Ornithischia
Los ornitisquios (Ornithischia, en griego "caderas de ave"), antiguamente Predentata, son uno de los dos órdenes de dinosaurios que vivieron desde el Triásico Superior en el Carniense y el Cretácico superior en el Maastrichtiense, hace aproximadamente entre 228 y 66 millones de años, y tuvieron una distribución mundial. El nombre proviene del griego, estando formado por las palabras ornitheos (ορνιθειος) que significa ave y ischion (ισχιον) por cadera, siendo conocidos como los dinosaurios de cadera de ave. Esta similitud solo es superficial, ya que las aves derivan de un grupo de saurisquios, el otro orden de dinosaurios.
| Ornithischia | ||
|---|---|---|
|
Rango temporal: 200 Ma - 66 Ma Jurásico Inferior - Cretácico Superior (posible registro del Triásico Superior) | ||
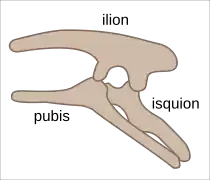 La pelvis de un ornitisquio. | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: |
Ornithischia Seeley, 1888 | |
| Subórdenes | ||
Siendo herbívoros, vivieron a veces en manadas, siendo más numerosa que los saurisquios. Los ornitisquios eran las presas para los terópodos y por lo general más pequeños que saurópodos. Muchos tipos desarrollaron características que hoy en día vemos en los mamíferos ungulados como las elaboradas baterías de dientes para masticar, y los cuernos y otros tipos de crestas y de ornamentos, usados en luchas entre congéneres más que contra depredadores.
Descripción
Los dinosaurios fueron divididos en dos grandes grupos por Harry Seeley en 1887, basándose en la forma de sus caderas. Así estaban los ornitisquios con la rama del pubis orientada en dirección caudal, como en las aves, y los saurisquios, en los que la rama del pubis tenía una orientación craneal, como en los reptiles. No obstante, el parecido es superficial, ya que las propias aves descienden de los dinosaurios saurisquios y no de los ornitisquios. Además poseen detalles característicos en las vértebras y la presencia de una posible armadura, aunque esto fue compartido con los saltasáuridos. Por otro lado en el frente de la mandíbula inferior, a partir del dentario se extendía el hueso predentario que junto con el premaxilar formaban un pico para cortar las plantas.

En los ornitisquios el pubis señala hacia abajo y hacia atrás (hacia la cola), paralelo al isquion, con un proceso prepúbico delantero para apoyar el abdomen. Esto hace una estructura pélvica de cuatro extremos, y una pelvis más ancha y estable. En contraste con esto, los extremos del pubis en los saurisquios eran hacia abajo y hacia la cabeza (hacia adelante), como en los lagartos anteriores. El arreglo del pubis, paralelo a la columna vertebral, como en las aves, se desarrolló independientemente tres veces durante la evolución de los dinosaurios, en los ornitisquios, tericinosáuridos y en los dromeosáuridos, estos últimos hoy son considerados los más probables ancestros de las aves. Los ornitisquios también tenían agujeros más pequeños por delante de sus órbitas oculares (ventanas anteorbitales) con el hueso palpebral en la órbita.
Otras características de los ornitisquios son dientes malares con las coronas bajas, subtriangular; dientes foliares con las raíces estrechas; 3 a 5 vértebras sacras; tendones osificados sobre el sacro; el quinto dedo del pie reducido sin ninguna falange. Todos tenían mejillas, picos córneos y eran herbívoros. Gran variedad de ornamentación defensiva.
