Talarurus plicatospineus
Talarurus plicatospineus es la única especie conocida del género extinto Talarurus (gr. "cola en cesta") de género de dinosaurio tireofóro anquilosáurido, que vivió a mediados del período Cretácico, hace aproximadamente entre 96 a 89 millones de años, durante el Turoniense, en lo que es hoy Asia.
| Talarurus plicatospineus | ||
|---|---|---|
| Rango temporal: 96 Ma - 89 Ma Cretácico superior | ||
 Restauración en vida. | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Ornithischia | |
| Suborden: | Thyreophora | |
| Infraorden: | Ankylosauria | |
| Familia: | Ankylosauridae | |
| Subfamilia: | Ankylosaurinae | |
| Género: | Talarurus | |
| Especie: |
T. plicatospineus Maleev, 1952 | |
Los primeros restos de Talarurus fueron descubiertos en 1948 y posteriormente descritos por el paleontólogo ruso Evgeny Maleev, llamando a la especie tipo T. plicatospineus. Se conoce a partir de múltiples pero escasos especímenes, lo que lo convierte en uno de los anquilosáuridos más conocidos, junto con Pinacosaurus. Los elementos de los especímenes consisten en varios huesos del cuerpo. Se han descubierto y asignado al género cinco cráneos, aunque los dos primeros estaban muy fragmentados.
Era un herbívoro cuadrúpedo de tamaño mediano, corpulento, que vivía en el suelo , que podía crecer hasta 5 metros de largo y pesaba alrededor de 454 a 907 kilogramos, entre media y una tonelada. Al igual que otros anquilosáuridos, tenía una armadura pesada y un garrote en la cola, lo que limitaba su velocidad. Talarurus se clasifica como miembro de Ankylosauria, en Ankylosaurinae, un grupo de anquiosaurianos derivados. Talarurus se conoce de la Formación Bayan Shireh, siendo probablemente un nicho dividido con Tsagantegia, como lo indica su hocico, que tiene una forma rectangular especializada para el pastoreo. Estos representan los anquilosauridos más antiguos conocidos de Asia, aunque no están muy relacionados entre sí. Parece que el pariente más cercano de Talarurus era Nodocephalosaurus , un anquilosáurido con osteodermos faciales similares de América del Norte.
La mayoría de las monturas esqueléticas de Talarurus están desactualizadas por numerosos problemas, como que las costillas apuntan hacia abajo en lugar de hacia los lados, como en la mayoría de los anquilosáuridos. Un molde de cráneo inexacto que se basa principalmente en especies relacionadas y no en el material de cráneo disponible. Patas delanteras y traceras muy abiertos. Cuatro dedos en los pies, que en realidad solo tenían tres como lo indican los anquilosáuridos relacionados. Todos estos errores fueron cometidos por malentendidos anatómicos ya que los anquilosáuridos no se conocían completamente en ese momento, además, Talarurus no se conoce de un esqueleto completo.
Descripción
Inicialmente, Maleev describió a Talarurus con cuatro dedos en el pie.[1] Sin embargo, el pie no se encontró en articulación. El pie montado es un compuesto, y tres es el número más probable, ya que todos los demás anquilosáuridos conocidos muestran tres dedos. Informes anteriores de que Pinacosaurus también poseía cuatro son incorrectos. Otra supuesta característica, los osteodermos tenían una ornamentación surcada, formando una armadura especialmente formidable, con cada placa adornada con espinas plisadas adicionales. Estos también se basaron en un malentendido. Eran segmentos de los medios anillos que protegían el cuello, con sus típicas quillas bajas. El montaje tiene la particularidad adicional de que muestra a Talarurus como construido como un hipopótamo, con un tórax en forma de barril, no con el tipo de cuerpo bajo y ancho característico de los anquilosáuridos, y las extremidades anteriores están fuertemente separadas. Esto fue causado por una posición incorrecta de las costillas como si estuvieran unidas en lugar de sobresalir hacia los lados; este error también impidió el montaje de los elementos pélvicos superiores anchos.[2][3]

