Hadrosauridae
Los hadrosáuridos (Hadrosauridae, gr.: ἁδρός , hadrós, "corpulento, grueso"), es una familia de dinosaurios ornitópodos hadrosauroideos. Este grupo también es conocido como "dinosaurios pico de pato", por la semejanza de su boca con los picos de las aves anátidas. La familia, que incluye a géneros como Edmontosaurus y Parasaurolophus, eran herbívoros comunes durante el Cretácico tardío y se distribuyeron en lo que hoy es Antártida, Asia, Europa, Norteamérica y Sudamérica.[7]
| Hadrosauridae | ||
|---|---|---|
| Rango temporal: 86 Ma - 66 Ma Cretácico Superior | ||
 Esqueleto montado de un Hadrosaurus. | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Ornithischia | |
| Suborden: | Neornithischia | |
| Infraorden: | Ornithopoda | |
| Superfamilia: | Hadrosauroidea | |
| Familia: |
Hadrosauridae Cope, 1870[1] | |
| Subfamilias | ||
| Sinonimia | ||
Los hadrosáuridos son descendientes dinosaurios iguanodontianos del Jurásico tardío y Cretácico temprano con los cuales comparten muchas características morfológicas. Como el resto de los ornitisquios, estos animales tenían un hueso predentario, así como el pubis señala hacia abajo y hacia atrás (hacia la cola), paralelo al isquion.[8] Los hadrosáuridos se dividen en dos subfamilias principales: los Lambeosaurinae o lambeosaurinos, que tenían crestas huecas craneales o tubos, y los Saurolophinae o saurolofinos, (identificados como hadrosaurinos en la mayoría de publicaciones anteriores al 2010)[9] que carecían de crestas craneales huecas (crestas óseas sólidas estaban presentes en algunos géneros).[10] Saurolofinos tendían a ser más voluminosos que lambeosaurinos. Los lambeosaurinos se dividen en 4 tribus, los aralosaurinis, lambeosaurinis, parasaurolophinis y tsintaosaurinis; mientras saurolofinos incluyen los brachylophosaurinis, edmontosaurinis, kritosaurinis y saurolofinis.[10]
Los hadrosáuridos eran bípedos facultativos, los jóvenes de algunas especies caminaban mayor parte de tiempo en dos patas y los adultos tendían hacerlo primordialmente en cuatro.[11][12] Sus mandíbulas estaban diseñadas para la molienda de las plantas, con múltiples hileras de dientes que se sustituían entre con los dientes que iban debajo, los cuales se remplazaban continuamente.[13]
Descripción

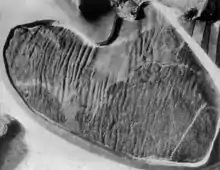
El aspecto más reconocible de la anatomía de los hadrosáuridos, son los huesos rostrales aplanados y alargados lateralmente, lo que da la apariencia del pico distintivo de los patos, asimismo, algunos géneros de hadrosáuridos también tenían grandes crestas en la cabeza, probablemente para su exhibición,[10] aunque la función de esta cresta es aún materia de debate. Otra hipótesis que cuenta con más consenso es que pudiera servir de caja de resonancia para comunicarse con otros miembros de la misma especie.[14] En algunos géneros, incluido el género Edmontosaurus, la mayor parte frontal del cráneo era plana y se ensanchaba para formar un pico,[15] que era ideal para recortar las hojas y ramas de los bosques de Asia, Europa y América del Norte. Sin embargo, la parte posterior de la boca contenía miles de dientes adecuados para moler alimentos antes de ingerirlos. Esta hipótesis postula que esta característica pudo haber sido un factor crucial en el éxito de este grupo en el Cretácico en comparación con los saurópodos.[13]
Se han encontrado múltiples impresiones de la piel de hadrosáuridos. A partir de estas impresiones, se ha podido determinar que los hadrosáuridos estaban recubiertos de escamas y no tenían plumas como dinosaurios de otros grupos. Asimismo, las impresiones de la piel de estos animales podrían permitir diferenciar ciertas especies o géneros.[16]
Los hadrosáuridos eran dinosaurios herbívoros de muy variados tamaños. Algunos géneros, como el Shantungosaurus y el Magnapaulia, alcabann con facilidad los 12 metros.[17][18][19] Por otro lado, géneros como el Arenysaurus, tenían una longitud media que oscilaba entre los 5 y los 6 metros.[20]
Diferencias craneales
 Cráneo de saurolofino Gryposaurus notabilis.
