Edmontonia
Edmontonia ("de Edmonton") es un género representado por dos o tres especies según el autor, de dinosaurios tireóforos nodosáuridos, que vivió a finales del período Cretácico, hace aproximadamente 83 y 65 millones de años, en el Campaniense y el Maastrichtiense, en lo que hoy es Norteamérica. Las especies son Edmontonia longiceps, la especie tipo, Edmontonia rugosidens y una tercera, a veces incluida en Glyptodontopelta mimus, Edmontonia australis.
| Edmontonia | ||
|---|---|---|
| Rango temporal: 83 Ma - 66 Ma Cretácico superior | ||
 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Ornithischia | |
| Suborden: | Thyreophora | |
| Infraorden: | Ankylosauria | |
| Familia: | Nodosauridae | |
| Género: |
Edmontonia Sternberg, 1928 | |
| Especie tipo | ||
|
Edmontonia longiceps Sternberg, 1928 | ||
| Especies | ||
| ||
| Sinonimia | ||
| ||
Descripción
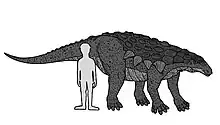
Edmontonia fue un dinosaurio voluminoso y armado de alrededor de 6,6 metros de largo y 2 de alto.[1] Tenía placas óseas pequeñas y ovales en la espalda y la cabeza, y muchas puntas afiladas a lo largo de sus lados. Los cuatro picos más grandes sobresalían de los hombros en cada lado, el segundo de los cuales se dividió en especímenes de E. rugosidens. Su cráneo tenía forma de pera cuando se veía por arriba.[1] Su cuello y hombros estaban protegidos por tres medios anillos hechos de grandes placas de quilla. En 2010, Gregory S. Paul consideró que ambas especies principales de Edmontonia, E. longiceps y E. rugosidens, son igualmente largas a 6 metros y pesan 3 toneladas[2]
En 1990, Kenneth Carpenter estableció algunos rasgos diagnósticos para el género como un todo, principalmente comparándolo con su pariente cercano Panoplosaurus. En la vista superior, el hocico tiene lados más paralelos. La armadura del cráneo tiene una superficie lisa. En el paladar, el vomer se hunde. Los arcos neurales y las espinas neurales son más cortos que los de Panoplosaurus . El sacro propiamente dicho consiste en tres vértebras sacras. En la cintura escapular, la escápula y la coracoides no están fusionadas.[3]
Carpenter también indicó de qué manera las principales especies difieren entre sí. La especie tipo, E. longiceps, se distingue de E. rugosidens por carecer de osteodermos que sobresalen lateralmente detrás de las cuencas oculare, por tener filas dentales que son menos divergentes, poseyendo un paladar más estrecho, tiene un sacro que es más ancho que largo y más robusto; y al tener espinas más cortas a los lados. Además, no se ha encontrado una placa de mejilla osificada, conocida de E. rugosidens, como en E. longiceps.[3]
Esqueleto

El cráneo de Edmontonia, de hasta medio metro de largo, es algo alargado con un hocico truncado que sobresale. El hocico llevaba un pico superior córneo y los huesos del hocico delantero, los premaxilares , no tenían dientes. El borde cortante del pico superior continuó en las filas de dientes superiores, cada uno con catorce a diecisiete dientes pequeños. En cada dentario de las mandíbulas inferiores, dieciocho a veintiún dientes estaban presentes. En los lados del hocico estaban presentes grandes depresiones, "vestíbulos nasales", que cada uno poseía dos aberturas más pequeñas. La parte superior era un óvalo horizontal y representaba la fosa nasal externa ósea, la entrada a la cavidad nasal, el pasaje de aire normal. La segunda abertura más redondeada por debajo y oblicuamente por delante, era la entrada a un tracto "paranasal", que corre a lo largo del lado externo de la cavidad nasal, en una posición algo inferior. Un estudio de Matthew Vickaryous en 2006 demostró por primera vez la presencia de aperturas múltiples en un nodosaurido; tales estructuras ya habían sido bien establecidas en anquilosáuridos. Sin embargo, los tractos aéreos son mucho más simples que en la condición anquilosáurida típica, y no son intrincados a la vez que carecen de huesos de cornetes óseos . La cavidad nasal está separada en dos mitades a lo largo de la línea media por una pared ósea. Este septo es continuado hasta abajo por los vómer , que están aquillados, la quilla presenta un apéndice en forma de péndulo.[4] Otra similitud con Ankylosauridae es la presencia de un paladar óseo secundario, un posible caso de evolución paralela . Esto también se ha demostrado en Panoplosaurus.[3]