Historia
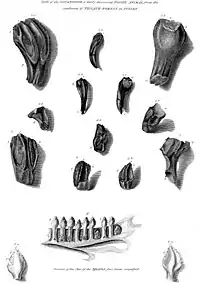
Un ornitisquio tiene el honor de ser el primer dinosaurio descrito: en 1822 el geólogo inglés Gideon Mantell, describía a partir de unos dientes encontrados en una cantera de Inglaterra a Iguanodon.[1][2] Luego en 1832 el mismo Mantell, describió al primer tireofóro, Hylaeosaurus .[3] En 1858 se encontraba el primer dinosaurio en Norteamérica, Hadrosaurus, un ornitópodo. Fue un descubrimiento sumamente importante: el primer esqueleto de dinosaurio encontrado casi completo, en el cual se evidenciaba una postura claramente bípeda.[4] Un año después, 1859, se encontraba a Scelidosaurus.[5] En 1878 en la mina Bernissart de Bélgica se encontraba un gran grupo de iguanodontes, con varios géneros mezclados y que fueron ensamblados en una posición similar a un canguro por Louis Dollo.[6]

El primer Hypsilophodon fue descubierto en 1849.[7][8] Sin embargo, originalmente se creyó que pertenecía a un ejemplar joven de iguanodonte. No fue sino hasta 1870 que el paleontólogo Thomas H. Huxley publicó la descripción completa del nuevo género. En 1882, algunos paleontólogos sugirieron que como un moderno canguro arborícola, era capaz de trepar árboles para buscar refugio. Esta postura fue aceptada por casi un siglo, hasta que Peter M. Galton finalmente realizó en 1971 un análisis detallado de la estructura esqueleto-muscular y convenció a la mayoría de sus colegas de la imposibilidad de tal comportamiento.[9][10]
El término "Ornithischia" fue acuñado por Harry Seeley en 1887 pero recién fue publicado en 1888 para agrupar los dinosaurios con cadera semejante a aves.[11] En 1894 Othniel Charles Marsh acuñó el nombre Predentata para los mismos, que fuera muy usado por todo el siglo XX, hasta que Robert Bakker propusiera reemplazarlo por el más antiguo Ornithischia en 1986.[12]
En 1877 Marsh describió a Stegosaurus,[13] y en 1888 encontraba a Ceratops en Norteamérica y descrito, que aunque incompleto es el primer ceratopsiano conocido.[14] En el mismo año también se encontró a Triceratops.[15]
En la expedición alemana de 1909 a 1912 a Tendaguru en Tanzania se encontraba a Kentrosaurus[16] y a Dryosaurus, este último ya conocido de la Formación Morrison en el oeste de Estados Unidos. Esto llevó a la idea que ambas masas de tierras estaban unidas en el Jurásico.
En la expedición al Desierto de Gobi de 1922 para buscar los ancestros humanos el fotógrafo J.B. Shackelford descubrió el primer espécimen de Protoceratops, uno de los dinosaurios con mayor registro fósil. Fue encontrado cerca de una nidada de alrededor de 30 huevos, que se asignaron a Protoceratops, pero recientes estudios han demostrado que estos pertenecían al terópodo Oviraptor.[17]

En Sudamérica se habían encontrado pequeños restos asignados a ornitisquios como Notoceratops[18] en 1918 por Tapia o Secernosaurus[19] en 1973, pero fue Secernosaurus, encontrado por José Fernando Bonaparte en 1983 el que brindó la información sobre la comunicación de las faunas sureñas y norteñas a finales del Cretácico.[20] En 1996 Rodolfo Coria y Leonardo Salgado, propusieron un nuevo clado, Euiguanodontia dentro de Ornithopoda para incluir a un grupo de ornitisquios sudamericanos que se diferenciaban del resto a partir del descubrimiento de Gasparinisaura.[21]
En 1989 Leaellynasaura fue encontrado en el sur de Australia. El hecho de que viviera en temperaturas extremadamente bajas lleva a muchos científicos a creer que este animal pudo haber sido endotérmico.[22]
En 2006 se descubrió el primer dinosaurio antártico, un tireofóro al que Leonardo Salgado y Zulma Gasparini llamaron Antarctopelta.[23]
Clasificación
Se define a Ornithischia como el clado más inclusivo que contiene a Triceratops horridus (Marsh, 1889) pero no a Passer domesticus (Linnaeus, 1758) ni a Saltasaurus loricatus (Bonaparte & Powell, 1980).