Cráneo de saurolofino Gryposaurus notabilis. Cráneo de lambeosaurino Hypacrosaurus altispinus.
Cráneo de lambeosaurino Hypacrosaurus altispinus. Cráneo del saurolofino Edmontosaurus annectens.
Cráneo del saurolofino Edmontosaurus annectens. Cráneo del lambeosaurino Lambeosaurus lambei.
Cráneo del lambeosaurino Lambeosaurus lambei.
Algunas especies de hadrosáuridos, como por ejemplo los lambeosaurinos, poseían una cresta ósea hueca y alta, elaborada encima de su cráneo, que contenía canales nasales alargados.[14][21] Por otro lado, los |saurolofinos tenían narices anchas, con una probable semejanza a la función de las crestas del otro grupo, la cual era comunicación o exihibición. La mayoría de estos últimos tenían la cabeza plana, sin embargo. un descubrimiento reciente, demuestra que Edmontosaurus regalis tenía una cresta compuesta de queratina,[22] lo que demuestra que probablemente otros miembros tanto de la tribu Edmontosaurini y la familia Saurolophinae también pudieran desarrollar este tipo de cresta, aunque los géneros Augustynolophus, Brachylophosaurus, Maiasaura, Saurolophus, Probrachylophosaurus y Prosaurolophus cuentan con crestas óseas de formas tubulares las cuales apuntan hacia atrás, sin embargo, tampoco se sabe con certeza su utilidad.[23][24]
Historia

En 1856, Joseph Leidy, describió los géneros Trachodon y Thespesius a partir de un puñado de dientes y vértebras aisladas respectivamente, siendo estos, unos de los primeros dinosaurios descritos de Norteamérica. En 1858, nuevamente Leidy describió un animal con mayor cantidad de restos y mejor preservados provenientes formaciones del Cretácico de Nueva Jersey, Estados Unidos y lo nombró Hadrosaurus.

En 1869, Edward Drinker Cope describiría dos nuevos géneros de dinosaurios con cierta afinidad a los restos de Hadrosaurus: Hypsibema y Orthomerus. Posteriormente, en 1870, publica una sinopsis de diferentes animales extintos de Norteamérica donde define por vez primera a la familia Hadrosauridae de forma impresa (el documento se había presentado en forma oral en 1868 y 1869), asimismo, postula que las especies Trachodon mirabilis y Thespesius occidentalis deberían reasignarse en especies del género Hadrosaurus.[1]
Más tarde, Richard Lydekker en 1888 invierte la asiganacíon de Cope, posicionando a Hadrosaurus foulkii como Trachodon foulkii y erigiendo la familia Trachodontidae para abarcar a estos animales, dejando como obsoleto el término Hadrosauridae por la sinonimia de su género tipo. Esta clasificación prevaleció por años, donde Kritosaurus, Saurolophus, Hypacrosaurus, Corythosaurus y otros géneros erigidos en la década de 1910, fueron catalogados bajo el nombre de "trachodóntidos".[6]
Fue hasta 1918, cuando Lawrence Lambe concluyé que el género Trachodon se basaba en restos muy pobres y posiblemente eran un quimera de un hadrosáurido y un ceratópsido, regresando la prioridad al clado Hadrosauridae principalmente porque este fue descrito anteriormente, además de que el género Hadrosaurus tenía mayor validez por tener elementos fósiles mucho mejor preservados. A partir de aquí, a los dinosaurios con batería dental compleja compuesta por hileras y con pico similar al de los patos se le empezó a nombrar "hadrosáuridos".[25]
Clasificación
En 1870, Edward Drinker Cope definió inicialmente a esta familia con la siguiente diagnosis muy simple: "dientes en varias hileras formando una pared vertical, posiblemente tenga 3 metatarsos".[1] Paul Sereno emitió una publicación en 1998 de la filogenia general de los dinosaurios, en esta define a Hadrosauridae como: "Saurolophus, Parasaurolophus, su ancestro común y todos sus descendientes.[9]

Taxonomía
- ¿Claosaurus? - (Kansas, EE. UU.)