las placas de la armadura de la cabeza, son lisos. Los detalles difieren entre los diferentes especímenes, pero todos comparten una gran placa nasal central en el hocico, dos grandes placas en los bordes posteriores del hocico y una gran placa central en el techo del cráneo. Las placas detrás del borde del alvéolo superior en E. longiceps no sobresalen tanto como en E. rugosidens, combinado con un hocico más estrecho y puntiagudo en el anterior. Se conocen algunas muestras de E. rugosidens que poseen una "placa de mejilla" por encima de la mandíbula inferior. Al contrario de lo que se descubrió con Panoplosaurus, es de "flotación libre", no fusionado con el hueso de la mandíbula inferior.[5]

La columna vertebral contiene alrededor de ocho vértebras cervicales, alrededor de doce vértebras dorsales "libres", una "vara sacra" de cuatro vértebras dorsales dorsales fusionadas, tres vértebras sacras, dos caudosacrales y al menos veinte, pero probablemente unas cuarenta, vértebras de la cola. En el cuello, las dos primeras vértebras, el atlas y el axis están fusionadas. En la cintura escapular, el coracoide tiene un perfil rectangular, en contraste con la forma más redondeada con Panoplosaurus . Dos placas esternal están presentes, conectadas a las costillas esternal. La extremidad anterior es robusta pero relativamente larga. En Edmontonia longiceps y E. rugosidens, la cresta deltopectoral del húmero se redondea gradualmente. losel metacarpo es robusto comparado con el de Panoplosaurus . La mano muy probablemente era tetradactilo, tenía cuatro dedos.[3] Se desconoce el número exacto de falanges pero la fórmula fue de WP. Coombs sugirió que fuera 2-3-3-4- ?.[6]
Osteodermos
Además de la armadura de la cabeza, el cuerpo estaba cubierto de osteodermos, osificaciones de la piel. La configuración de la armadura de Edmontonia es relativamente bien conocida, mucho de lo que se descubrió en articulación. La región del cuello y el hombro estaba protegida por tres medias anillos cervicales, cada una de las cuales consistía en placas óseas rectangulares fusionadas, asimétricamente quilladas. Estos medios anillos no tenían una banda ósea subyacente continua. El primer y segundo medio anillos tenían tres pares de segmentos cada uno. Debajo de cada extremo inferior de la segunda mitad estaba presente una espiga lateral, un osteodermo triangular separado apuntando oblicuamente hacia adelante. En la tercera mitad sobre los hombros, los dos pares de segmentos centrales están bordeados en cada lado por una espiga muy grande que se bifurca hacia adelante, presentando un punto secundario sobre el principal. Una tercera espiga grande detrás de ella apunta más hacia los lados, un cuarto más pequeño, a menudo conectado al tercero en la base, se dirige oblicuamente hacia atrás. La fila de espinas laterales continúa en la parte trasera, pero allí los osteodermos son mucho más bajos, curvándose fuertemente hacia atrás, con el punto sobresaliendo por el borde posterior. Gilmore tuvo problemas para creer que los picos de los hombros realmente apuntaban al frente, ya que esto habría obstaculizado en gran medida al animal mientras se movía a través de la vegetación. Sugirió que los puntos habían cambiado durante el entierro del cuerpo. Sin embargo, Carpenter y Paul, tratando de reposicionar los picos, descubrieron que era imposible rotarlos sin perder la conformidad con el resto de la armadura. Los picos laterales tienen bases sólidas, no huecas. Los picos difieren en tamaño entre Los picos laterales tienen bases sólidas, no huecas. Los picos difieren en tamaño entre individuos de E. rugosidens, los del holotipo E. longiceps son relativamente pequeños.[3]
Detrás de la tercera mitad, la espalda y la cadera están cubiertas por numerosas filas transversales de osteodermos ovalados, mucho más pequeños y ovales. Estos no están ordenados en filas longitudinales. Las filas delanteras tienen placas orientadas a lo largo de la longitud del cuerpo, pero en la parte posterior, el eje longitudinal de estos osteodermos gira gradualmente hacia los lados, sus quillas en última instancia, se extienden transversalmente. Los rosetones faltan. La configuración de la armadura de cola es desconocida. Las placas más grandes de todas las partes del cuerpo estaban conectadas por pequeños osículos.[3] Estas pequeñas escudetes redondas también cubren la garganta.[4]
