Segnosaurus galbinensis
Segnosaurus galbinensis es la única especie conocida del género extinto Segnosaurus (lat. «reptil lento») de dinosaurio terópodo tericinosáurido, que vivió a mediados del período Cretácico, hace aproximadamente entre 98 y 90 millones de años, en el Cenomaniense y el Turoniense, en lo que hoy es Asia. En la década de 1970 se descubrieron múltiples especímenes incompletos pero bien conservados en el desierto de Gobi de Mongolia y en 1979 se nombró al género y la especie Segnosaurus galbinensis.[1] El nombre genérico Segnosaurus significa "lagarto lento" y el nombre específico S. galbinensis se refiere a la región de Galbin. El material conocido de este dinosaurio incluye la mandíbula inferior, las vértebras del cuello y la cola, la pelvis, la cintura escapular y los huesos de las extremidades. Partes de las muestras han desaparecido o se han dañado desde que fueron recolectadas.
| Segnosaurus galbinensis | ||
|---|---|---|
| Rango temporal: 98 Ma - 90 Ma Cretácico superior | ||
 Recreación de un Segnosaurus. | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| Infraorden: | Therizinosauria | |
| Familia: | Therizinosauridae | |
| Género: |
Segnosaurus Perle, 1979 | |
| Especie: |
S. galbinensis Perle, 1979 | |
Segnosaurus era un terizinosaurio de gran tamaño que se estima que medía entre 6 y 7 metros de largo y pesaba alrededor de 1,3 toneladas. Habría sido bípedo, con el tronco de su cuerpo inclinado hacia arriba. La cabeza era pequeña con un pico en la punta de las mandíbulas y el cuello era largo y delgado. La mandíbula inferior estaba inclinada hacia abajo en la parte delantera y los dientes se distinguían por tener dentículos adicionales, así como terceros bordes cortantes en algunos de los dientes posteriores. Las extremidades anteriores eran robustas y tenían tres dedos que tenían grandes garras, y los pies tenían cuatro dedos que sostenían el pie, aparte de los terizinosaurios, todos los terópodos tenían pies de tres dedos. La parte delantera de la pelvis se adaptó para soportar el abdomen agrandado. El pubis se volvió hacia atrás, una característica que solo se ve en las aves y los dinosaurios más estrechamente relacionados con ellas.
Las afinidades de Segnosaurus eran originalmente oscuras y recibió su propia familia de terópodos, Segnosauridae, y más tarde, cuando se identificaron géneros relacionados, un infraorden, Segnosauria. Se propusieron esquemas de clasificación alternativos hasta que se describieron parientes más completos en la década de 1990, lo que los confirmó como terópodos. Los nuevos fósiles también mostraron que Segnosauridae era un sinónimo menor de la anterior familia Therizinosauridae. Se cree que el Segnosaurus y sus parientes eran animales de movimiento lento que, como lo indican sus características inusuales, eran principalmente herbívoros, mientras que la mayoría de los otros grupos de terópodos eran carnívoros. Los terizinosaurios probablemente usaban sus largas patas delanteras, cuellos largos y picos cuando ramoneaban y tripas grandes para procesar alimentos. Segnosaurus es conocido de la Formación Bayan Shireh, donde vivió junto a sus compañeros terizinosaurios Erlikosaurus y Enigmosaurus, estos géneros relacionados probablemente estaban divididos por nichos.
Descripción

Segnosaurus era un terizinosaurio de gran tamaño que se estima que medía entre 6 y 7 metros de largo y pesaba alrededor de 1,3 toneladas.[2][3][4] Campione & Evans en 2020, sin embargo, calcularon su peso en 4,2 a 4,6 toneladas, con rangos de 3,1 a 5,8 toneladas.[5] Segnosaurus no se conoce por completo, pero como terizinosaurio , habría sido bípedo y de construcción robusta con el tronco del cuerpo inclinado hacia arriba en comparación con otros terópodos. La cabeza habría sido pequeña con un rhamphotheca, o pico córneo, en la punta de las mandíbulas y un cuello largo y delgado. Los dedos no eran particularmente largos, pero tenían grandes garras. La parte delantera de la pelvis se adaptó para soportar el aparato digestivo agrandado.[2][4][1] Se sabe que los terizinosaurios tenían plumas simples y primitivas , como lo demuestran los fósiles del género basal, o "primitivo", Beipiaosaurus, el segundo dinosaurio conocido que no es ave conservado con tales tegumentos después de Sinosauropteryx y Jianchangosaurus.[6][7] Dado que la mayoría de los terizinosaurios no se conocen por completo, no se sabe cuántas de las características anatómicas se utilizan para distinguir a los Segnosaurus están muy extendidas dentro del grupo, muchos géneros no pueden compararse directamente porque no se conservan los huesos equivalentes.[8][4]
Mandíbulas

Las dos mandíbulas de Segnosaurus son conocidas, faltando solamente pequeñas porciones de los huesos anatómicos y el hueso angular entero. La dentadura es muy compleja comparada con otros tericinosáuridos. La mandíbula de Segnosaurus era baja y alargada, pero relativamente robusta y sin forma en comparación con la de Erlikosaurus, que era más grácil. La hemimandibula derecha, la mitad de la mandíbula, casi completa mide 379 milímetros de largo de adelante hacia atrás, 55,5 milímetros en el punto más alto y 24,5 milímetros en el más bajo. El hueso dentario, el hueso portador de dientes que forma la mayor parte de la parte frontal de la mandíbula, tenía una forma compleja en comparación con los primeros terizinosaurios. La parte portadora de dientes era casi rectangular y se inclinaba hacia abajo en la vista lateral con un arco pronunciado a lo largo de la longitud superior del extremo frontal, más extremo que lo que se conoce en otros terizinosaurios. La parte más frontal del dentario se desvió fuertemente hacia abajo en un ángulo de aproximadamente 30 grados, una característica única de este género. Cuando cada hemimandible se articula con el otro, forman una sínfisis mandibular sin dientes, en forma de U, que se proyecta hacia arriba, hacia el frente, como en Erlikosaurus y Neimongosaurus. La región frontal expansiva y sin dientes del dentario se extiende 25,5 milímetros En la hemimandibula derecha del holotipo. Proporcionalmente, la parte sin dientes del dentario es el 20% de su fila de dientes, que tiene 150,3 milímetros de largo. En comparación, la región desdentada de Erlikosaurus tenía aproximadamente el 12% de la longitud de la fila de dientes y estaba casi ausente en Jianchangosaurus. La altura del dentario disminuyó hacia la última extensión de la hilera de dientes, después de lo cual se abanicó bruscamente para hacer contacto con el hueso surangular detrás de él; por el contrario, la parte trasera del dentario en Erlikosaurus se acercó gradualmente al surangular en un arco suave.[4][1]

Segnosaurus fue distinto entre los terizinosaurios en que la parte posterior del dentario no tenía dientes. Los dientes estaban restringidos a los dos tercios frontales del dentario, que tenía 24 alvéolos, cavidades dentales, de una manera similar a Jianchangosaurus pero diferente de Erlikosaurus, en el que casi todo el dentario estaba dentado, con 31 alvéolos. La fila de dientes de Segnosaurus estaba insertada y demarcada por un estante en el lado exterior como lo estaba en todos los terizinosaurios derivados o "avanzados". A diferencia de otros taxones relacionados, la plataforma estaba restringida a la parte posterior del dentario y el borde elevado que lo definía no era tan pronunciado. Segnosaurus era único en tener una cresta baja que se elevaba entre el quinto y el decimocuarto alvéolos que dividía el dentario en dos partes delantera y trasera de casi el mismo tamaño. Justo encima de esta cresta, el dentario estaba atravesado por una hilera de agujeros como en Jianchangosaurus y Alxasaurus, que se volvieron menos regulares en la región alrededor de la sínfisis mandibular, donde las dos mitades de la mandíbula se unían en la parte delantera. En cambio, esta fila estaba directamente en línea con y en el lado de la cresta en Erlikosaurus. El surco meckeliano que corría a lo largo del lado interno de la mandíbula, se colocó más abajo que en Erlikosaurus.y tuvo una profundidad constante hasta la posición del decimotercer diente, después de lo cual se ensanchó. Los elementos de la mandíbula inferior detrás del dentario, los huesos esplenial , surangular, angular y prearticular, eran distintos de los de otros terizinosaurios, eran gráciles y lineales, y contribuían a que la parte trasera del hemimandibula fuera alargada y casi rectangular.[4] El surangular era largo y en forma de espada, el angular tenía forma de ala, el prearticular era estrecho y curvo, y el esplenial era delgado y de contorno triangular. La fenestra mandibular externa, una abertura en el lado externo de la mandíbula, era más grande que la de Erlikosaurus porque el surangular era poco profundo de arriba abajo.[1]

Segnosaurus tenía la menor cantidad de dientes en el dentario, 24 en cada mitad determinado a partir del número de alvéolos, así como los dientes más grandes conocidos entre los terizinosaurios. Los dientes dentarios eran foliodontes, en forma de hoja y llevaban coronas comprimidas lateralmente agrandadas, relativamente altas, con una ligera recursividad en el margen superior de las puntas. En comparación, los dientes de Erlikosaurus eran más pequeños, simétricos y más simples. Las bases de las coronas aumentaron ligeramente de tamaño hacia atrás a lo largo de la fila de dientes, lo que reflejó una disminución en la compresión lateral. Las superficies frontales de las coronas y los lados que miraban hacia afuera eran convexas, mientras que los lados que miraban hacia adentro eran cóncavos. Una cresta engrosada corría a lo largo de la longitud del lado orientado hacia adentro cerca de la mitad superior de la corona, que estaba flanqueada por surcos débiles cerca de los bordes frontal y posterior de los dientes, llegando casi hasta el cuello del diente, la transición entre la corona y raíz, de los dientes. En general, los 18 dientes más frontales eran relativamente homodónticos, del mismo tipo, aunque la corona del segundo diente era relativamente más corta y cónica. Esto también puede haber sido cierto para el primer diente, pero no se conservó. Los dientes más atrás en la fila también disminuyeron en altura relativa hacia atrás. En comparación, los cuatro o cinco dientes dentarios delanteros deLos Erlikosaurus eran conidontes, en forma de cono, con una transición gradual a dientes foliodontes.[1][4]