- Eotrachodon - (Alabama, EE. UU.)
- Lapampasaurus - (La Pampa, Argentina)
- ¿Tanius? - (Shandong, China)
- ¿Telmatosaurus? - (Brașov, Rumania)
- Subfamilia Hadrosaurinae
- Hadrosaurus - (Nueva Jersey, EE. UU.)
- Subfamilia Lambeosaurinae
- Adelolophus - (Utah, EE. UU.)
- Arenysaurus - (Huesca, España)
- ¿Arstanosaurus? - (Kyzylorda, Kazajistán)
- Blasisaurus - (Huesca, España)
- Kazaklambia - (Kyzylorda, Kazajistán)
- Nanningosaurus - (Guangxi, China)
- Tribu Aralosaurini
- Aralosaurus - (Kyzylorda, Kazajistán)
- Canardia - (Alto Garona, Francia)
- Jaxartosaurus - (Kyzylorda, Kazajistán & ¿Sinkiang?, China)
- Tribu Lambeosaurini
- Amurosaurus- (Amur, Rusia)
- ¿Angulomastacator? - (Texas, EE. UU.)
- Corythosaurus - (Alberta, Canadá)
- Hypacrosaurus - (Alberta, Canadá & Montana, EE. UU.)
- Lambeosaurus - (Alberta, Canadá & ¿Montana, EE. UU.?)
- Magnapaulia - (Baja California, México)
- Nipponosaurus - (Sajalín, Rusia)
- Olorotitan - (Amur, Rusia)
- Sahaliyania - (Heilongjiang, China)
- Velafrons - (Coahuila, México)
- Tribu Parasaurolophini
- Charonosaurus - (Heilongjiang, China)
- Parasaurolophus - (Alberta, Canadá; Utah & Nuevo México, EE. UU.)
- Tribu Tsintaosaurini
- Koutalisaurus - (Lérida, España)
- Pararhabdodon - (Lérida, España)
- Tsintaosaurus - (Shandong, China)
- Subfamilia Saurolophinae
- ¿Barsboldia? - (Ömnögovi, Mongolia)
- ¿Lophorhothon? - (Albama & ¿Carolina del Norte?, EE. UU.)
- ¿Willinakaqe? - (Río Negro, Argentina)
- Tribu Brachylophosaurini
- Acristavus - (Montana & Utah, EE. UU.)
- Brachylophosaurus - (Alberta, Canadá & Montana, EE. UU.)
- Maiasaura - (Montana, EE. UU.)
- Probrachylophosaurus - (Montana, EE. UU.)
- ¿Wulagasaurus? - (Heilongjiang, China)
- Tribu Edmontosaurini
- Edmontosaurus - (Alberta, Canadá; Dakota del Sur & Montana, EE. UU.)
- Kerberosaurus - (Amur, Rusia)
- Kundurosaurus - (Amur, Rusia)
- Shantungosaurus - (Shandong, China)
- Ugrunaaluk - (Alaska, EE. UU.)
- Tribu Kritosaurini
- Gryposaurus - (Alberta, Canadá, Montana, ¿Texas? & Utah, EE. UU.)
- Kritosaurus - (¿Coahuila?, ¿Chihuahua?, México; Nuevo México & ¿Texas?, EE. UU.)
- Latirhinus - (Coahuila, México)
- Naashoibitosaurus - (Nuevo México, EE. UU.)
- Rhinorex - (Utah, EE. UU.)
- Secernosaurus - (Chubut, Argentina)
- Tribu Saurolophini
- Augustynolophus - (California, EE. UU.)
- Bonapartesaurus - (Provincia de Río Negro, Argentina)
- Prosaurolophus - (Alberta, Canadá & Montana, EE. UU.)