Talarurus era un anquilosaurio de tamaño mediano, Thomas Holtz y Gregory S. Paul estimaron su longitud en 5 metros, Paul dio un peso de 2 toneladas,[4] sin embargo, Holtz lo estimó alrededor de 454 a 907 kilogramos.[5] Al igual que otros anquilosáuridos, tenía un amplio pico, formidables osteodermos que recorren su cuerpo, formando una extensa armadura, y el famoso garrote de la cola . Sus extremidades eran robustas y cortas, soportando un vientre ancho y redondeado.[1][6] Los diagnósticos proporcionados por Maleev 1956 y Tumanova 1987 fueron de utilidad limitada ya que enumeraban en gran medida rasgos compartidos con muchos otros anquilosáuridos. Arbour también notó que el pie en realidad tenía tres dedos. Estableció una autapomorfía única, en los frontales, en el techo medio del cráneo, está presente una región elevada en forma de V. También determinó que Talarurus se diferenciaba de todos los anquilosáuridos conocidos con la excepción del norteamericano Nodocephalosaurus en la posesión de caputegulas, osteodermos o placas de armadura faciales, en los frontales y nasales, que tienen forma de cono con base circular.[2] H Sin embargo, según Parks et al. en 2019, Talarurus se puede distinguir en función de diversas características rostrales, como la caputegula internaria prominente, las caputegulas alargadas, las numerosas caputegulas que rodean el área nasal, las caputegulas de forma cónica/poligonal, entre otras.[6]
Cráneo
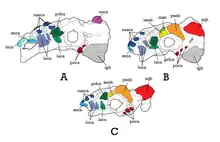
El cráneo de Talarurus medía unos 30 centímetros de longitud total.[6] El holotipo, PIN 557-91, se basa únicamente en un techo posterior del cráneo,[7][6] que presenta caputegulas similares a tubérculos que se asemejan a los de Nodocephalosaurus . Como señalaron Arbor y Currie, el techo del cráneo de PIN 3780/1 parece tener caputegulas ligeramente diferentes y no estaba claro si esto representa una diferencia taxonómica.[3] Sin embargo, todos los cráneos referidos comparten los mismos patrones de caputegulas (aunque se observa poca variación entre los individuos y por lo tanto son atribuibles al género.[6] El cráneo más completo es el MPC-D 100/1354. Conserva muy bien el rostro maxilar, el cual es ancho, de forma semi-rectangular y fusionado, no se encuentran dientes en esta zona; probablemente útil en superficies planas. Como era de esperar, las caputegulae tienen forma de polígono a tubérculo, y algunas están presentes en el área nasal. Cada maxilar conserva 23 alvéolos, sin embargo, los dientes no se conservaron. La mayoría de las caputegulas presentes en la parte superior de la cabeza son de forma hexagonal, en el holotipo PIN 557-91 algunas son triangulares. Los orbitales están fuertemente blindados con algunos osteodermos detrás de ellos. En MPC-D 100/1355 y MPC-D 100/1356 los cuernos escamoso y cuadratoyugal son gruesos y triangulares, siendo los escamosos más redondeados. Por lo general, los restos de la mandíbula inferior de Talarurus no se conservan, sin embargo, MPC-D 100/1355 se encontró en asociación con un dentario derecho , aunque muy fragmentado.[6]
Esqueleto postcraneal
La mayoría de los restos poscraneales se conocen de individuos fragmentarios. Según los especímenes PIN 557, las nervaduras están fuertemente curvadas y engrosadas, con una longitud que oscila entre 50 y 100 cm. Falta parcialmente la zona vertebral, conservando algunas cervicales, dorsales, caudales y el sacro. Las vértebras cervicales son anficelosas, cóncavas en ambos lados, con centros engrosados, los arcos neurales son ligeramente más altos. Las dorsales tienen el centro alto y ancho, y también son anficelosas con arcos neurales muy alargados y de constitución robusta, algunas están rotas. Más completo que los apartados anteriores, el sacro consta de 4 presacras, 4 sacros y la primera vértebra caudal, estas vértebras están fusionadas, con algunas costillas unidas. Su tamaño aumenta gradualmente de atrás hacia adelante. Algunas vértebras caudales aisladas están presentes, parecen indicar que la cola constaba de aproximadamente 25 a 30 caudales. Las caudales más anteriores tienen arcos neurales y centros algo alargados, los cheurones están fusionados y tienen forma de triángulo. Por el contrario, las caudales más posteriores tienen cheurones y arcos neurales muy aplanados.[1][8] Debido a la conservación parcial de las muestras,se desconoce el arreglo exacto el osteodermos, sin embargo, algunos han sido desenterrados. Los osteodermos conservados son muy escasos y consisten en algunos medios anillos cervicales y osteodermos corporales en forma de columna. Los osteodermos son fuertemente robustos, la mayoría de los osteodermos en forma de columna miden 15 cm de altura.[1][8] MPC-D 100/1355 se encontró en asociación con un medio anillo parcial, así como con otros elementos del cuerpo.[6]