Los dientes dentarios estaban apretados, pero no comprimidos juntos, con las coronas de los dientes acercándose entre sí en la mitad de su longitud. Los dentículos, estrías, eran grandes y bulbosos, disminuyendo ligeramente de tamaño hacia las puntas de los dientes, con aproximadamente 5-6 dentículos por 3 milímetros. La carina delantera, bordes cortantes, doblados hacia arriba para superponer la superficie interna de las coronas en los dientes tercero a décimo octavo, pero tales pliegues estaban ausentes en la segunda y probablemente la primera corona. Los dentículos eran aproximadamente perpendiculares con la punta de las coronas de los dientes, pero paralelos a la altura de la corona en el pliegue del lado frontal y la faceta triangular en el lado posterior. Había una serie de dentículos accesorios (además de los de la carina) que se proyectaban desde las superficies frontales de los pliegues carinales, lo que hacía que los bordes frontales de las coronas fueran más rugosos. Las carinas de los bordes posteriores también estaban muy modificadas y bifurcadas, partidas en dos, cerca del cuello del diente, donde formaban una faceta triangular aplanada y elevada, que se proyectaba desde la corona del diente y contactaba o se acercaba a las carinas plegadas en el borde frontal de las coronas detrás de ellas,esta disposición está presente en los dientes 2-12. Dichas carinas divididas se conocen de otrosterópodos tetanuranos, donde se consideran anomalías causadas por traumatismos, reemplazo de dientes aberrantes o factores genéticos. Aunque la condición en Segnosaurus era similar, se expresaba uniformemente en los dientes de ambos dentarios y no parece haber sido una anomalía, pero sirvió para endurecer los contactos entre las bases de los dientes.[4]
Los dientes dentarios 22 y 23 de Segnosaurus eran significativamente más pequeños que el resto, casi conidontes y tenían una tercera carina adicional con dentículos en sus lados internos. La mayoría de las otras coronas de los últimos dientes están dañadas, por lo que se desconocen sus características completas. La carina adicional en el diente 23 parece haber sido completamente denticulada mientras que los dentículos estaban restringidos al lado basal de la corona en el diente 22. Segnosaurus fue único entre todos los terópodos conocidos en poseer triple carina. El decimocuarto alvéolo en el dentario derecho del holotipo está amurallado por aparentemente debido a una lesión o enfermedad, que genera crecimiento óseo, pero los dientes en esa parte del dentario están dañados, por lo que no es posible determinar cómo los dientes se vieron afectados por esto. Los dientes de la misma zona del dentario izquierdo presentan triple carina, aunque este dentario no tiene indicios externos de patología que pudieran haber llevado a esta condición, por lo que no se puede concluir ni descartar que este rasgo sea el resultado de una patología. Segnosaurus reemplazó sus dientes en ondas que iban desde atrás hacia adelante de las mandíbulas, que abarcaban de dos a tres coronas en erupción. Algunos de los dientes completamente erupcionados tienen desgaste en las carinas de sus lados traseros, a diferencia de lo que se ve en otros terizinosaurios. La textura del esmalte parece haber sido ampliamente irregular y las raíces de los dientes eran casi circulares.[4]
Esquelto postcraneal
La escápula de Segnosaurus era recta y plana en el extremo superior, y estaba fusionada al hueso coracoides, formando el escapulocoracoides. El coracoides era muy ancho, de contorno rectangular y grueso en el medio. El húmero, masivo, tenía 560 milímetros de longitud, tenía un eje casi cilíndrico y cóndilos bien definidos para la articulación con el radio y el cúbito del antebrazo. La cresta deltopectoral, donde el músculo deltoides estaba unido a la parte frontal superior del húmero, estaba bien desarrollada.[1] El húmero era distinto a los de otros terizinosaurios, ya que era recto en lugar de forma sigmoidea y no expandido o desviado hacia adelante en su extremo superior. El húmero tampoco se expandió en el medio y el entepicóndilo no estaba bien desarrollado. La falta de estas características era más similar a los ornitomimosaurios y trodóntidos que a otros terizinosaurios.[8] El radio también era masivo, alrededor del 60 por ciento del húmero, con un eje recto. El cúbito era más grueso que el radio y un poco más largo, alrededor del 70 por ciento del húmero y estaba ligeramente torcido a lo largo de su eje medio. La mano tenía tres dedos. Los huesos de la falange de los dedos estaban aplanados de arriba abajo y las depresiones articulares de los lados no estaban muy desarrolladas. La primera falange del primer dedo era larga y delgada, mientras que la primera y la segunda falanges del segundo dedo eran cortas. El ungual del tercer dedo era algo más largo que la segunda falange y bastante plano de arriba abajo, lo que puede haber sido una característica única de Segnosaurus. Este ungual era muy curvado, muy puntiagudo y comprimido de lado a lado. El tubérculo inferior, donde los tendones flexores se unían al ungual, era grueso y robusto.[1][8][9][10]

La pelvis de Segnosaurus era robusta y tenía lóbulos marcadamente dirigidos hacia los lados en la parte delantera. La pelvis se acortó en la parte delantera, una característica que se encuentra entre los terópodos parecidos a las aves, pero poco común entre los terópodos en general.[1] El hueso púbico se dirigió hacia atrás y hacia abajo en paralelo con el isquion, esta orientación hacia atrás del hueso púbico se conoce como condición opistopúbica. Esta característica sólo se conoce de las aves y sus más cercanos familiares celurosaurios, mientras que otros dinosaurios terópodos habían dirigido hacia delante huesos púbicos.[11][10] El hueso púbico estaba alargado, aplanado hacia los lados y tenía una proyección elipsoide o "bota" en la parte delantera de su extremo inferior.[9][10] La pelvis era distinta de las de otros terizinosaurios en que el margen superior del ilion tenía un saliente pronunciado en el lado inferior y que el proceso de proyección posterior del isquion era extenso, casi el 50 por ciento de la longitud de adelante hacia atrás del proceso obturador. Algunas características de la pelvis eran similares a las de Nothronychus , particularmente el isquion, pero no está claro si estas similitudes se debieron a que tenían un ancestro común con exclusión de otros terizinosaurios derivados, o porque conservaron características basales desde que se perdieron en otros parientes. El isquion de Segnosaurus era distinto al de Nothronychus en que tenía un proceso obturador casi rectangular y un foramen obturador casi circular. La pelvis se diferenciaba de la de Enigmosaurus por su proceso obturador profundo que no se fusionaba con su contraparte en el medio, por su bota púbica no fusionada y porque la parte inferior del eje púbico era ancha de adelante hacia atrás. Segnosaurus se diferenciaba de Nothronychus y Enigmosaurus por tener una fosa brevis profunda, un surco donde se originaba el músculo caudofemoralis brevis de la cola, y porque su bota púbica carecía de una proyección hacia atrás bien desarrollada.[8]
El fémur era recto con una sección transversal ovalada y tenía una longitud de 840 milímetros. La cabeza del fémur se colocó sobre un "cuello" largo y los cóndilos inferiores estaban bien definidos. La tibia era recta, ligeramente más corta que el fémur y estaba torcida a lo largo de su eje. El peroné era largo y estrecho hacia su extremo inferior. El metatarso del pie era corto, macizo y constaba de cinco huesos, cuatro de los cuales funcionaban como elementos de soporte y terminaban en cuatro dedos. Los pies funcionalmente tetradáctilos, de cuatro dedos, eran exclusivos de los terizinosaurios derivados, los terizinosaurios basales y todos los demás terópodos tenían pies tridáctilos en los que el primer dedo era corto y no llegaba al suelo. Externamente, el metatarso era similar, aunque proporcionalmente más grande, que los de los prosaurópodos, un grado evolutivo tempranode sauropodomorfos . Las epífisis de los metatarsianos superiores estaban hipertrofiadas, agrandadas, una característica distintiva del género. El primer dedo era más corto que los demás pero tenía la misma importancia funcional; el segundo y tercer dedo del pie eran igualmente largos, mientras que el cuarto era el más delgado. El dedo del pie ungual era robusto, muy curvado, plano a los lados y más puntiagudo que los de los prosaurópodos. El tubérculo inferior donde se unían los ligamentos flexores era robusto. Si bien la falta de una fuerte compresión de los unguales del dedo del pie distinguía a Segnosaurus de Erlikosaurus de la misma formación, la falta de compresión era común entre los terizinosaurios y, por lo tanto, no era exclusiva de Segnosaurus.[1][8][9] Las vértebras cervicales eran platicoelosas y tenían centros grandes y masivos y arcos neurales bajos. El sacro constaba de seis vértebras firmemente fusionadas. Los centros de estas vértebras estaban ensanchados y relativamente alargados, y cada centro era un poco más largo que su ancho. Las espinas neurales aquí no eran muy largas, pero sobrepasaban el nivel del ilíaco. Las vértebras caudales más cercanas al cuerpo eran enormes, altas y algo comprimidas de lado a lado. El arco neural era bajo con un pequeño canal neural. Las vértebras caudales más cercanas a la punta de la cola eran platicolas y tenían centros cortos y masivos. Los procesos transversales de las vértebras caudales y las costillas eran robustos y alargados.[1][9]