- Saurolophus - (Alberta, Canadá; Ömnögovi, Mongolia; ¿Heilongjiang?, China)
- Incertae sedis y géneros dudosos
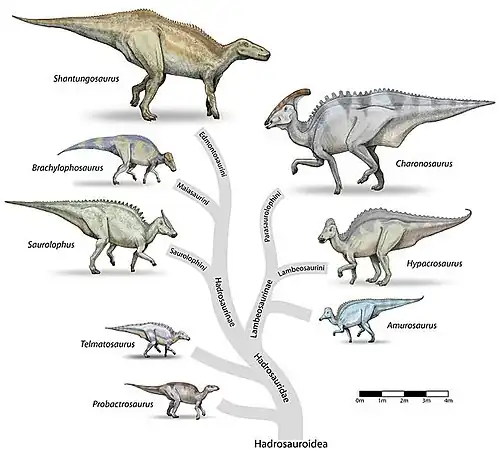
Filogenia
El siguiente cladograma sigue el análisis filogenético de Prieto-Márquez et al. 2012.[19]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Cladograma siguiendo el análisis filogenético de Prieto-Márquez, 2014 (Las relaciones con Lambeosaurinae al igual con los hadrosauroideos basales no son mostradas).[26]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiología

Alimentación
La boca de los hadrosáuridos tenía centenares de dientes colocados en baterías dentales muy complejas. Estos dientes serían reemplazados continuamente por dientes nuevos.[13][14] Los picos de los hadrosáuridos se usaron para cortar los alimentos, ya sea pelando las hojas o cortándolas.[27][28] Asimismo, se cree que los hadrosáuridos tenían mejillas para mantener la comida en la boca.[29]
En 2009 se descubrió que los hadrosáuridos probablemente pastaron en zonas donde abundaban colas de caballo y vegetación cercana al suelo, en lugar de buscar hojas y ramas de mayor crecimiento. Esta conclusión se basó en la uniformidad de los arañazos en los dientes, lo que sugiere que utilizaban la misma serie de movimientos de la mandíbula una y otra vez. Como resultado, el estudio determinó que la dieta del hadrosáuridos estaba hecha principalmente de hojas y en menor cantidad de los objetos más abultados, tales como ramas o tallos, que podrían haber requerido un método de masticación diferente y creado diferentes patrones de desgaste.[30]
Posteriormente en 2013 se examinó la coexistencia de herbívoros en el continente insular de Laramidia, durante el Cretácico tardío. Se concluyó que los hadrosáuridos podían llegar a árboles y arbustos de bajo crecimiento que estaban fuera del alcance de ceratopsidos, anquilosaurios y otros pequeños herbívoros. Los hadrosáuridos eran capaces de alimentarse con un alcance hasta 2 metros cuando estaban en posición cuadrúpeda, y hasta 5 metros en posición bípeda.[31]
Coprolitos colectados de la Formación Two Medicine que datan del Cretácico superior, y que han sido asignados como elementos procedentes de hadrosáuridos, muestran que los animales a veces deliberadamente comían madera podrida. La madera en sí misma no es nutritiva, pero la madera en descomposición habría contenido hongos, material de madera descompuesto y detritos, todos los cuales habrían sido nutritivos.[32]
Por otro lado, coprolitos procedentes de la Formación Kaiparowits presentan fragmentos parcialmente digeridos de crustáceos, concretamente cangrejos de tamaño medio, por lo que estos restos plantean que, al igual que un gran número de aves y mamíferos herbívoros modernos, los hadrosáuridos ingerían pequeños invertebrados con el objetivo de cubrir las deficiencias de proteínas y calcio de sus dietas. Se teoriza que este comportamiento alimenticio, al igual que en las aves, puede haber estado asociado con la anidación, como parece sugerir la acumulación inusualmente grande de coprolitos en el área, con algunos acumulaciones formadas en periodos diferentes (el uso de letrinas comunales es común entre las aves que anidan).[33]
Desarrollo