Descubrimiento e investigación
Fue encontrado en la Formación Cañón Herradura, antiguamente llamada formación Edmonton de donde proviene su nombre, en Alberta Canadá. La especie tipo de Edmontonia, E. longiceps fue descubierta en 1924 por George Paterson. Sin embargo no fue nombrada hasta 1928 por Charles M. Sternberg. Una segunda especie fue nombrada E. rugosidens, descrita formalmente por Charles W. Gilmore en 1930, a partir de restos de la Formación Aguja de Texas EE. UU.. En 1915, el Museo Americano de Historia Natural obtuvo la mitad frontal casi completa y articulada de un dinosaurio blindado, encontrado el mismo año por Barnum Brown en Alberta , Canadá. En 1922, William Diller Matthew remitió este espécimen, AMNH 5381, a Palaeoscincus en un artículo de ciencia popular, sin indicar ninguna especie en particular.[7] Se pretendía nombrar una nueva especie de Palaeoscincus en cooperación con Brown, pero su artículo nunca se publicó.[3] Matthew también refirió el espécimen AMNH 5665, el frente de un esqueleto encontrado por Levi Sternberg en 1917. En 1930 Charles Whitney Gilmorer emitió ambos especímenes a Palaeoscincus rugosidens.[8] Esta especie se basó en el espécimen tipo USNM 11868, un esqueleto encontrado por George Fryer Sternberg en junio de 1928. El nombre específico se deriva del latín "rugosus", "rugoso", y dens , "diente". En 1940, Loris Shano Russell remitió los tres especímenes a Edmontonia , como Edmontonia rugosidens.[9]
Mientras tanto, la especie tipo de Edmontonia , Edmontonia longiceps, había sido nombrada por Charles Mortram Sternberg en 1928. El nombre genérico Edmontonia se refiere a Edmonton o la formación Edmonton. El nombre específico E. longiceps significa "cabeza dura" en latín. Su holotipo es el espécimen NMC 8531, que consiste en un cráneo, la mandíbula inferior derecha y gran parte del esqueleto postcraneal, incluida la armadura. Fue descubierto cerca de Morrin en 1924 por George Paterson, el jefe de la expedición liderada por C. M. Sternberg.[10]
Especies
- E. longiceps la especie tipo, conocido a partir de un cráneo completo, se conoce a partir de la parte media de la formación Cañón Herradura en la Unidad 2 que solía tener una fecha de hace 71,5 a 71 millones de años.[11] Esta unidad, que se extiende por el límite Campaniano-Maastrichtiano , ha sido recalibrada a una edad de aproximadamente 72 millones de años. Los huesos aislados y los dientes del cobertizo de E. longiceps también se conocen de la formación superior Judith River en Montana.
- E. rugosidens, al cual ocasionalmente se le da su propio género, Chassternbergia, acuñado por primera vez como un subgénero por el Dr. Robert Bakker en 1988 como Edmontonia (Chassternbergia) rugosidens, basado en las diferencias de proporciones con el cráneo de E. longiceps.[5][12] Este subgénero o género no es generalmente aceptado;[13][14] El nombre de Chassternbergia fue acuñado en honor al Dr. Charles Mortram "Chas" Sternberg (1885-1981), un paleontólogo canadiense que en 1928, nombrara y describiera a Edmontonia longiceps, un nodosáurido a que Robert Bakker usara como base para proponer una nueva familia Edmontoniidae, en la cual incluyó al subgénero, Chassternbergia y al nuevo género Denversaurus schlessmani. De esta manera Sternberg, fue honrado por su trabajo sobre Edmontonia longiceps.[15] E. rugosidens se encuentra en el Campaniense inferior formación parque de dinosaurios, que data de hace unos 76,5-75 millones de años.[11] Muchos hallazgos posteriores han sido referidos a E. rugosidens, entre ellos CMN 8879, la parte superior de un cráneo encontrado en 1937 por Harold D'acre Robinson Lowe, ROM 433, una espina bífida descubierta por Jack Horner en 1986 entre material Oohkotokia, ROM 5340, placas mediadas emparejadas; ROM 1215, un esqueleto, RTMP 91.36.507, una calavera, RTMP 98.74.1, un posible cráneo de Edmontonia, RTMP 98.71.1, un esqueleto,[16] RTMP 98.98.01, un cráneo y la mandíbula inferior derecha; y RTMP 2001.12.158, un cráneo.[4]
- E. australis,[12] Solo conocido por placas cervicales, siendo considerado dudoso[13] o sinónimo de Glyptodontopelta mimus.[17] Usualmente en este género se incluye a Denversaurus schlessmani.