Descubrimiento e investigación

En 1973, una expedición soviético-mongola en la localidad de Amtgay, al norte de Khan-Bogdsomona descubrieron un fósil de dinosaurio. Durante 1974 y 1975 se descubrieron más restos del mismo taxón en Amtgay y Khara-Khutul. Los huesos estaban relativamente en buen estado y algunas regiones del esqueleto estaban al descubierto. Estos fósiles fueron descritos, en 1979, por, quien los describió con el nombre binomial de Segnosaurus galbiensis. El nombre genérico es derivado del Latín segnis, que significa "lento" o "rechoncho", y del Griego Antiguo, sauros, que significa "lagarto", por la masiva complexión del animal. El nombre específico hace referencia a la región Galbin del desierto del Gobi, donde los fósiles fueron encontrados. En la descripción de Segnosaurus durante 1979, Perle señaló que el conjunto único de rasgos presentes probablemente era representativo de una nueva familia, a la que llamó 'Segnosauridae' , y también señaló que la familia sólo podría ser provisionalmente clasificada al suborden Theropoda.[12] El espécimen de holotipo de la localidad de Amtgay se encuentra en la Academia de Ciencias de Mongolia con el número de espécimen IGM 100/80, anteriormente GIN 100/80. Incluye la mandíbula inferior, un húmero incompleto, un radio y cúbito completos , falanges de los dedos, un miembro anterior ungual, una pelvis casi completa, un fémur derecho incompleto, seis partes sacras vértebras, diez vértebras caudales desde la parte delantera de la cola, quince desde la parte posterior de la cola, la primera costilla gastral y fragmentos de las costillas dorsales. Dos especímenes más fueron designados como especímenes paratipo. El espécimen IGM 100/82 de la localidad de Khara Khutul incluye un fémur, tibia y peroné , tarsos y metatarsianos , cinco falanges de dedos incluyendo un pie ungual, fragmentos de costillas, ilia completa, la porción superior de un isquion y la parte inferior porción de un pubis. La muestra IGM 100/83 incluye un escapulocoracoides izquierdo, un radio, un cúbito, unguales de las extremidades anteriores y un fragmento de una vértebra cervical.[12][8] En 1980, otro espécimen de la localidad de Amtgay fue referido a Segnosaurus por Rinchen Barsbold y Perle, IGM 100/81, que incluía una tibia derecha y un peroné.[9]

En 1983, Barsbold enumeró muestras adicionales GIN 100/87 y GIN 100/88. En 2010, sin embargo, la paleontóloga Lindsay E. Zanno sugirió que estos pueden referirse a los paratipos IGM 100/82 e IGM 100/83, que ya se habían incluido en 1979, porque la traducción del ruso al inglés del artículo de Barsbold tiene varios errores tipográficos en lo que respecta al número de muestras. Zanno también señaló que en el momento de su estudio, había numerosos problemas con los especímenes de Segnosaurus IGM, incluyendo daños causados desde la recolección, desaparición de elementos del holotipo, identificación incorrecta de elementos asignados y más de un individuo con el mismo número de espécimen. Los elementos del holotipo a los que Zanno pudo acceder en 2010 incluían un hueso ilíaco severamente dañado, un sacro al que le faltaban las costillas sacras izquierdas con daño, por lo que no podía unirse bien con el resto del hueso ilíaco, y un hueso púbico e isquion sin sus porciones superiores. Se localizaron más huesos con el número de muestra IGM 100/82, pero no se mencionaron en la descripción de Perle, mientras que se desconocía el paradero de algunos elementos paratipo.[13][8] En una nueva descripción de 2016 de la mandíbula holotípica, que se había estudiado poco desde 1979, Zanno y sus colegas informaron que la mayoría de las coronas dentales se habían dañado después de la recolección y que a la mayoría les faltaban las puntas. De los dos hemimandibulas, las mitades de la mandíbula inferior, el derecho está casi completo, sólo faltaba la parte posterior y el frente superior de su sínfisis mandibular, el área donde se juntan las mitades de la mandíbula. El hemimandibula izquierdo está fragmentado y conserva la parte anterior con algún desplazamiento de hueso por aplastamiento.[4]
Clasificación

Perle asignó, en 1979, Segnosaurus a Segnosauridae. Al año siguiente, en una descripción sobre el nuevo taxón Erlikosaurus, Barsbold & Perle nombraron un nuevo orden para la familia, Segnosauria. En aquel momento, los dos géneros del grupo eran Segnosaurus y Erlikosaurus.[9] En ese punto también quedó claro que Therizinosaurus era un pariente cercano y el grupo por lo tanto, pasó a llamarse Therizinosauridae, un nombre que tiene prioridad; y que este grupo estaba formado por comedores de plantas en vez de por piscívoros como originalmente había presumido Perle. Segnosaurus y sus parientes, que ahora se reconocen como terizinosaurios, "reptiles guadaña", fueron considerados durante mucho tiempo un grupo enigmático. Su mosaico de características que se asemejan a las de diferentes grupos de dinosaurios y la escasez de sus fósiles llevaron a una controversia sobre sus relaciones evolutivas durante décadas después de su descubrimiento inicial, los elementos de las extremidades anteriores del propio Therizinosaurus se identificaron originalmente como pertenecientes a una tortuga gigante cuando se describieron en 1954.[14][15][10] En 1979, Perle notó que los fósiles de Segnosaurus eran posiblemente representativos de una nueva familia de dinosaurios, a la que llamó Segnosauridae, siendo Segnosaurus el género tipo y miembro único. Tentativamente clasificó a los Segnosauridae como terópodos, tradicionalmente considerados como los dinosaurios "carnívoros", señalando similitudes en la mandíbula y sus dientes frontales. Utilizando características de sus húmero y garras de mano, distinguió Segnosauridae de las familias de terópodos Deinocheiridae y Therizinosauridae, que entonces solo se conocían de los géneros Deinocheirus y Therizinosaurus, respectivamente, representados principalmente por grandes extremidades anteriores que se encuentran en Mongolia.[1] Más tarde, en 1979, Barsbold y Perle encontraron que las características pélvicas de los segnosauridos y dromeosáuridos eran tan diferentes de las de los terópodos "verdaderos" que deberían separarse en tres taxones del mismo rango, posiblemente al nivel del infraorden dentro de Saurischia , una de las dos divisiones principales de los dinosaurios, siendo la otra Ornithischia.[16]
En 1980, Barsbold y Perle nombraron al nuevo infraorden de terópodos Segnosauria, que solo contiene Segnosauridae. En el mismo artículo, nombraron al nuevo género Erlikosaurus, conocido por un cráneo bien conservado y un esqueleto parcial, que tentativamente consideraron un segnosaurido, y reportaron una pelvis parcial de un segnosaurio indeterminado, ambos de la misma formación que Segnosaurus. Los especímenes proporcionaron datos relativamente completos sobre este grupo, estaban unidos por su pelvis opistopúbica, mandíbula delgada y la parte frontal desdentada de sus mandíbulas. Barsbold y Perle afirmaron que, aunque algunas de sus características se parecían a las de los ornitisquios y saurópodos, estas similitudes eran superficiales y distintas cuando se examinaban en detalle. Si bien eran esencialmente diferentes de otros terópodos, tal vez debido a que se separaron de ellos relativamente temprano, y justificaron un nuevo infraorden, mostraron similitudes con los terópodos. Debido a que el espécimen de Erlikosaurus carecía de pelvis, los autores no estaban seguros de que el segnosaurio indeterminado pudiera pertenecer al mismo género, en cuyo caso lo considerarían parte de una familia separada.[9] Aunque Erlikosaurus era difícil compararlo directamente con Segnosaurus porque sus restos estaban incompletos, Perle declaró en 1981 que no había justificación para separarlo en otra familia.[17]
En 1982, Perle informó del descubrimiento de fragmentos de extremidades posteriores similares a los de Segnosaurus y los asignó a Therizinosaurus, cuyas extremidades anteriores se habían encontrado en casi el mismo lugar. Concluyó que Therizinosauridae, Deinocheiridae y Segnosauridae, todos con extremidades anteriores agrandadas, representaban el mismo grupo taxonómico. Segnosaurus y Therizinosaurus eran particularmente similares, lo que llevó a Perle a sugerir que pertenecían a una familia con exclusión de Deinocheiridae, hoy, Deinocheirus es reconocido como un ornitomimosaurio.[18][19] Barsbold retuvo a Segnosaurus y Erlikosaurus en la familia Segnosauridae en 1983 y nombró al nuevo género Enigmosaurus basado en la pelvis segnosauriana previamente indeterminada. La estructura de la pelvis de Erlikosaurus era desconocida, pero Barsbold consideró poco probable que la pelvis de Enigmosaurus le perteneciera porque Erlikosaurus y Segnosaurus eran muy similares en otros aspectos, mientras que la pelvis de Enigmosaurus era muy diferente a la de Segnosaurus . Barsbold descubrió que los segnosauridos eran tan peculiares en comparación con los terópodos más típicos que eran una desviación muy significativa en la evolución de los terópodos o posiblemente estaban fuera del grupo. Sin embargo, los retuvo dentro de Theropoda.[13] Más tarde, en 1983, Barsbold declaró que la pelvis segnosauriana se desvió significativamente de la norma de los terópodos y encontró que la configuración de sus huesos ilíacos generalmente era similar a la de los saurópodos.[20]