Taxonomía

- Orden Ornithischia
- Pisanosaurus
- Lesothosaurus
- Clado Genasauria
- Suborden Thyreophora
- Tatisaurus?
- Scutellosaurus
- Clado Eurypoda
- Brachypodosaurus?
- Lusitanosaurus?
- Infraorden Stegosauria
- Infraorden Ankylosauria
- Suborden Neornithischia
- Stormbergia
- Agilisaurus
- Hexinlusaurus
- Infraorden Ornithopoda
- Clado Heterodontosauriformes
- Familia Heterodontosauridae
- Clado Marginocephalia
- Infraorden Pachycephalosauria
- Infraorden Ceratopsia
- Suborden Thyreophora
Filogenia
Ornithischia se divide en dos clados principales, Thyreophora y Cerapoda. Thyreophora incluye a Stegosauria (como Stegosaurus) y Ankylosauria (como Ankylosaurus). Cerapoda incluye a Marginocephalia que a su vez se divide entre los Ceratopsia (como Triceratops) y Pachycephalosauria (como Stegoceras) y a Ornithopoda (como Edmontosaurus). Cerapoda es una agrupación relativamente reciente (Sereno, 1986), y puede ser idéntica con el más nuevo grupo Neornithischia. La mayor parte de estas divisiones no son verdades por definición.
En la base de Ornithischia, como la base de muchos otros grandes grupos, hay un gran grupo de especímenes poco conocidos que son imposibles de clasificar con mucha exactitud. En el caso de Ornithischia, conocen a este grupo colectivamente como los Fabrosauridae, mientras que los más avanzados son agrupados en el clado Genasauria, que incluye a los tireofóros y neornitisquios.
El siguiente cladograma se debe a Butler et al., 2011.[24]
| Ornithischia |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Historia natural

El ornitisquio más antiguo que se conoce es Pisanosaurus, contemporáneo a Eoraptor y Herrerasaurus, todos del Carniense, Triásico superior, en Ischigualasto, noroeste de la República Argentina.[25] Existe gran controversia sobre este dinosaurio, si bien es considerado un ornitisquio primitivo e incluido dentro de los fabrosáuridos,[26] hay quien los considera un heterodontosáurido y otros no piensan que fuese un ornitisquio.[27]
Los fabrosáuridos típicos provienen de principios del Jurásico en África, como Lesothosaurus y Fabrosaurus aunque estos dos son considerados sinónimos por muchos. Se lo considera menos primitivo que Pisanosaurus y Eocursor, pero retrasado filogenéticamente respecto a Heterodontosaurus.[28] En el 2005, Richard J. Butler publicó un estudio filogenético de ornitisquios, en el cual propuso que el lesotosaurio fue un miembro basal del clado Neornithischia, el cual incluye paquicefalosaurianos, ceratopsianos y ornitópodos.[29]

El tireóforo más antiguo que se conoce bien es Scutellosaurus[30] de Norteamérica un pariente cercano de Scelidosaurus, aunque este último es cuadrúpedo y el primero es bípedo.[31] Los tireofóros más derivados son agrupados en el clado Eurypoda. Este incluye a los estegosaurianos y a los anquilosaurianos.[28]
Los primeros estegosaurianos provienen del Jurásico medio en China e Inglaterra, siendo el mejor conocido Huayangosaurus.[32] La mayoría de los estegosaurianos conocidos son los estegosáuridos, de finales del Jurásico y principios del Cretácico, entre ellos se encuentran Stegosaurus de Norteamérica, Lexovisaurus de Europa, Tuojiangosaurus de Asia y Kentrosaurus de África.[33]
Los primeros anquilosaurianos conocidos fueron los nodosáuridos, que aparecieron a mediados del Jurásico en Europa (Sarcolestes) y se distribuyeron por Asia y Norteamérica entre principios y mediados del Cretácico (Nodosaurus, Zhongyuansaurus y Acanthopholis).[34]