Las extremidades de los hadrosáuridos juveniles son anatómica y proporcionalmente similares a las de los animales adultos. Sin embargo, las articulaciones suelen mostrar "erosión predeposicional o superficies articulares cóncavas", probablemente debido a la tapa cartilaginosa que cubre los extremos de los huesos. La pelvis de un los especímenes juveniles era similar a la de un individuo mayor.[11]
La evidencia sugiere que los hadrosáuridos jóvenes habrían caminado solamente sobre sus dos patas traseras, mientras que los adultos habrían caminado sobre los cuatro. A medida que el animal envejecía, las extremidades delanteras se volvieron más robustas para asumir peso, mientras que las patas traseras se volvieron menos robustas al pasar a caminar sobre las cuatro patas. Además, los miembros delanteros de los animales eran más cortos que sus miembros traseros.[12]
Ritmo circadiano
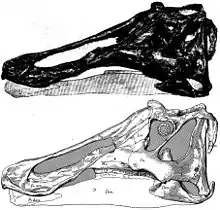
Las comparaciones entre el anillo esclerótico de varios géneros de hadrosáuridos, como Corythosaurus, Prosaurolophus y Saurolophus, con los de las aves modernas y los reptiles sugieren que pudieron tener intervalos de actividad esporádicos y aleatorios durante el día o la noche en para adquirir el alimento, socializar con otros organismos y realizar otras actividades necesarias para el sustento.[34]
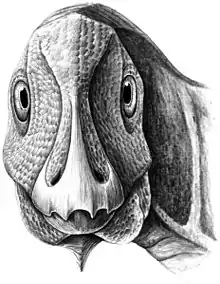
Paleopatología
En 2003 se descubrieron evidencias de tumores, incluyendo hemangioma, fibroma desmoplásico, cáncer metastásico y osteoblastoma en esqueletos fosilizados de Brachylophosaurus.[35]
Restos de un Telmatosaurus juvenil fue examinado en 2016 donde se identificó un gran tumor no canceroso llamado ameloblastoma en su dentario izquierdo. La presencia de este tumor benigno en un dinosaurio no tiene precedentes, pues antes del descubrimiento, los ameloblastomas solo eran conocidos de mamíferos modernos (incluyendo humanos) y reptiles.[36]
También en 2016 se describió una ulna y un radio de un hadrosáurido indeterminado, proveniente de la Formación Navesink en Nueva Jersey, el cual fue severamente afectada por una artritis séptica.[37]
Referencias
- Cope, E. D. (1870). «Synopsis of the extinct Batrachia, Reptilia and Aves of North America». Transactions of the American Philosophical Society, New Series 14 (1): 91.
- Lull, R. S.; Wright, N. E. (1942). Hadrosaurian Dinosaurs of North America. Geological Society of America Special Paper 40. pp. 1-270.
- Marsh, O. C. (1890). «Additional characters of the Ceratopsidae, with notice of new Cretaceous dinosaurs». American Journal of Science 39: 418-426.
- Parks, W. A. (1923). «Corythosaurus intermedius, a new species of trachodont dinosaur». University of Toronto Studies, Geological Series 15: 1-57.
- Brown, B. (1914). «Article XXXV - Corythosaurus casuarius, a new crested dinosaur from the Belly River Cretaceous; with Provisional classification of the family Trachodontidae». Bulletin of the AMNH 33: 559-565.
- Lydekker, R. (1888). «Part 1. Containing the Orders Ornithosauria, Crocodilia, Dinosauria, Squamata, Rhynchocephalia, and Proterosauria». Catalogue of the fossil Reptilia and Amphibia in the British Museum. Londres, Inglaterra: Order of the Trustees. p. 241.
- Lund, E. K.; Gates, T. A. (2006). «A historical and biogeographical examination of hadrosaurian dinosaurs». En Lucas, S.G.; and Sullivan, R. M. (eds.), ed. Late Cretaceous Vertebrates from the Western Interior. New Mexico Museum of Natural History and Science Bulletin, 35. Albuqueque, New Mexico: New Mexico Museum of Natural History and Science. pp. 263-276.