Gregory S. Paul concluyó en 1984 que los segnosaurianos no tenían características terópodas sino que eran prosaurópodos del Cretácico que sobrevivieron tardíamente y que tenían adaptaciones similares a las de los ornitisquios. Encontró que los segnosaurianos eran similares a los prosaurópodos en la morfología de su hocico, mandíbula y retropié. A los ornitisquios en las mejillas, el paladar, el pubis y el tobillo y a los primeros dinosaurios en otros aspectos. Propuso que los ornitisquios descendían de los prosaurópodos y que los segnosaurianos eran una reliquia intermedia de esta transición, que supuestamente tuvo lugar durante el período Triásico. De esta manera, consideró que los segnosaurianos tenían una posición comparable a los dinosaurios herbívoros en general, que los monotremas tienen a los mamíferos. Lo encontró improbable, pero no descartó que los segnosaurianos pudieran haber derivado de terópodos o que los segnosaurianos, prosaurópodos y ornitisquios se derivaran independientemente de los primeros dinosaurios.[21] David B. Norman consideró la idea de Paul polémica y "destinada a provocar mucha discusión" en 1985.[22] En 1988, Paul sostuvo que los segnosaurianos eran prosaurópodos de tipo ornitisquio supervivientes tardíos y propuso una identidad segnosauriana para Therizinosaurus. También colocó segnosauria dentro de Phytodinosauria, un superorden que Robert Bakker había creado en 1985 para contener a todos los dinosaurios herbívoros.[23] En un estudio de 1986 sobre las interrelaciones de los dinosaurios saurisquios, Jacques Gauthier concluyó que los segnosaurianos eran prosaurópodos. Si bien admitió que tenían similitudes con los ornitisquios y los terópodos, propuso que estas características habían evolucionado de forma independiente.[24] En un resumen de la conferencia de 1989 sobre las interrelaciones de los sauropodomorfos, Paul Sereno también consideró a los segnosaurianos como prosaurópodos, basándose en las características del cráneo.[25]

En un artículo de revisión de 1990, Barsbold y Teresa Maryańska encontraron que Segnosauria era un grupo raro y aberrante de saurisquios en una posición no resuelta entre los sauropodomorfos y terópodos, y probablemente más cerca del primero. En consecuencia, los enumeraron como Saurischia sedis mutabilis, posición sujeta a cambios. Estuvieron de acuerdo en que las patas traseras asignadas a Therizinosaurus en 1982 eran segnosaurianas, pero no consideraron que esto fuera una justificación suficiente para que el propio Therizinosaurus fuera un segnosaurio porque solo se conocía de las extremidades anteriores.[26] En 1993, Dale A. Russell y Dong Zhi-Ming describieron el nuevo género Alxasaurus de China, en ese momento este era el terópodo grande más completo de su tiempo y lugar. Si bien Alxasaurus era similar en algunos aspectos a los prosaurópodos, la morfología detallada de sus extremidades lo vinculaba con Therizinosaurus y segnosaurianos. Debido a que se conservaron sus patas delanteras y traseras, Alxasaurus mostró que la asignación de Perle de extremidades traseras segnosaurianas a Therizinosaurus probablemente era correcta. Russell y Dong, por lo tanto, propusieron que Segnosauridae era un sinónimo menor del nombre más antiguo Therizinosauridae, y que Alxasaurus era el representante más conocido hasta ahora. También nombraron al nuevo rango taxonómico superior Therizinosauroidea para contener Alxasaurus y Therizinosauridae porque el nuevo género era algo diferente de sus parientes. Llegaron a la conclusión de que los terizinosaurios eran terópodos tetanuranos, más estrechamente relacionados con los ornitomimidos, troodóntidos y oviraptóridos, que colocaron juntos en el grupo Oviraptorosauria, porque encontraron Maniraptora , el grupo convencional de estos, inválido, y la taxonomía de nivel superior de los terópodos era inestable.[27][28][29]

Perle y sus colaboradores de un redescripción de 1994 del cráneo de Erlikosaurus, aceptaron la sinonimia de Segnosauridae con Therizinosauridae y consideraron therizinosauria como maniraptoriano, el grupo que también incluye las aves modernas, porque sí encontraron Maniraptora válida a través de su análisis. También discutieron las hipótesis anteriores alternativas para las afinidades de los terizinosaurios y demostraron fallas con ellos.[30] En 1995, Lev A. Nessov rechazó la idea de que los terizinosaurios fueran terópodos, los consideraba un grupo distinto dentro de Saurischia.[31] En 1996, Thomas R. Holtz Jr. encontró que los terizinosaurios se agrupaban con los oviraptorosaurios en un análisis filogenético de terópodos celurosaurios.[32] Russell acuñó el nombre Therizinosauria para el grupo más amplio en 1997.[28] En 1999, Xing Xu y sus colegas describieron Beipiaosaurus, un pequeño terizinosaurio basal de China, que confirmó que el grupo pertenecía a los terópodos celurosaurianos, y que las similitudes con los prosaurópodos habían evolucionado de forma independiente. Publicaron el primer cladograma que muestra las relaciones evolutivas de Therizinosauria y demostraron que Beipiaosaurus conservaba características de terópodos y celurosaurios más basales, lo que los vinculaba con los terizinosaurios. La preservación de estructuras parecidas a plumas en Beipiaosaurus También sugirió que esta característica se distribuyó más ampliamente entre los terópodos de lo que se pensaba anteriormente.[6]
A principios del siglo XXI, se habían descubierto muchos más taxones de terizinosaurios, incluidos algunos fuera de Asia, siendo el primero Nothronychus de América del Norte en 2001. Los taxones basales que ayudaron a iluminar la evolución temprana del grupo, como Falcarius en 2005, también fueron descubierto. Los terizinosaurios ya no se consideraban raros o aberrantes, sino más diversos en características, incluido el tamaño, de lo que se pensaba anteriormente y su clasificación como terópodos maniraptoranos fue generalmente aceptada.[33][34][35] La ubicación de Therizinosauria dentro de Maniraptora continuó sin estar clara, en 2017, Alan H. Turner y sus colegas encontraron que se agrupaban con oviraptorosaurios, mientras que en 2009 Zanno y sus colegas encontraron que eran el clado más basal dentro de Maniraptora, delimitado por Ornithomimosauria y Alvarezsauridae.[36][37] A pesar del material fósil adicional, las interrelaciones dentro del grupo también eran inciertas en 2010, cuando Zanno realizó el análisis filogenético más detallado de Therizinosauria hasta ese momento. Citó la inaccesibilidad, el daño, la pérdida potencial de especímenes de holotipos, la escasez de restos craneales y los especímenes fragmentarios con pocos elementos superpuestos como los obstáculos más importantes para resolver las relaciones evolutivas dentro del grupo. La posición de Segnosaurus y la de algunos otros terizinosauridos asiáticos se vio afectada por estos factores. Zanno afirmó que serían necesarios más especímenes bien conservados y que sería necesario redescubrir los elementos faltantes. Zanno también revisó Therizinosauroidea para excluir a Falcarius y lo retuvo en el clado más amplio Therizinosauria, que se convirtió en el sinónimo principal de Segnosauria.[8] Para 2015, Segnosaurus seguía siendo uno de los terizinosaurios más conocidos, según Christophe Hendrickx y sus colegas.[7]

El terizinosaurio más básico es el Falcarius del Cretácico Inferior de América del Norte. Mostró que la pelvis y la dentición fueron las primeras características que se modificaron del plan maniraptorano más general en los terizinosaurios, probablemente reflejando su transición de carnívoro a herbívoro.[35][38] Los terizinosaurios se conocen principalmente del Cretácico de Asia y América del Norte, y los posibles restos de otras épocas y lugares son controvertidos. Dado que se sabe que los terizinosaurios vivieron en el supercontinente Laurasia, que consistía en lo que ahora son América del Norte, Europa y Asia, Zanno sugirió dos escenarios para su distribución paleobiogeográfica en 2010. Una posibilidad es que se dispersaron a través de la vicarianza, por lo que los terizinosaurios estaban presentes en las áreas que se convirtieron en Asia y América del Norte antes de la ruptura que dividió estas áreas en el Triásico Tardío. La otra posibilidad es que los terizinosaurios basales se dispersaron entre Asia y América del Norte a través de Europa después del evento de ruptura pero antes del Barremiense medio, hace entre 132 y 138 millones de años, un puente terrestre temporal conectaba América del Norte y Europa, después de lo cual las masas terrestres se volvieron a aislar unas de otras, lo que explica por qué los terizinosaurios basales Beipiaosaurus de Asia y Falcarius de América del Norte eran morfológicamente divergentes entre sí, aunque coetáneos. La presencia del terizinosaurio derivado Nothronychus, que estaba más estrechamente relacionado con los géneros asiáticos, en América del Norte durante la etapa Turoniense del Cretácico superior temprano también muestra que habría habido un intercambio de fauna entre América del Norte y Asia a través de un puente de tierra antes de eso en el Cretácico temprano tardío durante el Aptiense o Albiense, que también se ve en algunos otros grupos de dinosaurios.[8]
Filogenia
El siguiente cladograma está basado en el análisis de Phil Senter et al., 2012.[39]
| Therizinosauroidea |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
El siguiente cladograma muestra las relaciones dentro de Therizinosauria según un estudio de 2013 de Hanyong Pu y sus colegas, que se basó en el análisis de Zanno de 2010, con la adición del género basal Jianchangosaurus. [40]