El otro grupo de anquilosaurianos, los anquilosáuridos se conocen desde el Jurásico superior (Gargoyleosaurus)[35] a finales del Cretácico (Ankylosaurus),[36] en Norteamérica (Euoplocephalus), Asia (Talarurus), Europa (Polacanthus). Los primeros en aparecer fueron los polacantinos a finales del Jurásico hasta mediados del Cretácico (Hoplitosaurus), luego los shamosaurinos de mediados del Cretácico en Asia (Shamosaurus) y los más derivado y últimos en aparecer fueron los anquilosaurinos de mediados del Cretácico hasta la extinción de los dinosaurios (Tarchia)[34]
Los Neornithischia, como Agilisaurus, incluyen a los heterodontosauriformes y a los ornitópodos.[26][27] La primera rama de los heterodontosauriformes son los heterodontosáuridos, bien conocidos en principios del Jurásico en África (Heterodontosaurus) y también conocidos por restos fragmentarios en Argentina, México y Estados Unidos.[37][38] Hay un heterodontosáurido sin describir de finales del Jurásico en la Formación Morrison y el último parece ser Echinodon de principios del Cretácico en Inglaterra.[39]

De estos derivaron en los marginocéfalos,[40] que se dividen en dos, por un lado los paquicefalosaurianos cuyos ejemplares más basales son Ferganocephale, del Jurásico medio de Asia y Stenopelix del Cretácico inferior de Europa.[28] De estos a finales del Cretácico aparecieron los paquicefalosáuridos como Stegoceras Norteamérica y Tylocephale de Asia.[41]
El otro gran grupo son los ceratopsianos,[42] cuyo miembro más antiguo es Yinlong del Jurásico medio de Asia.[43] Luego a principios del Cretácico aparecen los psitacosáuridos como Psittacosaurus,[44] para posteriormente aparecer los neoceratopsianos cuyo miembros más basales del Cretácico inferior como Archaeoceratops.[28] Más avanzados que estos son los coronasaurianos como Protoceratops, todas formas asiáticas.[28] Los ceratópsidos propiamente dichos de finales del Cretácico son las formas más avanzadas de mayor tamaño y norteamericanas como Triceratops estos se dividen en dos, los centrosaurinos y los ceratopsianos.[45]
Los ornitopódos más primitivos al parecer derivan directamente de los fabrosáuridos,[26] siendo Hexinlusaurus un nexo entre estos y los más derivados hipsilofodóntidos.[46] Este último es un heterogéneo grupo de pequeños bípedos corredores que se usa para englobar a los más primitivos ornitopódos. El más antiguo conocido es Gongbusaurus,[47] del Jurásico medio en Asia, estos existieron hasta finales del Cretácico como el derivado Thescelosaurus.[48]

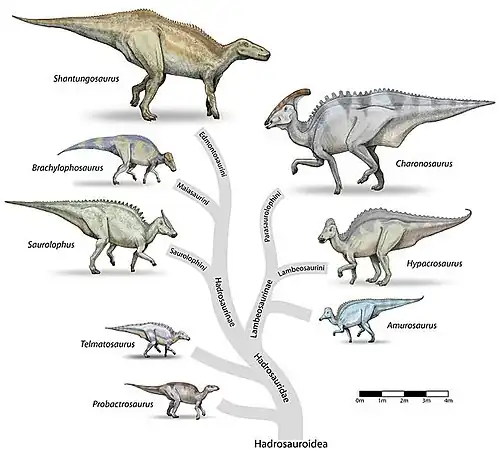
A finales del Jurásico, a partir de un grupo de hipsilofodóntidos hacen su aparición los iguanodontianos como Dryosaurus y Camptosaurus de Norteamérica, África y Europa.[49] De estos derivan los más avanzados ornitópodos, los hadrosauriformes en el Cretácico inferior, que incluyen a Iguanodon.[50] A finales del Cretácico hacen aparición los hadrosáuridos propiamente dicho como Parasaurolophus y Edmontosaurus de Norteamérica, Secernosaurus de Suramérica, Shantungosaurus y Charonosaurus asiáticos y Telmatosaurus de Europa. Divididos en saurolofinos y lambeosaurinos.[51]
En Sudamérica, a partir de ornitópodos primitivos evolucionaron los euiguanodóntidos[21] a finales del Cretácico, como Talenkauen.[52]
Referencias
- Sues, Hans-Dieter (1997). «European Dinosaur Hunters». En James Orville Farlow and M. K. Brett-Surman (eds.), ed. The Complete Dinosaur. Bloomington: Indiana University Press. pp. 14. ISBN 0-253-33349-0.