- Norman, D. B.; Witmer, L. M.; Weishampel, D. B. (2004). «14. Basal Ornithischia». En Weishampel, D. B.; Dodson, P.; Osmólska, H., eds. The Dinosauria (2nd edición). Berkeley and Los Angeles, California: University of California Press. pp. 325-334. ISBN 0-520-24209-2.
- Sereno, P. C. (1998). «A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria». Neues Jahrbuch fur Geologie und Palaontologie Abhandlungen 210 (1): 41-83.
- Prieto-Márquez, A. (2010). «Global phylogeny of Hadrosauridae (Dinosauria: Ornithopoda) using parsimony and Bayesian methods». Zoological Journal of the Linnean Society 159 (2): 435-502. doi:10.1111/j.1096-3642.2009.00617.x.
- Dilkes, D. W. (2001). «An ontogenetic perspective on locomotion in the Late Cretaceous dinosaur Maiasaura peeblesorum (Ornithischia: Hadrosauridae)». Canadian Journal of Earth Science 38 (8): 1205-1227. doi:10.1139/e01-016.
- Fiorillo, A. R.; Tykoski, R. S. (2016). «Small hadrosaur manus and pes tracks from the Lower Cantwell Formation (Upper Cretaceous) Denali National Park, Alaska: implications for locomotion in juvenile hadrosaurs». PALAIOS 31 (10): 479-482. doi:10.2110/palo.2016.049.
- LeBlanc, A. R.; Reisz, R. R.; Evans, D. C.; Bailleul, A. M. (2016). «Ontogeny reveals function and evolution of the hadrosaurid dinosaur dental battery». BMC Evolutionary Biology 16 (152): 1-13. doi:10.1186/s12862-016-0721-1.
- Horner, J. R.; Weishampel, D. B. (1990). «Twenty: Hadrosauridae». En Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.), ed. The Dinosauria (1st edición). Berkeley: University of California Press. pp. 438–463. ISBN 0-520-24209-2.
- Lambe, L. M. (1917). «A new genus and species of crestless hadrosaur from the Edmonton Formation of Alberta.». The Ottawa Naturalist 31 (7): pp. 65-73.
- Bell, P. R. (2012). «Standardized terminology and potential taxonomic utility for hadrosaurid skin impressions: a case study for Saurolophus from Canada and Mongolia». En Dodson, Peter, ed. PLoS ONE 7 (2): e31295. doi:10.1371/journal.pone.0031295.
- Glut, D. F. (1997). «Shantungosaurus». Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co. pp. 816-817. ISBN 0-89950-917-7.
- Hu C.; Cheng Z.; Pang Q; Fang X. (2001). Shantungosaurus giganteus (en chino). Pekín: Geological Publishing House. pp. 123–135 [English abstract]. ISBN 7-116-03472-2.
- Prieto-Márquez, A.; Chiappe, L. M.; Joshi, S. H. (2012). «The lambeosaurine dinosaur Magnapaulia laticaudus from the Late Cretaceous of Baja California, Northwestern Mexico». En Dodson, Peter, ed. PLoS ONE 7 (6): e38207. doi:10.1371/journal.pone.0038207.
- Pereda-Suberbiola, X.; Canudo, J. I.; Cruzado-Caballero, P.; Barco, J. L.; López-Martínez, N.; Oms, O.; Ruiz-Omeñaca, J. I. (2009). «The last hadrosaurid dinosaurs of Europe: A new lambeosaurine from the Uppermost Cretaceous of Aren (Huesca, Spain)». Comptes Rendus Palevol 8 (6): 559-572. doi:10.1016/j.crpv.2009.05.002.
- Hopson, J. A. (1975). «The evolution of cranial display structures in hadrosaurian dinosaurs». Paleobiology, Cambridge University Press 1 (1): 21-43.
- Bell, P. R.; Fanti, F.; Currie, P. J.; Arbour, V. M. (2014). «A mummified duck-billed dinosaur with a soft-tissue cock’s comb». Current Biology 24 (1): 70-75. doi:10.1016/j.cub.2013.11.008.