- E. schlessmani era un cambio de nombre en 1992 de Denversaurus schlessmani, " lagarto de Denver deSchlessman" por Adrian Hunt y Spencer Lucas.[18] Este taxón fue erigido por Bakker en 1988 para un cráneo de la formación Lance del Cretácico Superior en el Maastrichtiense del sur de Dakota del Sur, muestra DMNH 468 encontrada por Philip Reinheimer en 1922. Este tipo de espécimen de Denversaurus está en las colecciones del Museo de Denver Historia Natural (ahora el Museo de la Naturaleza y la Ciencia de Denver), Denver, Colorado por lo que el género fue nombrado. El nombre específico honra a Lee E. Schlessman, cuya Fundación de la Familia Schlessman patrocinó el museo.[5] Bakker describió el cráneo como mucho más ancho en la parte trasera que los especímenes de Edmontonia.[5] Sin embargo, los trabajadores posteriores explicaron esto por su aplastamiento,[3] y consideraron al taxón como un sinónimo más moderno de Edmontonia longiceps.[14] El Instituto Black Hills ha referido un esqueleto de la Formación Lance a Denversaurus , apodado "Tanque". Tiene el número de inventario BHI 127327.[19] Nuevas investigaciones indican que está estrechamente relacionado con Panoplosaurus.[20]
Clasificación
C. M. Sternberg originalmente no proporcionó una clasificación de Edmontonia. . En 1930, L. S. Russell colocó el género en los Nodosauridae, lo que ha sido confirmado por análisis posteriores. En general se demostró que Edmontonia era un nodosaurido derivado, estrechamente relacionado con Panoplosaurus. Russell en 1940 nombró Edmontoniinae por separado . En 1988 Bakker propuso que los Edmontoniinae con los Panoplosaurinae se unieran a Edmontoniidae, el presunto grupo hermano de los Nodosauridae dentro de Nodosauroidea, que supuso que no serían anquilosaurianos sino los últimos estegosáuridos supervivientes.[5] Sin embargo, el análisis cladístico no ha confirmado estas hipótesis, y los conceptos de Edmontoniinae y Edmontoniidae no están en uso moderno.
Filogenia

El siguiente cladograma muestra la posición de Edmontonia en el árbol evolutivo nodosáurido de acuerdo con un análisis realizado en 2011 por los paleontólogos Richard Stephen Thompson , Jolyon C. Parish, Susannah CR Maidment y Paul M. Barrett.[21]
| Nodosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiología
Sus largas espinas eran probablemente usadas entre machos para demostraciones de fuerza para defender el territorio o conseguir pareja.[1] Las espinas pudieron usarse también para intimidar a sus depredadores o manadas rivales, protección o auto-defensa.[1] Para protegerse de los depredadores los Edmontonia pudieron agacharse en grupo para minimizar la posibilidad de ataque a su indefenso vientre, tratando de evitar ser volteados por un depredador. Los grandes picos de hombro que apuntan hacia delante podrían haberse utilizado para atacar a los terópodos.[2] Carpenter sugirió que los picos más grandes de AMNH 5665 indicaban que se trataba de un espécimen masculino, un caso de dimorfismo sexual. Sin embargo, admitió la posibilidad de que sean resultado de la ontogenia, los individuos mayores tenían espinas más largas, ya que el espécimen era relativamente grande.[3]
Paleoecología
Anillos en la madera petrificada de los árboles contemporáneos a Edmontonia muestran una fuerte estacionalidad de los cambios en precipitación y temperatura.[1] lo que puede tener una explicación de por qué tantos especímenes se han encontrado con su blindaje y espigas en la misma posición que en su vida. El Edmontonia podría haber muerto debido a la sequía, se secó y, a continuación, se cubren rápidamente en los sedimentos cuando la temporada de lluvias comenzó.[1]
Edmontonia rugosidens existió en la sección superior de la Formación Dinosaur Park, hace aproximadamente 76.5-75 millones de años. Vivió junto a otros numerosos herbívoros gigantes, como los hadrosáuridos Gryposaurus, Corythosaurus y Parasaurolophus, los ceratopsidos Centrosaurus y Chasmosaurus y los anquilosáuridos Scolosaurus y Dyoplosaurus.[11] Los estudios de la anatomía de la mandíbula y la mecánica de estos dinosaurios sugieren que probablemente todos estén ocupados nichos ecológicos ligeramente diferentes para evitar la competencia directa por la comida en un espacio ecológico tan concurrido.[22] Los únicos grandes depredadores conocidos de los mismos niveles de la formación con Edmontonia son los tiranosáuridos Gorgosaurus libratus y una especie anónima de Daspletosaurus.[11]