- Lucas, Spencer G. (diciembre de 1999). «Book review: Gideon Mantell and the discovery of dinosaurs». PALAIOS 14 (6): 601-602. ISSN 0883-1351. doi:10.2307/3515316. Consultado el 21 de febrero de 2007.
- Mantell, F. R. S. 1833. The Geology of the Sout-East of England. Saurians, and chapter X, observations on the fossil remains of the Hylaeosaurus, and other saurian reptiles discovered in the Strata of Tilgate Forest, in Sussex. 260-233.
- Leidy, J. (1858). «Proceedings of The Academy of Natural Sciences of Philadelphia» 10: 215-218.
- Newman, B.H. (1968) The Jurassic dinosaur Scelidosaurus harrisoni, Owen. Palaeontology 11 (1), 40-3.
- Norman, David B. (marzo de 1987). «A mass-accumulation of vertebrates from the Lower Cretaceous of Nehden (Sauerland), West Germany». Proceedings of the Royal Society of London. Series B, Biological Sciences 230 (1259): 215-255. doi:10.1098/rspb.1987.0017.
- Huxley, T. H. 1869. On Hypsilophodon, a new genus of Dinosauria. Proceedings of the Geological Society of London, 204, 3-4.
- Owen, R. 1873. Discussion on Hypsilophodon foxii. Quarterly Journal of the Geological Society of London, 24, 531 - 532.
- Galton, P.M.1971. Hypsilophodon, the cursorial non-arboreal dinosaur. Nature, 231, 159-161.
- Galton, P. M. 1971. The mode of life of Hypsilophodon, the supposedly arboreal ornithopod dinosaur. Lethaia, 4, 453-465.
- Seeley, H. G. 1887-1888. On the classification of the fossil animals commonly called Dinosauria. Proceedings of the Royal Society London, XLIII, 165-171.
- Marsh, O. 1896. The Dinosaurs of North America. United States Geological Survey 16th Annual Report. 1894-95,133-414.
- Marsh, O. C. 1877. New order of extinct Reptilia (Stegosauria) from the Jurassic of the Rocky Mountains. American Journal of Science (ser. 3),14, 513-514.
- Marsh, O. C. 1889. Notice of new American dinosaurs. American Journal of Science (ser. 3),37, 331-336.
- Marsh, O. C.1889. Notice of gigantic horned Dinosauria from the Cretaceous. American Journal Science, 138, 173-175.
- Nopcsa, F. 1916. Doryphorosaurus nov. nom. für Kentrosaurus Hennig. Cbl. Mineral. Geol. Palaeontol., 1916, 511-512.
- Granger, W. & Gregory, W. K. 1923. Protoceratops andrewsi, a pre-ceratopsian dinosaur from Mongolia. American Museum Novitates, 72, 1-9.
- Dodson, Peter; The Horned Dinosaurs (1996)
- Brett-Surman, M. K. 1979. Phylogeny and Paleobiogeography of Hadrosaurian Dinosaurs. Nature, 277, 15, 560-562.
- Bonaparte, J. F., Franchi, M. R., Powell, J. E. y Sepúlveda, E. G. 1984. La Formación Alamitos (Campaniaense-Maastrichtiense) del sudeste del Río Negro, con descripción de Kritosaurus australis n. sp. (Hadrosauridae). Significado paleogeográfico de los vertebrados. Revista de la Asociación Geológica Argentina, 39, 284-299.