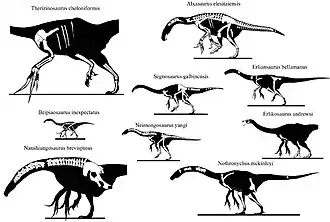
| Therizinosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiología
Generalmente se acepta que los tericinosáuridos fueron omnívoros o herbívoros, Mostrando características herbívoras como cuellos alargados, tamaño reducido del cráneo, ampliación de la cavidad de la pelvis, reducción de la fuerza de mordida, reducción de la musculatura del cráneo, La pérdida de dientes anteriores, el desarrollo de una rhamphotheca y una porción anterior del dentario, que están presentes o presumiblemente presentes en el Segnosaurus. Sin embargo, no ha habido complejidad en la morfología dental, que se encuentra en muchos otros grupos herbívoros. Algunas de las modificaciones dentales incluyen aumento de la simetría en Erlikosaurus , agrandamiento de los dentículos en Segnosaurus , con las únicas morfologías de dientes únicas previamente conocidas que son los dientes anteriores cúspides e incisivos de Falcarius y los dientes exteriores convexos de Jianchangosaurus .[4]
En 1979 y 1981, Barsbold y Perle dijeron que el metatarso corto y masivo y los dedos extendidos inusualmente grandes indicaban que Segnosaurus y sus parientes no estaban adaptados para la locomoción rápida, tal vez porque no era requerido por su estilo de vida. Barsbold y Perle sugirieron que podrían haber sido anfibios.[16][17] Barsbold y Maryańska acordaron en 1990 que las patas cortas y anchas y los troncos voluminosos del grupo indicaban que eran animales de movimiento lento.[26] Paul representó un esqueleto "segnosaurio" parecido a un prosaurópodo, un compuesto de varios géneros, en una postura cuadrúpedo en 1988.[23] Basado en los restos más completos de Alxasaurus y la articulación de su columna vertebral, Russell concluyó en 1993 que la restauración esquelética de Paul era inexacta y que los brazos de los terizinosaurios se mantenían despejados del suelo.[29] En 1995, Nessov sugirió que las garras alargadas de los terizinosaurios se usaban para defenderse de los depredadores y que sus crías podrían haber usado sus garras para la locomoción arbórea a lo largo de los troncos y las copas de los árboles de manera similar a los perezosos.[31]
En un estudio de 2012 de la anatomía endocraneal de Erlikosaurus y otros terizinosaurios que preservan las cajas cerebrales, Stephan Lautenschlager y sus colegas encontraron que estos dinosaurios tenían sentidos del olfato, el oído y el equilibrio bien desarrollados. Los dos primeros sentidos pueden haber jugado un papel en la búsqueda de comida, la evasión de los depredadores y el comportamiento social. Estos sentidos también estaban bien desarrollados en celurosaurios anteriores, por lo que los terizinosaurios pueden haber heredado estos rasgos de sus ancestros carnívoros y los usaron para diferentes propósitos dietéticos.[41] En un estudio de 2014 sobre la función de las garras de las manos de los terizinosaurios, Lautenschlager descubrió que estas no se habrían utilizado para cavar, lo que se habría hecho con las garras de los pies porque, como en otros maniraptoranos, las plumas de las extremidades anteriores habrían interferido con esta función. No pudo confirmar ni ignorar que las garras de la mano podrían haber sido utilizadas para la defensa, el combate, la estabilización al agarrarse de los troncos de los árboles durante el ramoneo, la exhibición sexual o el agarre de las parejas durante la cópula. Descartó en gran medida que cavaran madrigueras, debido a su tamaño.[42]

Los huevos de dinosaurio con embriones del tipo Dendroolithidae de la Formación Nanchao de China fueron identificados como pertenecientes a terizinosaurios y descritos por Martin Kundrát y sus colegas en 2007. El desarrollo de los embriones y el hecho de que no se encontraron adultos en asociación con los nidos indican que Las crías de terizinosaurios eran precoces, capaces de moverse desde el nacimiento y podían dejar sus nidos para alimentarse solos, independientemente de sus padres.[43][10] En un resumen de la conferencia de 2013, Yoshitsugu Kobayashi y sus colegas informaron un lugar de anidación de dinosaurios terópodos de la Formación Javkhlant de Mongolia que contenía al menos 17 nidadas de huevos.dentro de un área de 22 por 52 metros. Cada nidada contenía ocho huevos esféricos con superficies rugosas Los huevos estaban en contacto entre sí y dispuestos en una estructura circular sin abertura central. Los investigadores identificaron los huevos como dendroolítidos y, por lo tanto, como terizinosaurios. Aunque no se conocen terizinosaurios de la formación, se superpone a la Formación Bayan Shireh donde se encontraron Segnosaurus, Erlikosaurus y Enigmosaurus. Las múltiples nidadas indican que algunos terizinosaurios anidaban coloniales como hadrosáuridos, prosaurópodos, titanosaurianos y aves. Los huevos se encontraron en una sola capa estratigráfica, lo que sugiere que los dinosaurios anidaron en el sitio en una sola ocasión y no exhibieron fidelidad al sitio.[44]
Alimentación
Las características inusuales de los terizinosaurios han llevado a varias interpretaciones de su comportamiento de alimentación. No hay evidencia directa de su dieta, como el contenido del estómago y los rastros de alimentación. En 1970, Anatoly K. Rozhdestvensky sugirió que Therizinosaurus, el único miembro del grupo conocido en ese momento, usaba sus grandes garras para abrir montículos de termitas o recolectar frutos de los árboles.[10] Barsbold y Perle señalaron en 1979 y 1980 que sus características peculiares probablemente reflejaban una dirección evolutiva diferente a la de los terópodos más típicos, muchos de los cuales se consideraban depredadores activos y efectivos. Sus delicadas mandíbulas, dientes y picos pequeños y débiles, y patas cortas y compactas indicaban que no habrían usado el armamento de otros terópodos para conseguir comida, sino que podrían haber cazado peces.[16][9] En 1983, Barsbold dijo que el pico córneo en la parte delantera de las mandíbulas y los dientes debilitados en la parte posterior eran características comunes entre los dinosaurios herbívoros, pero no entre los terópodos carnívoros y especuló que esto podría indicar que los segnosaurios se habían convertido en herbívoros.[13] En 1984, Paul sugirió que eran herbívoros debido a las similitudes de sus cráneos con los de los dinosaurios prosaurópodos y ornitisquios, que incluyen picos córneos, filas de dientes insertados y un estante al lado de las mandíbulas que indica la presencia de mejillas. Como los ornitisquios, podían, por tanto, cosechar, manipular y masticar plantas de una manera sofisticada. También sugirió que el hueso ilíaco de la pelvis tenía hojas que se ensanchaban hacia los lados en la parte delantera similares a las de los saurópodos para sostener un intestino grande que se usaba para fermentar y procesar alimentos.[21] Norman declaró en 1985 que la posibilidad de que Segnosaurus fuera un devorador de peces acuáticos pudiera explicar sus dientes pequeños y puntiagudos y sus pies anchos y quizás palmeados, pero encontró misterioso por qué debería tener un pico córneo.[22]
En 1993, Russell y Dong consideraron que el pequeño tamaño de la cabeza, los picos romos y los grandes pesos corporales de los terizinosaurios eran compatibles con la herbivoría.[27] En 1993 y 1997, Russell sugirió que los terizinosaurios se habrían "sentado" sobre sus pelvis y apoyado sus cuerpos sobre sus extremidades traseras mientras usaban sus largos brazos, garras y cuellos flexibles para alcanzar las hojas de los árboles y arbustos con sus picos. Podrían haber llegado aún más alto mientras estaban de pie y navegando bípedo. Esto es paralelo a la forma en que algunos mamíferos herbívoros usan sus extremidades anteriores para manipular la vegetación; Russell consideró que los calicoterios y los perezosos terrestres extintos , así como los gorilas, convergen adaptativamente con los terizinosaurios. Debido a que los restos de terizinosaurios se encuentran a menudo ensedimentos depositados en entornos de ríos y lagos, Russell dijo que pueden haber ramoneado en arbustos y árboles ribereños.[28][29] Basado en el ensamblaje de fósiles en la Formación Bissekty de Uzbekistán, Nessov sugirió en 1995 que los terizinosaurios podrían haber sido parte de sus ecosistemas acuáticos ricos en nutrientes, aunque quizás indirectamente, al alimentarse de avispas que se habían alimentado de carroña de vertebrados acuáticos. Encontró esto consistente con la sugerencia de Rozhdestvensky de que los terizinosaurios pueden haberse alimentado de insectos sociales.[31] En un resumen de la conferencia de 2006, Sara Burch presentó el rango de movimiento inferido en los brazos del terizinosaurio Neimongosaurus y concluyó que el movimiento general en la articulación glenoide-humeral en el hombro era aproximadamente circular, y se dirigía hacia los lados y ligeramente hacia abajo, que divergía de los rangos más ovalados, dirigidos hacia atrás y hacia abajo de otros terópodos. Esta capacidad de extender los brazos considerablemente hacia adelante puede haber ayudado a los terizinosaurios a alcanzar y agarrar el follaje.[45]