Edmontonia longiceps es conocida de la formación Horseshoe Canyon, de la unidad central, que data de 71.5-71 millones de años atrás en 2009.[11] La fauna de la Formación Horseshoe Canyon es bien conocida, como fósiles de vertebrados, incluidos los de los dinosaurios, son bastante comunes. tiburones, rayas, esturiones , Amiidae , Lepisosteidae y el Aspidorhynchus parecido a un Lepisosteidae componen la fauna de peces. El plesiosáurido de agua salada Leurospondylus ha sido encontrado en sedimentos marinos en el Cañón Horseshoe, mientras que los ambientes de agua dulce fueron poblados por tortugas, Champsosaurus y cocodrilos como Leidyosuchus y Stangerochampsa . Los dinosaurios dominan la fauna, especialmente los hadrosáuridos, que constituyen la mitad de todos los dinosaurios conocidos, incluidos los géneros Edmontosaurus, Saurolophus e Hypacrosaurus . Los ceratópsidos y ornitomímidos también eran muy comunes, y juntos constituían otro tercio de la fauna conocida. Los anquilosáuridos y paquicefalosáuridos mucho más raros , todos estos animales habrían sido presas de una gran variedad de terópodos carnívoros, incluidos troodóntidos, dromeosáuridos y canagnátidos.[23][24] El Albertosaurus adulto fue el depredador principal en este entorno, con nichos intermedios posiblemente llenos de albertosaurios juveniles.[23]
Referencias
- "Edmontonia." In: Dodson, Peter & Britt, Brooks & Carpenter, Kenneth & Forster, Catherine A. & Gillette, David D. & Norell, Mark A. & Olshevsky, George & Parrish, J. Michael & Weishampel, David B. The Age of Dinosaurs. Publications International, LTD. p. 141. ISBN 0-7853-0443-6.
- Paul, G.S., 2010, The Princeton Field Guide to Dinosaurs, Princeton University Press p. 238
- Carpenter, K. 1990. "Ankylosaur systematics: example using Panoplosaurus and Edmontonia (Ankylosauria: Nodosauridae)", In: Carpenter, K. & Currie, P.J. (eds) Dinosaur Systematics: Approaches and Perspectives, Cambridge University Press, Cambridge, pp. 281-298
- Vickaryous, Matthew K. (2006). «New information on the cranial anatomy of Edmontonia rugosidens Gilmore, a Late Cretaceous nodosaurid dinosaur from Dinosaur Provincial Park, Alberta». Journal of Vertebrate Paleontology 26 (4): 1011-1013. doi:10.1671/0272-4634(2006)26[1011:niotca]2.0.co;2.
- Bakker, R.T. (1988). Review of the Late Cretaceous nodosauroid Dinosauria: Denversaurus schlessmani, a new armor-plated dinosaur from the Latest Cretaceous of South Dakota, the last survivor of the nodosaurians, with comments on Stegosaur-Nodosaur relationships. Hunteria 1(3):1-23.(1988).
- Coombs, W. P Jr.; Maryańska, T. (1990). «Ankylosauria». En Weishampel, D. P.; Dodson, P.; Osmólka, H., eds. The Dinosauria. University of California Press. pp. 456-483.
- Matthew, W. D. (1922). «A superdreadnaught of the animal world—the armoured dinosaur Palaeoscincus». Natural History 22: 333-342.
- Gilmore, C.W. (1930). «On dinosaurian reptiles from the Two Medicine Formation of Montana». Proceedings of the United States National Museum 77 (16): 1-39. doi:10.5479/si.00963801.77-2839.1.
- Russell, L.S. (1940). «Edmontonia rugosidens (Gilmore), an armored dinosaur from the Belly River Series of Alberta». University of Toronto Studies, Geology Series 43: 3-28.
- Sternberg, C.M. (1928). «A new armored dinosaur from the Edmonton Formation of Alberta». Transactions of the Royal Society of Canada, series 3 22: 93-106.
- Arbour, V.M.; Burns, M. E.; Sissons, R. L. (2009). «A redescription of the ankylosaurid dinosaur Dyoplosaurus acutosquameus Parks, 1924 (Ornithischia: Ankylosauria) and a revision of the genus». Journal of Vertebrate Paleontology 29 (4): 1117-1135. doi:10.1671/039.029.0405.