Sólo se conserva la mano izquierda , está prácticamente completa conservando cinco dígitos, sólo faltan algunos ungueales y falanges. El pie izquierdo conservado es muy singular, se describió que tenía cuatro dígitos,[1][8] sin embargo, se demostró que esta afirmación era un producto del montaje del esqueleto inicial y tres dígitos es más probable, como se ve en los anquilosáuridos relacionados.[3] En términos generales, tanto los metatarsianos de la mano como los de los pies son muy robustos y fuertes. La cintura escapular está representada por un escapulocoracoides que mide unos 60 cm de largo, húmero, radio y ulna , estos elementos son muy robustos en comparación con otros poscraneales y pertenecen al brazo izquierdo. El escapulocoracoides izquierdo es muy robusto con una inserción bien desarrollada para el húmero, cavidad glenoide, aunque la hoja escapular es algo más corta en longitud. El húmero es muy ancho en los extremos y engrosado mide 33,5 cm (335 mm) de largo; muestra cierto parecido con el de Ankylosaurus . Aunque fragmentaria, la cintura pélvica está representada por un ilion parcial y aplanado con el isquion. Los elementos de las extremidades posteriores incluyen el fémur derecho y la tibia izquierda. El fémur preservado es recto y fuertemente robusto, el trocánter mayor está fusionado con el trocánter menor y mide 47 cm de largo. La tibia, sin embargo, es más corta pero muy ancha, tiene una longitud total de 29,8 cm. Además, también se conservó un peroné tiene una longitud de 26,7 cm. En general, el peroné es más recto y delgado en comparación con el fémur y la tibia, su extremo distal es más ancho que el extremo proximal.[1][8]
Descubrimiento e investigación

Se han descubierto restos de Talarurus en las partes sureste del desierto de Gobi en lo que ahora es Mongolia. El espécimen holotipo PIN 557-91 fue descubierto en 1948 por la Expedición Paleontológica Conjunta soviético-Mongola, y desenterrado de arcilla calcárea roja arenosa en la localidad de Baynshire de la Formación Bayan Shireh. Talarurus fue descrito y nombrado por el paleontólogo ruso Evgeny Maleev en 1952 . La muestra PIN 557, el holotipo original designado por Maleev, incluía un cráneo fragmentario con la parte posterior del techo del cráneo, incluida la región occipital y el basicráneo, numerosas vértebras, varias costillas, un escapulocoracoides, un húmero, un radio, un cúbito, una mano casi completa, un ilion parcial, un isquion, un fémur, una tibia, un peroné, un pie casi completo y una variedad de osteodermos y escudos. El nombre genérico , Talarurus , se deriva del griego τάλαρος, "tálaros", que significa cesto o jaula de mimbre, y οὐρά, "ourā́", que significa parte trasera o cola, en referencia al extremo del rabo de la cola que se asemeja a una canasta de mimbre, y la longitud de la cola que consiste en puntales óseos entrelazados, que recuerdan el tejido que se emplea para hacer cestas de mimbre. El nombre de la especie, T. plicatospineus, se deriva del latín plicātus, que significa doblado y spīneus, que significa espinoso, en referencia a los numerosos osteodermos que dieron forma a su coraza en vida.[1] De hecho, consistía en restos fragmentarios de seis individuos descubiertos en el sitio.[9] En 1977, Teresa Maryańska eligió PIN 557-91, una parte posterior posterior del cráneo, como holotipo, dadas las irregularidades de los individuos combinados. Además, cambió el nombre de Syrmosaurus disparoserratus a una segunda especie, Talarurus disparoserratus.[7] Sin embargo, en 1987, este fue rehecho en el género separado Maleevus.[9] Los elementos de todos estos especímenes se combinaron en una montura esquelética exhibida en el Museo Orlov. Aunque muy completa, en varios aspectos la montura es muy imprecisa.[2][3] El tipo y única especie válida conocida hoy en día es Talarurus plicatospineus.[3][6]