En 2009, Zanno y sus colegas afirmaron que los terizinosaurios eran los candidatos más considerados para la herbivoría entre los terópodos y enumeraron las características asociadas con esta dieta. Estos incluían estrías pequeñas, densamente empaquetadas y gruesas, dientes lanceolados, en forma de lanza, con una baja tasa de reemplazo, un pico en la parte delantera de las mandíbulas, una fila de dientes incrustada que sugiere mejillas carnosas, un cuello alargado, un pequeño cráneo, una capacidad intestinal muy grande, como lo indica la circunferencia de las costillas en el tronco y los procesos de abocinamiento hacia afuera del hueso ilíaco y la pérdida de adaptaciones cursoriales en las extremidades traseras, incluido el desarrollo de patas funcionalmente tetradáctilos. Zanno y sus colegas encontraron que los clados en la base de Maniraptora, Ornithomimosauria, Therizinosauria y Oviraptorosauria tenían evidencia directa o morfológica de herbivoría, lo que significaría que esta dieta evolucionó de forma independiente varias veces en terópodos celurosaurianos o que la condición primitiva del grupo era herbivoría al menos facultativa con la carnivoría solo emergiendo en maniraptoranos más derivados.[37] Zanno y Peter J. Makovicky encontraron, en 2011, los terizinosaurios y algunos otros grupos de dinosaurios herbívoros que tenían picos y dientes retenidos no podían perder sus dientes por completo porque carecían de molinos gástricos, mollejas, y necesitaban los dientes para procesar los alimentos, y que la dieta folívora rica en fibra, basada en hojas, de los terizinosaurios y otros arcosaurios también puede haber impedido la evolución de un pico completo.[46] Lautenschlager descubrió en 2014 que las manos de los terizinosaurios debían poder extender el rango del animal hasta un punto que no podría alcanzar la cabeza si se usaran para ramonear y derribar vegetación. Sin embargo, en los géneros en los que se conservan los elementos del cuello y las extremidades anteriores, los cuellos tenían la misma longitud o eran más largos que las extremidades anteriores, por lo que arrancar la vegetación solo tendría sentido si las partes inferiores de las ramas largas se tiraran hacia abajo para acceder a las partes fuera del alcance de árboles.[42]
Zanno y sus colegas declararon en 2016 que en general se aceptaba que los terizinosaurios se encontraban dentro del espectro de omnivoría y herbivoría, con una tendencia hacia la herbivoría intensificada. Si bien se han utilizado varias características anatómicas para apoyar esta idea, la morfología de los dientes se ha considerado relativamente simplista y con pocas especializaciones únicas en comparación con otros dinosaurios herbívoros. Las pocas modificaciones incluyen el aumento de la simetría en los dientes de Erlikosaurus y el agrandamiento de los dentículos en Segnosaurus. Zanno y sus colegas identificaron características novedosas y complejas en los dientes dentarios de Segnosaurus, incluida la presencia de carinae adicional y carinae plegada con bordes frontales denticulados, que indican Segnosaurus tenía un mayor grado de procesamiento de alimentos por vía oral que otros terizinosaurios. Estos rasgos juntos crearon una superficie rugosa y triturada cerca de la base de las coronas de los dientes que era exclusiva de Segnosaurus y sugiere que consumía recursos alimenticios únicos o utilizaba estrategias de alimentación altamente especializadas. Debido a que múltiples formaciones geológicas muestran evidencia de especies de terizinosaurios simpátricos, especies relacionadas que vivieron en la misma área al mismo tiempo, es posible que la partición de nichos entre ellas haya jugado un papel en el éxito evolutivo del grupo. Esto es apoyado por Segnosaurus con su dentición altamente especializada siendo contemporánea de Erlikosaurus, con sus dientes relativamente indistintos, lo que indica su división en la adquisición, procesamiento o recursos de alimentos. Esta conclusión también se ve reforzada por la gran diferencia en las masas corporales estimadas de los terizinosaurios simpátricos en la Formación Bayan Shireh, que fue de hasta un 500%.[4]
En un estudio de 2017 sobre la partición de nichos en terizinosaurios a través de simulaciones digitales, Lautenschlager descubrió que los dentarios de Segnosaurus experimentaron una de las magnitudes de estrés más bajas durante los escenarios de alimentación extrínseca. Segnosaurus y Erlikosaurus fueron ayudados por la punta hacia abajo de la mandíbula inferior y las regiones sinfisarias, y probablemente también por los picos, que se sabe que mitigan el estrés y la tensión. Por el contrario, los dentarios más rectos y alargados de los terizinosaurios basales, típicos de sus ancestros celurosaurios, tenían las mayores magnitudes de estrés y tensión. Un movimiento de la cabeza hacia abajo mientras agarra la vegetación era más probable que un movimiento hacia los lados o hacia arriba, aunque tal comportamiento sería más probable en Segnosaurus y Erlikosaurus con sus mandíbulas que mitigan el estrés. La diferencia en la fuerza de mordida relativa entre el Segnosaurus y el simpátrico Erlikosaurus muestra que el primero habría podido alimentarse de una vegetación más dura, mientras que la robustez general del segundo sugiere una mayor flexibilidad en su forma de alimentarse, porque los niveles de estrés se mantuvieron bajos en las simulaciones de alimentación. Lautenschlager estuvo de acuerdo en que los dos taxones se adaptaron a diferentes modos de alimentación y selección de alimentos. Segnosaurus estaba más adaptado a usar su dentición para obtener o procesar alimentos, mientras que Erlikosaurus usaba principalmente su pico para cortar y la musculatura de su cuello mientras buscaba alimento. La diferencia de tamaño entre Segnosaurus y Erlikosaurus, el primero de los cuales se estima que pesó más que el segundo, indica que estos efectos se incrementaron y que hubo más mecanismos que dividían sus recursos, como diferentes alturas. Debido a que otros taxones de terizinosaurios estaban más divididos en tiempo y espacio, otros factores además de la competencia dentro de su grupo también pueden haber contribuido a su variación, como las adaptaciones a diferentes flora y la competencia con otros tipos de herbívoros.[47]
En 2018, Loredana Macaluso y sus colegas señalaron que las caderas de los terizinosaurios eran peculiares porque el eje del hueso púbico se giraba hacia atrás, mientras que la bota púbica se proyectaba fuertemente hacia adelante. Si bien el intestino más grande asociado con la herbivoría fue capaz de empujar el eje hacia atrás, sugirieron que la bota púbica estaba restringida por músculos ventilatorios que eran cruciales para la ventilación corasal, respirar con sacos de aire adicionales, lo que muestra la importancia de este modo de respiración.[48] En un estudio de 2019 sobre la musculatura de la mandíbula, Ali Nabavizadeh concluyó que los terizinosaurios se alimentaban principalmente de ortigas, movían la mandíbula hacia arriba y hacia abajo, y levantaban la mandíbula de forma isognata, por lo que los dientes superiores e inferiores de cada lado se ocluían,se pusieron en contacto entre sí, a la vez. El origen y los sitios de inserción de los músculos de la mandíbula también agregaron fuerza al cierre de la mandíbula..[49] David J. Button y Zanno encontraron en 2019 dinosaurios herbívoros que siguieron principalmente dos modos distintos de alimentación, ya sea procesando alimentos en el intestino, caracterizado por cráneos gráciles y bajas fuerzas de mordida, o la boca, caracterizada por características asociadas con un procesamiento extensivo. Segnosaurus, junto con los saurópodos diplodocoides y titanosaurianos, los ornitomimosaurios deinoqueíridos y ornitomimidos y los caenagnátidos, se encontraban en la primera categoría, mientras que Erlikosaurus era más similar a algunos taxones de sauropodomorfos y ornitisquios, lo que indica que estos dos terizinosaurios estaban funcionalmente separados y ocupaban diferentes nichos.[50]
Paleoecología

Se han recuperado fósiles de Segnosaurus de la Formación Bayan Shireh en Mongolia, que data de hace unos 102-86 millones de años durante las etapas Cenomaniense a Turoniense del período Cretácico Tardío, basándose en análisis paleomagnéticos y mediciones de calcita U-Pb.[51][52] Los restos se encontraron en arenas grises mal cementadas que contienen conglomerados intraformacionales, grava y arcillas grises.[26] La Formación Bayan Shireh se superpone a la Formación Baruunbayan y subyace a la Formación Javkhlant. Los sedimentos de estas formaciones fueron depositados por ríos serpenteantes y lagos en una llanura aluvial,terreno llano formado por sedimentos depositados por ríos de la sierra, con un clima semiárido.[53][54]
Los terizinosaurios fueron los terópodos más abundantes en la Formación Bayan Shireh en términos de biodiversidad. Además de Segnosaurus, los miembros del grupo incluían Erlikosaurus, Enigmosaurus y posiblemente un cuarto tipo.[54][55] Otros terópodos incluyeron al tiranosáurido Alectrosaurus, el ornitomimido Garudimimus y el dromeosáurido Achillobator.[51][56] Otros dinosaurios incluyeron el anquilosáurido Talarurus,[51] el hadrosáurido Gobihadros,[57] el saurópodo Erketu,[58] y el ceratopsiano Graciliceratops.[59] También se han encontrado huevos de dinosaurio, algunos de los cuales fueron identificados como Dendroolithidae, así como huellas de dinosaurios y crocodiliformes . La formación se distingue por su variedad y abundancia de tortugas, y los invertebrados incluyen ostrácodos y moluscos de agua dulce.[51][26][54] La Formación Bayan Shireh es posiblemente coetánea con la Formación Iren Dabasu de la región de Mongolia Interior de China, de donde también se han encontrado fósiles de terizinosaurios similares a los de Segnosaurus y Erlikosaurus.[60]
Referencias
- Perle, A., 1979, Segnosauridae-a new family of theropods from the Late Cretaceous of Mongolia: Joint Soviet-Mongolian Paleontological Expeditions, Transactions v. 8, p. 45-55.
- Paul, G.S. (2016). The Princeton Field Guide to Dinosaurs (2 edición). Princeton: Princeton University Press. pp. 162, 166-167. ISBN 978-0-691-16766-4.
- Holtz, T. R. Jr. (2012). Dinosaurs: The Most Complete, Up-to-date Encyclopedia for Dinosaur Lovers of All Ages. New York: Random House. p. 382. ISBN 978-0-375-82419-7.
