especies: Triceratops