- Coria, R. A., and L. Salgado. 1996. A basal iguanodontian (Ornithischia: Ornithopoda) from the Late Cretaceous of South America. Journal of Vertebrate Paleontology 16:445–457
- Rich, T. H. V. & Rich, P. V.1989. Polar dinosaurs and biotas of Early Cretaceous of Southeastern Australia. National Geographic Research, 5, 1, 15-53.
- Gasparini, Z., Olivero, E., Scasso, R., & Rinaldi, C. 1996. New data on the ankylosaurian dinosaur from the Late Cretaceous of the Antarctic Peninsula. Memoirs of the Queensland Museum 39: 583-594.
- Richard J. Butler, Jin Liyong, Chen Jun, Pascal Godefroit (May 2011). «The postcranial osteology and phylogenetic position of the small ornithischian dinosaur Changchunsaurus parvus from the Quantou Formation (Cretaceous: Aptian–Cenomanian) of Jilin Province, north-eastern China». Palaeontology 54 (3): 667-683. doi:10.1111/j.1475-4983.2011.01046.x.
- R. M. Casamiquela, 1967. Un nuevo dinosaurio ornitisquio triásico (Pisanosaurus mertii; Ornithopoda) de la Formación Ischigualasto, Argentina. Ameghiniana 4(2):47-64
- Sereno, P. C. 1991. Lesothosaurus, ""fabrosaurids"", and the early evolution of Ornithischia. Journal of Vertebrate Paleontology, 11, 168-197.
- Heckert, A. B. & Lucas, S. G.1999.Late Triassic orntihischian dinosaur evolution. Journal of Vertebrate Paleontology, Abstracts of papers, fifty-ninth annual meeting society of vertebrate paleontology, 20-23 October.19, Suppl. 3, 50A.
- Sereno, P.C. 1986. Phylogeny of the bird-hipped dinosaurs (order Ornithischia). National Geographic Research 2(2):234-256.
- Butler, R.J. 2005. The 'fabrosaurid' ornithischian dinosaurs of the Upper Elliot Formation (Lower Jurassic) of South Africa and Lesotho. Zoological Journal of the Linnean Society 145(2):175-218.
- Rosenbaum, J. & Padian, K. 2000. New material of the basal thyreophoran Scutellosaurus lawleri from the Kayenta Formation (Lower Jurassic) of Arizona. PaleoBios, 20, 1, 13-23.
- Martill, D. M., Batten, D. J. & Loydell, D. K. 2000. A new specimen of the thyreophoran dinosaur cf. Scelidosaurus with soft tissue preservation. Palaeontology, 43, 3, 549-559.
- Sereno, P. C. & Dong, Z.-M. 1992. The skull of the basal stegosaur Huayangosaurus taibaii and a cladistic diagnosis of Stegosauria. Journal of Vertebrate Paleontology, 12, 3, 318-343.
- Galton, P. M., 1990: Stegosauria. 435-455, in Weishampel, D. B., Dodson, P., & Osmólska, H. (eds.), The Dinosauria University of California Press, Berkley, Los Angeles, Oxford, 1990.
- Carpenter K (2001). "Phylogenetic analysis of the Ankylosauria", in Carpenter, Kenneth(ed): The Armored Dinosaurs. Indiana University Press, 455–484. ISBN 0-253-33964-2.
- Hill, R. V. 1999. Phylogenetic relationships among Ankylosauria: an analysis of cranial characters. Journal of Vertebrate Paleontology, Abstracts of papers, fifty-ninth annual meeting society of vertebrate paleontology, 20-23 October, 19, Suppl. 3, 51A.
- Brown, B. 1908. The Ankylosauridae, a new family of armored dinosaurs from the Upper Cretaceous. Bulletin of the American Museum of Natural History.24, 187-201.
- Weishampel, David B.; Witmer, Lawrence M. (1990). «Heterodontosauridae». En Weishampel, David B.; Osmólska, Halszka.; and Dodson, Peter. (eds.)., ed. The Dinosauria. Berkeley: University of California Press. pp. 486-497. ISBN 0-520-06727-4.