- Zanno, L.E.; Tsogtbaatar, K.; Chinzorig, T.; Gates, T.A. (2016). «Specializations of the mandibular anatomy and dentition of Segnosaurus galbinensis (Theropoda: Therizinosauria)». PeerJ 4: e1885. doi:10.7717/peerj.1885.
- Campione, Nicolás E.; Evans, David C. (2020). «The accuracy and precision of body mass estimation in non-avian dinosaurs». Biological Reviews (en inglés) 95 (6): 1759-1797. ISSN 1469-185X. PMID 32869488. S2CID 221404013. doi:10.1111/brv.12638.
- Xu, X.; Tang, Z.; Wang, X. (1999). «A therizinosauroid dinosaur with integumentary structures from China». Nature 399 (6734): 350-354. Bibcode:1999Natur.399..350X. S2CID 204993327. doi:10.1038/20670.
- Hendrickx, C.; Hartman, S. A.; Mateus, O. (2015). «An overview on non-avian theropod discoveries and classification». PalArch's Journal of Vertebrate Palaeontology 12 (1): 27-29. Archivado desde el original el 22 de junio de 2018.
- Lindsay E. Zanno (2010). «A taxonomic and phylogenetic re-evaluation of Therizinosauria (Dinosauria: Maniraptora)». Journal of Systematic Palaeontology 8 (4): 503-543. doi:10.1080/14772019.2010.488045.
- Barsbold, R.; Perle, A. (1980). «Segnosauria, a new infraorder of carnivorous dinosaurs». Acta Palaeontologica Polonica 25 (2): 187-195.
- Clark, J. M.; Maryańska, T.; Barsbold, R. (2004). «Therizinosauroidea». En Weishampel, D. B.; Dodson, P.; Osmolska, H., eds. The Dinosauria (2 edición). Berkeley: University of California Press. pp. 151–164. ISBN 978-0-520-24209-8.
- Barsbold, R. (1979). «Opisthopubic pelvis in the carnivorous dinosaurs». Nature 279 (5716): 792-793. Bibcode:1979Natur.279..792B. S2CID 4348297. doi:10.1038/279792a0.
- Perle, A. (1979). «Segnosauridae - novoe semeistvo teropod is posdnego mela Mongolii». Trdy - Sovmestnaya Sovetsko-Mongol'skaya Paleontologicheskaya Ekspeditsiya 8: 45-55.
- Barsbold, R. (1983). «Khishchnye dinosavry mela Mongoliy» [Carnivorous dinosaurs from the Cretaceous of Mongolia]. Transactions of the Joint Soviet-Mongolian Paleontological Expedition (en russian) 19: 51-52, 76, 108-109. Archivado desde el original el 13 de marzo de 2016. Consultado el 21 de septiembre de 2019. Parámetro desconocido
|translator2-last=ignorado (ayuda); Parámetro desconocido|translator2-first=ignorado (ayuda); Parámetro desconocido|translator-first=ignorado (ayuda); Parámetro desconocido|translator-last=ignorado (ayuda) - Zanno, L. E. (2006). «The pectoral girdle and forelimb of the primitive therizinosauroid Falcarius utahensis (Theropoda, Maniraptora): analyzing evolutionary trends within Therizinosauroidea». Journal of Vertebrate Paleontology 26 (3): 636. doi:10.1671/0272-4634(2006)26[636:TPGAFO]2.0.CO;2.
- Holtz, T. R. (2000). «Theropods». En Farlow, J. O.; Brett-Surman, M. K., eds. The Complete Dinosaur. Bloomington: Indiana University Press. p. 404. ISBN 978-0-253-21313-6.
- Barsbold, R.; Perle, A. (1979). «Modiphikatsiy tasa sayrisziy i parallelinoe rasvitie zishchnich dinosavrov» [Modification in the saurischian pelvis and the parallel development of predatory dinosaurs]. Transactions of the Joint Soviet Mongolian Paleontological Expedition (en russian) 8: 39-44. Parámetro desconocido
|translator-first=ignorado (ayuda); Parámetro desconocido|translator-last=ignorado (ayuda) - Perle, A. (1981). «Noviy segnozavrid iz verchnego mela Mongolii» [New Segnosauridae from the Upper Cretaceous of Mongolia]. Trudy – Sovmestnaya Sovetsko-Mongol'skaya Paleontologicheskaya Ekspeditsiya (en russian) 15: 50-59. Parámetro desconocido
|translator-first=ignorado (ayuda); Parámetro desconocido|translator-last=ignorado (ayuda); Parámetro desconocido|translator2-last=ignorado (ayuda); Parámetro desconocido|translator2-first=ignorado (ayuda) - Perle, A. (1982). «A hind limb of Therizinosaurus from the Upper Cretaceous of Mongolia». Problems in Mongolian Geology (en russian) 5: 94-98. Parámetro desconocido
|translator-first=ignorado (ayuda); Parámetro desconocido|translator-last=ignorado (ayuda) - Lee, Y. N.; Barsbold, R.; Currie, P. J.; Kobayashi, Y.; Lee, H. J.; Godefroit, P.; Escuillié, F. O.; Chinzorig, T. (2014). «Resolving the long-standing enigmas of a giant ornithomimosaur Deinocheirus mirificus». Nature 515 (7526): 257-260. Bibcode:2014Natur.515..257L. PMID 25337880. S2CID 2986017. doi:10.1038/nature13874.
- Barsbold, R. (1983). «O ptich'ikh chertakh v stroyenii khishchnykh dinozavrov» [“Avian” features in the morphology of predatory dinosaurs]. Transactions of the Joint Soviet Mongolian Paleontological Expedition (en russian) 24: 96-103. Parámetro desconocido
|translator-first=ignorado (ayuda); Parámetro desconocido|translator-last=ignorado (ayuda) - Paul, G. S. (1984). «The segnosaurian dinosaurs: relics of the prosauropod-ornithischian transition?». Journal of Vertebrate Paleontology 4 (4): 507-515. ISSN 0272-4634. JSTOR 4523011. doi:10.1080/02724634.1984.10012026.
- Norman, D. B. (1985). «Mischellaneous theropods». The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom. New York: Crescent Books. pp. 52-53. ISBN 978-0-517-46890-6.
- Paul, G. S. (1988). Predatory Dinosaurs of the World. New York: Simon & Schuster. pp. 185, 283. ISBN 978-0-671-61946-6.
- Gauthier, J. (1986). «Saurischian monophyly and the origin of birds». Memoirs of the California Academy of Sciences 8: 45. Archivado desde el original el 16 de agosto de 2019.
- Sereno, P. (1989). Prosauropod monophyly and basal sauropodomorph phylogeny. «Abstract of Papers. Forty-Ninth Annual Meeting Society of Vertebrate Paleontology». Journal of Vertebrate Paleontology 9 (3 Supplement): 39A. ISSN 0272-4634. JSTOR 4523276.
- Barsbold, R.; Maryańska, T. (1990). «Saurischia Sedis Mutabilis: Segnosauria». En Weishampel, D. B.; Osmolska, H.; Dodson, P., eds. The Dinosauria (1st edición). Berkeley: University of California Press. pp. 408-415. ISBN 978-0-520-06727-1.
- Russell, D. A.; Dong, Z.-M. (1993). «The affinities of a new theropod from the Alxa Desert, Inner Mongolia, People's Republic of China». Canadian Journal of Earth Sciences 30 (10): 2107-2127. Bibcode:1993CaJES..30.2107R. doi:10.1139/e93-183.
- Russell, D. A. (1997). «Therizinosauria». En Currie, Philip J.; Padian, Kevin, eds. Encyclopedia of Dinosaurs. San Diego: Academic Press. pp. 729–730. ISBN 978-0-12-226810-6.
- Russell, D. A.; Russell, D. E. (1993). «Mammal-dinosaur convergence». National Geographic Research 9: 70-79. ISSN 8755-724X.
- Clark, J. M.; Altangerel, P.; Norell, M. (1994). «The skull of Erlicosaurus andrewsi, a late Cretaceous "segnosaur" (Theropoda, Therizinosauridae) from Mongolia». American Museum Novitates (3115): 2, 32-36. hdl:2246/3712.
- Nessov, L. A. (1995). Dinosaurs of northern Eurasia: new data about assemblages, ecology and paleobiogeography. Saint Petersburg: Saint Petersburg State University. pp. 13, 18, 45-46. Parámetro desconocido
|translator-first=ignorado (ayuda); Parámetro desconocido|translator-last=ignorado (ayuda) - Holtz, T. R. (1996). «Phylogenetic taxonomy of the Coelurosauria (Dinosauria: Theropoda)». Journal of Paleontology 70 (3): 536-538. ISSN 0022-3360. JSTOR 1306452. doi:10.1017/S0022336000038506.
- Kirkland, James I.; Wolfe, Douglas G. (2001). «First definitive therizinosaurid (Dinosauria; Theropoda) from North America». Journal of Vertebrate Paleontology 21 (3): 410-414. ISSN 0272-4634. JSTOR 20061971. doi:10.1671/0272-4634(2001)021[0410:FDTDTF]2.0.CO;2.
- Zhang, X.-H.; Xu, X.; Zhao, Z.-J.; Sereno, P.; Kuang, X.-W.; Tan, L. (2001). «A long-necked therizinosauroid dinosaur from the Upper Cretaceous Iren Dabasu Formation of Nei Mongol, People's Republic of China». Vertebrata PalAsiatica 39 (4): 282-290. Archivado desde el original el 4 de marzo de 2016.
- Kirkland, J. I.; Zanno, L. E.; Sampson, S. D.; Clark, J. M.; DeBlieux, D. D. (2005). «A primitive therizinosauroid dinosaur from the Early Cretaceous of Utah». Nature 435 (7038): 84-87. Bibcode:2005Natur.435...84K. PMID 15875020. S2CID 4428196. doi:10.1038/nature03468.