- Freedman Flower, E. A.; Horner, J. R. (2015). «A New Brachylophosaurin Hadrosaur (Dinosauria: Ornithischia) with an Intermediate Nasal Crest from the Campanian Judith River Formation of Northcentral Montana». En Dodson, Peter, ed. PLoS ONE 10 (11): e0141304. doi:10.1371/journal.pone.0141304.
- Prieto-Márquez, A.; Wagner, J. R.; Bell, P. R.; Chiappe, L. M. (2015). «The late-surviving ‘duck-billed’dinosaur Augustynolophus from the upper Maastrichtian of western North America and crest evolution in Saurolophini». Geological Magazine, Cambridge University Press 152 (02): 225-241.
- Lambe, L. M. (1918). «On the genus Trachodon of Leidy». The Ottawa Naturalist 31 (11): 135-139.
- Prieto-Márquez, A. (2014). «Skeletal morphology of Kritosaurus navajovius (Dinosauria: Hadrosauridae) from the Late Cretaceous of the North American south-west, with an evaluation of the phylogenetic systematics and biogeography of Kritosaurini.». Journal of Systematic Palaeontology 12 (2): pp. 133-175. doi:10.1080/14772019.2013.770417.
- Ostrom, J. H. (1964). «A reconsideration of the paleoecology of the hadrosaurian dinosaurs». American Journal of Science 262 (8): 975-997. doi:10.2475/ajs.262.8.975.
- Galton, P. M. (1970). «The posture of hadrosaurian dinosaurs». Journal of Paleontology 44 (3): 464-473.
- Galton, P. M. (1973). «The cheeks of ornithischian dinosaurs». Lethaia 6 (1): 67-89. doi:10.1111/j.1502-3931.1973.tb00873.x.
- Williams, Vincent S.; Barrett, Paul M.; Purnell, Mark A. (2009). «Quantitative analysis of dental microwear in hadrosaurid dinosaurs, and the implications for hypotheses of jaw mechanics and feeding». Proceedings of the National Academy of Sciences 106 (27): 11194-11199. doi:10.1073/pnas.0812631106.
- Mallon, Jordan C.; Evans, David C.; Ryan, Michael J.; Anderson, Jason S. (2013). «Feeding height stratification among the herbivorous dinosaurs from the Dinosaur Park Formation (upper Campanian) of Alberta, Canada». BMC Ecology 13: 14. doi:10.1186/1472-6785-13-14.
- Chin, K. (2007). «The Paleobiological Implications of Herbivorous Dinosaur Coprolites from the Upper Cretaceous Two Medicine Formation of Montana: Why Eat Wood?». PALAIOS 22 (5): 554. doi:10.2110/palo.2006.p06-087r.
- Chin, K.; Feldmann, R. M.; Tashman, J. N. (2017). «Consumption of crustaceans by megaherbivorous dinosaurs: dietary flexibility and dinosaur life history strategies». Scientific reports 7 (11163): 1-11. doi:10.1038/s41598-017-11538-w.
- Schmitz, L.; Motani, R. (2011). «Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology». Science 332 (6030): 705-708. doi:10.1126/science.1200043.
- Rothschild, B. M.; Tanke, D. H.; Helbling II, M.; Martin, L. D. (2003). «Epidemiologic study of tumors in dinosaurs». Naturwissenschaften 90 (11): 495-500. doi:10.1007/s00114-003-0473-9.
- Dumbravă, M. D.; Rothschild, B. M.; Weishampel, D. B.; Csiki-Sava, Z.; Andrei, R. A.; Acheson, K. A.; Codrea, V. A. (2016). «A dinosaurian facial deformity and the first occurrence of ameloblastoma in the fossil record». Scientific Reports 6: 29271. doi:10.1038/srep29271.
- Anné, J.; Hedrick, B. P.; Schein, J. P. (2016). «First diagnosis of septic arthritis in a dinosaur». Royal Society Open Science 3 (8): 160222. doi:10.1098/rsos.160222.
| Control de autoridades |
|
|---|
 Datos: Q271841
Datos: Q271841 Multimedia: Hadrosauridae / Q271841
Multimedia: Hadrosauridae / Q271841 Especies: Hadrosauridae
Especies: Hadrosauridae