Talarurus ahora se conoce a partir de al menos una docena de especímenes individuales de varias localidades. El espécimen PIN 3780/1 fue recolectado de sedimentos terrestres en la localidad de Bayshin-Tsav de la Formación Bayan Shireh, por una expedición paleontológica conjunta soviética-mongola en 1975 y ahora está depositado en el Instituto Paleontológico de la Academia Rusa de Ciencias, en Moscú. Este material fue asignado a Talarurus y también se considera que data de la etapa Turoniense del Cretácico. Este espécimen consiste en la parte superior de un cráneo y un esqueleto fragmentario.[10] Desde 2006, en el contexto del Proyecto Internacional Conjunto de Dinosaurios Corea-Mongolia, numerosos especímenes adicionales han sido referidos a Talarurus, que se encuentra en las localidades de Baynshire y Shine Us Khuduk. Estos en 2014 aún no estaban descritos.[2] Otro espécimen referido a este género de la localidad de Bayshin Tsav está compuesto por un cráneo incompleto, no descrito, con techo craneal, parte occipital y caja craneana. Un segundo espécimen no descrito, recolectado en la localidad de Baga Tarjach, consiste en un fragmento de un maxilar con ocho dientes. Arbor ha enumerado muchos de los materiales adicionales y referidos para Talarurus.[2]
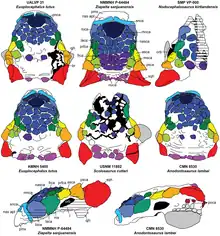
Incluso se ha encontrado más material adicional. En 2007, durante la Expedición Internacional de Dinosaurios Corea-Mongolia, se recolectaron especímenes MPC-D 100/1354, MPC-D 100/1355 y MPC-D 100/1356 de la parte oriental del desierto de Gobi, en la localidad de Bayn Shire en la Formación Bayan Shireh.[11] Estos especímenes se describieron en detalle en 2019 y consisten en tres cráneos medianos conservados con datos anatómicos y ecológicos adicionales para Talarurus. Este nuevo material ahora se encuentra alojado permanentemente en el Instituto de Paleontología y Geología de la Academia de Ciencias de Mongolia.[6]
Clasificación
Las relaciones filogenéticas de los anquilosáuridos son difíciles de determinar porque muchos taxones solo se conocen parcialmente, la configuración exacta de la armadura rara vez se ha conservado, los osteodermos fusionados oscurecen muchos detalles del cráneo y los Ankylosauridae son conservadores en su esqueleto poscraneal, mostrando poca variación en sus vértebras, pelvis y extremidades. Anteriormente se suponía que, como uno de los anquilosáuridos más antiguos conocidos, Talarurus poseía algunas características basales que se comparten con los nodosáuridos pero que luego se perdieron en los anquilosaurios más avanzados, como la presencia de cuatro dedos en los pies. Sin embargo, los supuestos rasgos "primitivos" demostraron ser en gran parte artefactos de la restauración esquelética inicial. El análisis filogenético reciente proporciona evidencia de una asignación de Talarurus a los Ankylosaurinae, un grupo derivado de anquilosáuridos. Esto puede conciliarse con su edad geológica relativamente antigua por la posibilidad de que los Ankylosauridae en su conjunto aparecieran mucho antes durante el Jurásico Temprano, lo que debe haber sido cierto si fueran el grupo hermano de los Nodosauridae en el sentido propuesto por Coombs en 1978, es decir, si todos los polacantinos fueran nodosáuridos.[12][3]
Talarurus fue asignado a Syrmosauridae, ahora conocido como Ankylosauridae, por Maleev en su descripción original en 1952.[1] Walter Preston Coombs sugirió que era el mismo dinosaurio que Euoplocephalus, aunque estudios posteriores no respaldaron esta afirmación.[12] Maryańska demostró que difería de Euoplocephalus, citando la forma del cráneo, la morfología del paladar y la presencia de cuatro dedos pedales.[7] Vickaryous et al. de 2004 señala la presencia de dos clados de anquilosáuridos distintos durante el Cretácico superior, uno compuesto por taxones norteamericanos y el otro restringido a taxones asiáticos..[13] Sin embargo, Arbor en 2014 recuperó árboles en los que Talarurus estaba más estrechamente relacionado con las formas norteamericanas que con los anquilosáuridos asiáticos. En algunos de estos , Talarurus era la especie hermana de Nodocephalosaurus.[2]