- Ford, T.L. (2000). A review of ankylosaur osteoderms from New Mexico and a preliminary review of ankylosaur armor. In: Lucas, S.G., and Heckert, A.B. (eds.). Dinosaurs of New Mexico. New Mexico Museum of Natural History and Science Bulletin 17:157-176.
- Carpenter K (2001). «Phylogenetic analysis of the Ankylosauria». En Carpenter, Kenneth(ed), ed. The Armored Dinosaurs. Indiana University Press. pp. 455–484. ISBN 0-253-33964-2.
- Vickaryous, M.K., Maryańska, T., and Weishampel, D.B., (2004). «Ankylosauria». En Weishampel, D. B., Dodson, P., and Osmólska, H. (eds.), ed. The Dinosauria (Second Edition). University of California Press. pp. 363–392. ISBN 0-520-24209-2.
- Etimología Chassternbergia cortesía de www.dinosaurnames.net
- Currie P.J., Russell D.A., 2005, "The geographic and stratigraphic distribution of articulated and associated dinosaur remains", In: Currie P.J., Koppelhus E.B., (editors). Dinosaur Provincial Park: A spectacular ancient ecosystem revealed. Bloomington: Indiana University Press. pp 537–569
- Burns, Michael E. (2008). «Taxonomic utility of ankylosaur (Dinosauria, Ornithischia) osteoderms: Glyptodontopelta mimus Ford, 2000: a test case». Journal of Vertebrate Paleontology 28 (4): 1102-1109.
- Hunt, A.P. and Lucas, S.G., 1992, "Stratigraphy, Paleontology and age of the Fruitland and Kirkland Formations (Upper Cretaceous), San Juan Basin, New Mexico", New Mexico Geological Society Guidebook, 43rd Field Conference, San Juan Basin, volume 4, p. 217-240
- Carpenter, K.; DiCroce, T.; Kinneer, B.; Simon, R. (2013). «Pelvis of Gargoyleosaurus (Dinosauria: Ankylosauria) and the Origin and Evolution of the Ankylosaur Pelvis». PLoS ONE 8 (11): e79887. PMC 3828194. PMID 24244573. doi:10.1371/journal.pone.0079887.
- Burns, ME. Intraspecific Variation in Late Cretaceous Nodosaurids (Ankylosauria: Dinosauria). Journal of Vertebrate Paleontology, Program and Abstracts, 2015, 99–100. («Archived copy». Archivado desde el original el 6 de noviembre de 2015. Consultado el 30 de octubre de 2015.)
- Richard S. Thompson; Jolyon C. Parish; Susannah C. R. Maidment; Paul M. Barrett (2011). «Phylogeny of the ankylosaurian dinosaurs (Ornithischia: Thyreophora)». Journal of Systematic Palaeontology 10 (2): 301-312. doi:10.1080/14772019.2011.569091.
- Mallon, J. C., Evans, D. C., Ryan, M. J., & Anderson, J. S. (2012). Megaherbivorous dinosaur turnover in the Dinosaur Park Formation (upper Campanian) of Alberta, Canada. Palaeogeography, Palaeoclimatology, Palaeoecology.
- Eberth, D.A., 1997, "Edmonton group". In: Currie, P.J., Padian, K. (Eds.), Encyclopedia of Dinosaurs. Academic Press, New York, pp. 199–204
- Larson, Derek W.; Brinkman, Donald B.; Bell, Phil R. (2010). «Faunal assemblages from the upper Horseshoe Canyon Formation, an early Maastrichtian cool-climate assemblage from Alberta, with special reference to the Albertosaurus sarcophagus bonebed». Canadian Journal of Earth Sciences 47: 1159-1181. doi:10.1139/e10-005.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikimedia Commons alberga una categoría multimedia sobre Edmontonia.
Wikimedia Commons alberga una categoría multimedia sobre Edmontonia. Wikispecies tiene un artículo sobre Edmontonia.
Wikispecies tiene un artículo sobre Edmontonia.- Edmontonia en The DINOSAUR ENCYCLOPAEDIA Version 4.0 (en inglés)
- Edmontonia dentro de Nodosauridae en The Thescelsosaurus (en inglés)
- Significado y pronunciación del nombre (en inglés)
| Control de autoridades |
|
|---|
 Datos: Q245262
Datos: Q245262- Multimedia: Edmontonia / Q245262