- Norman, David B.; Sues, Hans-Dieter; Witmer, Lawrence M.; Coria, Rodolfo A. (2004). «Basal Ornithopoda». En Weishampel, David B.; Osmólska, Halszka.; and Dodson, Peter. (eds.)., ed. The Dinosauria (2nd edition edición). Berkeley: University of California Press. pp. 393-412. ISBN 0-520-24209-2.
- Norman, D.B. & P.M. Barrett, 2002. Ornithischian dinosaurs from the Lower Cretaceous (Berriasian) of England. Special Papers in Palaeontology 68: 161-189.
- Dodson, P. 1990. Marginocephalia. In Dinosauria, Ed. D.B. Weishampel, P. Dodson, H. Osmólska. University of California Press, Ed. D.B. Weishampel, P. Dodson, H. Osmólska. University of California Press, 562- 563.
- Sues, H.-D., and P. M. Galton. (1987). "Anatomy and classification of the North American Pachycephalosauria (Dinosauria: Ornithischia)". Palaeontographica Abteilung A 198: 1–40
- Marsh, O.C. (1890). "Additional characters of the Ceratopsidae, with notice of new Cretaceous dinosaurs." American Journal of Science, 39: 418-429.
- Xu X., Forster, C.A., Clark, J.M., & Mo J. 2006. "A basal ceratopsian with transitional features from the Late Jurassic of northwestern China." Proceedings of the Royal Society B: Biological Sciences. doi:10.1098/rspb.2006.3566 [published online]
- Sereno, P. C. (1990). Psittacosauria. 579-592 in Weishampel, D. B., Dodson, P., & Osmólska, H. (eds.), 1990: The Dinosauria. University of California Press, Berkley, Los Angeles, Oxford, 1990 xvi-733
- Dodson, P. (1996). The Horned Dinosaurs. Princeton University Press, Princeton, New Jersey, pp. xiv-346
- Sues, Hans-Dieter; and Norman, David B. (1990). "Hypsilophodontidae, Tenontosaurus, Dryosauridae", in Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.): The Dinosauria, 1st, Berkeley: University of California Press, 498-509. ISBN 0-520-06727-4.
- Dong Zhiming (1989). "On a small ornithopod (Gongbusaurus wucaiwanensis sp. nov.) from Kelamaili, Junggar Basin, Xinjiang, China". Vertebrata PalAsiatica 27 (2): 140-146.
- Gilmore, Charles M. 1913. "A new dinosaur from the Lance Formation of Wyoming." Smithsonian Miscellaneous Collections 61(5): 1-5.
- Norman, David B. (2004). "Basal Iguanodontia", in Weishampel, D.B., Osmólska, H., and Dodson, P. (eds.): The Dinosauria. Berkeley: University of California Press, pp. 413-437. ISBN 0-520-24209-2.
- Norman, David B.; Weishampel, David B. (1990). "Iguanodontidae and related ornithopods", in Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.).: The Dinosauria. Berkeley: University of California Press, 510–533. ISBN 0-520-06727-4.
- Horner, John R.; Weishampel, David B.; and Forster, Catherine A (2004). "Hadrosauridae", in Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.): The Dinosauria, 2nd, Berkeley: University of California Press, 438-463. ISBN 0-520-24209-2.
- Novas, Fernando E.; Cambiaso, Andrea V; Ambrioso, Alfredo (2004). «A new basal iguanodontian (Dinosauria, Ornithischia) from the Upper Cretaceous of Patagonia». Ameghiniana 41 (1): 75-82.
Enlaces externos
 Wikimedia Commons alberga una galería multimedia sobre Ornithischia.
Wikimedia Commons alberga una galería multimedia sobre Ornithischia. Wikispecies tiene un artículo sobre Ornithischia.
Wikispecies tiene un artículo sobre Ornithischia.- Ornithischia en Paleos (en inglés)
- Ornithischia en The Thescelsosaurus (en inglés)
- Ornithischia en The taxonsearch.org
| Control de autoridades |
|
|---|
 Datos: Q191488
Datos: Q191488- Multimedia: Ornithischia / Q191488
- Especies: Ornithischia