Filogenia

Arból filogenético que muestras las relaciones propuestas por Arbour en 2014.[2]
| Ankylosaurinae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiología
Los cráneos de especímenes de 2007 han aportado nuevos conocimientos sobre las capacidades neurocraneales y los hábitos dietéticos de Talarurus, específicamente el espécimen MPC-D 100/1354, que es un cráneo casi completo y bien conservado. MPC-D 100/1354 fue descrito con gran detalle junto con un cráneo muy completo de Tarchia por Paulina-Carabajal y colegas en 2017. Observaron que los anquilosáuridos tenían una estabilización de la mirada y sentidos auditivos bien desarrollados, a diferencia de los nodosáuridos, al examinar la región endocraneal de los especímenes seleccionados.[11] La presencia del flóculo fue reportada por primera vez en Talarurus por Maryańska[7] y más tarde en otros anquilosáuridos relacionados, sin embargo, este lóbulo parece estar ausente o reducido en los nodosáuridos. El flóculo es relativamente grande en la mayoría de los anquilosáuridos. Otro carácter neuroanatómico es la lagena alargada, que es prominente en Euoplocephalus, Tarchia y Talarurus . Esta característica anatómica indica que los anquilosáuridos tenían un amplio rango de percepción del sonido, especialmente para las bajas frecuencias.[11] Además de estos hallazgos, la boca conservada en MPC-D 100/1354 es ancha, de forma casi rectangular y algo robusta. La amplia boca de Talarurus probablemente trabajó con eficiencia en vegetación baja en terreno plano. La evidencia parece indicar que Talarurus tenía un método de alimentación de pastoreo.[6]
Paleoecología

Los restos fósiles de Talarurus se han encontrado en los sedimentos de la Formación Bayan Shireh, que se cree que datan de las etapas Cenomaniense a Santoniense del período Cretácico superior, hace unos 96 millones a 89 millones de años.[14] Según Park et al. en 2019, Talarurus llenó el nicho de un herbívoro de pastoreo, mientras que Tsagantegia , otro anquilosáurido de la formación, llenó el nicho de un herbívoro ramoneador. Por ejemplo, Talarurus tenía un hocico, o pico, ancho y rectangular, útil para la vegetación baja, mientras que el de Tsagantegia es más alargado y en forma de pala, útil para la vegetación alta. Para ilustrar esta estrategia de diferenciación, compararon a los anquilosáuridos con los rinocerontes africanos blancos y negros existentes. El rinoceronte blanco está equipado con un hocico ancho y rectangular, especializado para pastar. El rinoceronte negro, por otro lado, tiene un hocico puntiagudo para comer. Con estas características anatómicas y el ejemplo, está claro que estos taxones estaban divididos en nichos.[6] Otro caso de partición de nicho se puede observar en los terizinosaurios Erlikosaurus y Segnosaurus, taxones que también fueron descubiertos en la formación.[15]
Encontrados en localidades del Cretácico superior como Baynshire y Bayshin Tsav,[16] los taxones que eran contemporáneos con Talarurus en el Alto Bayan Shireh incluían una diversa fauna de dinosaurios, como el gran dromeosáurido Achillobator,[17] los terizinosáuridos Erlikosaurus[18] y Segnosaurus,[18] el ornitomimosaurio Garudimimus,[19] el pequeño y probablemente inmaduro ceratópsido Graciliceratops,[20] el pequeño hadrosauroideo Gobihadros,[16] y su homólogo ecológico, Tsagantegia.[21] Basado en la sedimentación del caliche, de tipo fluvial y lacustre, la Formación Bayan Shireh era un gran terreno semiárido con meandros y lagos,[22][23] junto con la fuerte presencia de plantas angiospermas y gimnospermas en toda la zona.[24][25] De acuerdo con algunas ocurrencias bioestratigráficas dentro de las formaciones asiáticas, hubo una probable correlación entre la Formación Iren Dabasu y la Formación Bayan Shireh.[26][27]
Referencias
- Maleev, E. A. (1952). «Noviy ankilosavr is verchnego mela Mongolii» [A new ankylosaur from the Upper Cretaceous of Mongolia]. Doklady Akademii Nauk SSSR (en russian) 87 (2): 273-276.