- Turner, A. H.; P., D.; Clarke, J. A.; Erickson, G. M.; Norell, M. A. (2007). «A basal dromaeosaurid and size evolution preceding avian flight». Science 317 (5843): 1378-1381. Bibcode:2007Sci...317.1378T. PMID 17823350. doi:10.1126/science.1144066. Parámetro desconocido
|doi-access=ignorado (ayuda) - Zanno, L. E.; Gillette, D. D.; Albright, L. B.; Titus, A. L. (2009). «A new North American therizinosaurid and the role of herbivory in 'predatory' dinosaur evolution». Proceedings: Biological Sciences 276 (1672): 3505-3511. ISSN 0962-8452. JSTOR 30244145. PMC 2817200. PMID 19605396. doi:10.1098/rspb.2009.1029.
- Zanno, L. E. (2010). «Osteology of Falcarius utahensis (Dinosauria: Theropoda): characterizing the anatomy of basal therizinosaurs». Zoological Journal of the Linnean Society 158 (1): 225. doi:10.1111/j.1096-3642.2009.00464.x. Parámetro desconocido
|doi-access=ignorado (ayuda) - Senter, P.; Kirkland, J. I.; Deblieux, D. D. (2012). «Martharaptor greenriverensis, a New Theropod Dinosaur from the Lower Cretaceous of Utah». En Dodson, Peter, ed. PLoS ONE 7 (8): e43911. PMC 3430620. PMID 22952806. doi:10.1371/journal.pone.0043911.
- Pu, H.; Kobayashi, Y.; Lü, J.; Xu, L.; Wu, Y.; Chang, H.; Zhang, J.; Jia, S. (2013). «An unusual basal therizinosaur dinosaur with an ornithischian dental arrangement from Northeastern China». En Claessens, Leon, ed. PLOS ONE 8 (5): e63423. Bibcode:2013PLoSO...863423P. PMC 3667168. PMID 23734177. doi:10.1371/journal.pone.0063423. Parámetro desconocido
|doi-access=ignorado (ayuda) - Lautenschlager, S.; Rayfield, E. J.; Altangerel, P.; Zanno, L. E.; Witmer, L. M.; Butler, R. J. (2012). «The endocranial anatomy of Therizinosauria and its implications for sensory and cognitive function». PLOS ONE 7 (12): e52289. Bibcode:2012PLoSO...752289L. PMC 3526574. PMID 23284972. doi:10.1371/journal.pone.0052289. Parámetro desconocido
|doi-access=ignorado (ayuda) - Lautenschlager, S. (2014). «Morphological and functional diversity in therizinosaur claws and the implications for theropod claw evolution». Proceedings of the Royal Society B: Biological Sciences 281 (1785): 20140497. PMC 4024305. PMID 24807260. doi:10.1098/rspb.2014.0497.
- Kundrát, M.; Cruickshank, A. R. I.; Manning, T. W.; Nudds, J. (2007). «Embryos of therizinosauroid theropods from the Upper Cretaceous of China: diagnosis and analysis of ossification patterns». Acta Zoologica 89 (3): 231-251. doi:10.1111/j.1463-6395.2007.00311.x.
- «First record of a dinosaur nesting colony from Mongolia reveals nesting behavior of therizinosauroids». Hokkaido University. 2013. Archivado desde el original el 21 de diciembre de 2019. Consultado el 21 de diciembre de 2019.
- Burch, S. (2006). The range of motion of the glenohumeral joint of the therizinosaur Neimongosaurus yangi (Dinosauria: Theropoda). Chicago Biological Investigator 3 (2). p. 20.
- Zanno, L. E.; Makovicky, P. J. (2011). «Herbivorous ecomorphology and specialization patterns in theropod dinosaur evolution». Proceedings of the National Academy of Sciences 108 (1): 232-237. Bibcode:2011PNAS..108..232Z. PMC 3017133. PMID 21173263. doi:10.1073/pnas.1011924108. Parámetro desconocido
|doi-access=ignorado (ayuda) - Lautenschlager, S. (2017). «Functional niche partitioning in Therizinosauria provides new insights into the evolution of theropod herbivory». Palaeontology 60 (3): 375-387. doi:10.1111/pala.12289.
- Macaluso, L.; Tschopp, E.; Mannion, P. (2018). «Evolutionary changes in pubic orientation in dinosaurs are more strongly correlated with the ventilation system than with herbivory». Palaeontology 61 (5): 703-719. doi:10.1111/pala.12362.
- Nabavizadeh, A. (2019). «Cranial musculature in herbivorous dinosaurs: a survey of reconstructed anatomical diversity and feeding mechanisms». The Anatomical Record 303 (4): 1104-1145. PMID 31675182. S2CID 207815224. doi:10.1002/ar.24283.
- Button, D. J.; Zanno, L. E. (2019). «Repeated evolution of divergent modes of herbivory in non-avian dinosaurs». Current Biology 30 (1): 158-168.e4. PMID 31813611. S2CID 208652510. doi:10.1016/j.cub.2019.10.050. Parámetro desconocido
|doi-access=ignorado (ayuda) - Hicks, J. F.; Brinkman, D. L.; Nichols, D. J.; Watabe, M. (1999). «Paleomagnetic and palynologic analyses of Albian to Santonian strata at Bayn Shireh, Burkhant, and Khuren Dukh, eastern Gobi Desert, Mongolia». Cretaceous Research 20 (6): 829-850. doi:10.1006/cres.1999.0188.
- Kurumada, Y.; Aoki, S.; Aoki, K.; Kato, D.; Saneyoshi, M.; Tsogtbaatar, K.; Windley, B. F.; Ishigaki, S. (2020). «Calcite U–Pb age of the Cretaceous vertebrate‐bearing Bayn Shire Formation in the Eastern Gobi Desert of Mongolia: usefulness of caliche for age determination». Terra Nova 32 (4): 246-252. Bibcode:2020TeNov..32..246K. doi:10.1111/ter.12456. Parámetro desconocido
|doi-access=ignorado (ayuda) - Khand, Y.; Badamgarav, D.; Ariunchimeg, Y.; Barsbold, R. (2000). «Cretaceous system in Mongolia and its depositional environments». Developments in Palaeontology and Stratigraphy (Elsevier) 17: 49-79. ISBN 9780444502766. doi:10.1016/S0920-5446(00)80024-2.
- Lee, Yu.-N.; Lee, H.-J.; Kobayashi, Y.; Paulina-Carabajal, A.; Barsbold, R.; Fiorillo, A. R.; Tsogtbaatar, K. (2019). «Unusual locomotion behaviour preserved within a crocodyliform trackway from the Upper Cretaceous Bayanshiree Formation of Mongolia and its palaeobiological implications». Palaeogeography, Palaeoclimatology, Palaeoecology 533: 109239. Bibcode:2019PPP...533j9239L. S2CID 197584839. doi:10.1016/j.palaeo.2019.109239.
- Kobayashi, Y.; Tsogtbaatar, C.; Tsogtbaatar, K.; Barsbold, R. (2015). «A new therizinosaur with functionally didactyl hands from the Bayanshiree Formation (Cenomanian-Turonian), Omnogovi Province, southeastern Mongolia». Journal of Vertebrate Paleontology (Dallas): 157.
- Perle, A.; Norell, M. A.; Clark, J. (1999). «A new maniraptoran theropod – Achillobator giganticus (Dromaeosauridae) – from the Upper Cretaceous of Burkhant, Mongolia». Contributions from the Geology and Mineralogy Chair, National Museum of Mongolia (101): 1-105. OCLC 69865262.
- Tsogtbaatar, K.; Weishampel, D. B.; Evans, D. C.; Watabe, M.; Mpodozis, J. M. (2019). «A new hadrosauroid (Dinosauria: Ornithopoda) from the Late Cretaceous Baynshire Formation of the Gobi Desert (Mongolia)». PLOS ONE 14 (4): e0208480. Bibcode:2019PLoSO..1408480T. PMC 6469754. PMID 30995236. doi:10.1371/journal.pone.0208480. Parámetro desconocido
|doi-access=ignorado (ayuda) - Ksepka, D. T.; Norell, M. (2006). «Erketu ellisoni, a long-necked sauropod from Bor Guvé (Dornogov Aimag, Mongolia)». American Museum Novitates (3508): 1-16. doi:10.1206/0003-0082(2006)3508[1:EEALSF]2.0.CO;2. hdl:2246/5783.
- Sereno, P. C. (2000). «The fossil record, systematics and evolution of pachycephalosaurs and ceratopsians from Asia». En Benton, M. J.; Shishkin, M. A.; Unwin, D. M. et al., eds. The Age of Dinosaurs in Russia and Mongolia. Cambridge: Cambridge University Press. pp. 480–516. ISBN 978-0-521-54582-2.
- Currie, P. J.; Eberth, D. A. (1993). «Palaeontology, sedimentology and palaeoecology of the Iren Dabasu Formation (Upper Cretaceous), Inner Mongolia, People's Republic of China». Cretaceous Research 14 (2): 127-144. doi:10.1006/cres.1993.1011.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikispecies tiene un artículo sobre Segnosaurus galbinensis.
Wikispecies tiene un artículo sobre Segnosaurus galbinensis. Wikimedia Commons alberga una categoría multimedia sobre Segnosaurus galbinensis.
Wikimedia Commons alberga una categoría multimedia sobre Segnosaurus galbinensis.
| Control de autoridades |
|
|---|
 Datos: Q131102
Datos: Q131102- Multimedia: Segnosaurus / Q131102
- Especies: Segnosaurus