- Arbour, V. M. (2014). «Systematics, evolution, and biogeography of the ankylosaurid dinosaurs». Ph.D Thesis: 265.
- Arbour, V. M.; Currie, P. J. (2015). «Systematics, phylogeny and palaeobiogeography of the ankylosaurid dinosaurs». Journal of Systematic Palaeontology 14 (5): 385-444. S2CID 214625754. doi:10.1080/14772019.2015.1059985.
- Paul, G. S. (2010). The Princeton Field Guide to Dinosaurs. Princeton, New Jersey: Princeton University Press. pp. 231. ISBN 978-0-6911-3720-9.
- Holtz, T. R.; Rey, L. V. (2007). Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. Random House. Genus List for Holtz 2012 Weight Information
- Park, J. Y.; Lee, Y. N.; Currie, P. J.; Kobayashi, Y.; Koppelhus, E.; Barsbold, R.; Mateus, O.; Lee, S. et al. (2019). «Additional skulls of Talarurus plicatospineus (Dinosauria: Ankylosauridae) and implications for paleobiogeography and paleoecology of armored dinosaurs». Cretaceous Research 108: 104340. S2CID 212423361. doi:10.1016/j.cretres.2019.104340.
- Maryańska, T. (1977). «Ankylosauridae (Dinosauria) from Mongolia». Palaeontologia Polonica 37: 85-151. Archivado desde el original el 12 de julio de 2020. Consultado el 23 de agosto de 2022.
- Maleev, E. A. (1956). «Pantsyrnye dinosavry verchnego mela Mongolii (Semeustvo Ankylosauridae)» [Armored Dinosaurs of the Upper Cretaceous of Mongolia Family Ankylosauridae]. Trudy Paleontologicheskogo Instituta Akademiy Nauk SSSR (en russian) 62: 51-91.
- Tumanova, T. A. (1987). «The armored dinosaurs of Mongolia». The Joint Soviet-Mongolian Paleontological Expedition Transaction (en russian) 32: 1-76.
- Kurzanov, S. M.; Tumanova, T. A. (1978). «The structure of the endocranium in some Mongolian ankylosaurs». Paleontologicheskii Zhurnal (en russian) 3: 369-374.
- Paulina-Carabajal, A.; Lee, Y. N.; Kobayashi, Y.; Lee, H. J.; Currie, P. J. (2017). «Neuroanatomy of the ankylosaurid dinosaurs Tarchia teresae and Talarurus plicatospineus from the Upper Cretaceous of Mongolia, with comments on endocranial variability among ankylosaurs». Palaeogeography, Palaeoclimatology, Palaeoecology 494: 135-146. Bibcode:2018PPP...494..135P. doi:10.1016/j.palaeo.2017.11.030.
- Coombs, W. P. (1978). «The families of the ornithischian dinosaur order Ankylosauria». Palaeontology 21 (1): 143-170.
- Vickaryous, M. K.; Maryańska, T.; Weishampel, D. B. (2004). «Ankylosauria». The Dinosauria, Second Edition. University of California Press. pp. 363–392. ISBN 978-0-5202-5408-4.
- Kurumada, Y.; Aoki, S.; Aoki, K.; Kato, D.; Saneyoshi, M.; Tsogtbaatar, K.; Windley, B. F.; Ishigaki, S. (2020). «Calcite U–Pb age of the Cretaceous vertebrate‐bearing Bayn Shire Formation in the Eastern Gobi Desert of Mongolia: usefulness of caliche for age determination». Terra Nova 32 (4): 246-252. Bibcode:2020TeNov..32..246K. doi:10.1111/ter.12456. Parámetro desconocido
|doi-access=ignorado (ayuda) - Zanno, L. E.; Tsogtbaatar, K.; Chinzorig, T.; Gates, T. A. (2016). «Specializations of the mandibular anatomy and dentition of Segnosaurus galbinensis (Theropoda: Therizinosauria)». PeerJ 4: e1885. PMC 4824891. PMID 27069815. doi:10.7717/peerj.1885.
- Khishigjav Tsogtbaatar; David B. Weishampel; David C. Evans; Mahito Watabe (2019). «A new hadrosauroid (Dinosauria: Ornithopoda) from the Late Cretaceous Baynshire Formation of the Gobi Desert (Mongolia)». PLOS ONE 14 (4): e0208480. Bibcode:2019PLoSO..1408480T. PMC 6469754. PMID 30995236. doi:10.1371/journal.pone.0208480. Parámetro desconocido
|doi-access=ignorado (ayuda) - Perle, A.; Norell, M. A.; Clark, J. (1999). «A new maniraptoran Theropod−Achillobator giganticus (Dromaeosauridae)−from the Upper Cretaceous of Burkhant, Mongolia». Contributions from the Geology and Mineralogy Chair, National Museum of Mongolia (101): 1-105. OCLC 69865262.
- Barsbold, R.; Perle, A. (1980). «Segnosauria, a new suborder of carnivorous dinosaurs». Acta Palaeontologica Polonica 25 (2): 190-192.
- Kobayashi, Y.; Barsbold, R. (2005). «Reexamination of a primitive ornithomimosaur, Garudimimus brevipes Barsbold, 1981 (Dinosauria: Theropoda), from the Late Cretaceous of Mongolia». Canadian Journal of Earth Sciences 42 (9): 1501−1521. Bibcode:2005CaJES..42.1501K. doi:10.1139/e05-044. hdl:2115/14579.
- Sereno, P. C. (2000). «The fossil record, systematics and evolution of pachycephalosaurs and ceratopsians from Asia». The Age of Dinosaurs in Russia and Mongolia. Cambridge University Press. pp. 489-491.
- Tumanova, T. A. (1993). «O novom pantsirnov dinozavre iz iugo-vostochnoy Gobi» [A new armored dinosaur from Southeastern Gobi]. Paleontologicheskii Zhurnal (en russian) 27 (2): 92-98.
- Jerzykiewicz, T.; Russell, D. A. (1991). «Late Mesozoic stratigraphy and vertebrates of the Gobi Basin». Cretaceous Research 12 (4): 345-377. ISSN 0195-6671. doi:10.1016/0195-6671(91)90015-5.
- Hicks, J. F.; Brinkman, D. L.; Nichols, D. J.; Watabe, M. (1999). «Paleomagnetic and palynologic analyses of Albian to Santonian strata at Bayn Shireh, Burkhant, and Khuren Dukh, eastern Gobi Desert, Mongolia». Cretaceous Research 20 (6): 829-850. doi:10.1006/cres.1999.0188.
- Khand, Y.; Badamgarav, D.; Ariunchimeg, Y.; Barsbold, R. (2000). «Cretaceous system in Mongolia and its depositional environments». Cretaceous Environments of Asia. Developments in Palaeontology and Stratigraphy 17. pp. 49-79. ISBN 9780444502766. doi:10.1016/s0920-5446(00)80024-2.
- Ksepka, D. T.; Norell, M. A. (2006). «Erketu ellisoni, a long-necked sauropod from Bor Guvé (Dornogov Aimag, Mongolia)». American Museum Novitates (3508): 1-16. doi:10.1206/0003-0082(2006)3508[1:EEALSF]2.0.CO;2.
- Averianov, A.; Sues, H. (2012). «Correlation of Late Cretaceous continental vertebrate assemblages in Middle and Central Asia». Journal of Stratigraphy 36 (2): 462-485. S2CID 54210424. Archivado desde el original el 7 de marzo de 2019. Parámetro desconocido
|url-status=ignorado (ayuda) - Tsuihiji, T.; Watabe, M.; Barsbold, R.; Tsogtbaatar, K. (2015). «A gigantic caenagnathid oviraptorosaurian (Dinosauria: Theropoda) from the Upper Cretaceous of the Gobi Desert, Mongolia». Cretaceous Research 56: 60-65. doi:10.1016/j.cretres.2015.03.007.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikispecies tiene un artículo sobre Talarurus plicatospineus.
Wikispecies tiene un artículo sobre Talarurus plicatospineus. Wikimedia Commons alberga una categoría multimedia sobre Talarurus plicatospineus.
Wikimedia Commons alberga una categoría multimedia sobre Talarurus plicatospineus.