Faringotremas
Los faringotremas son órganos de alimentación por filtración que se encuentran en cordados no vertebrados (cefalocordados y urocordados), y hemicordados que viven en ambientes acuáticos. Son segmentos repetidos ubicados en la faringe del animal, teniendo algunas especies de hemicordados hasta 200.[1] Los faringotremas están temporalmente presente en las etapas embriónicas de desarrollo de los tetrápodos. La presencia de hendiduras similares a farigotremas en el cuello de los embriones humanos llevó a Ernst Haeckel a postular la Teoría de la Recapitulación; que, aunque falsa, contenía elementos de verdad en ella, tal como fue explorado por Stephen Jay Gould en Ontogenia y Filogenia.[2] Sin embargo, es aceptado en la actualidad que los faringotremas son homólogos con las bolsas faríngeas y no con las hendiduras del cuello.
Arcos faríngeos en vertebrados
En los vertebrados los arcos faríngeos derivan de las tres capas germinales.[3] Las células de la cresta neural entran en estos arcos donde contribuyen con características craniofaciales, como hueso y cartílago.[3] Sin embargo, la existencia de estructuras faríngeas previas a la evolución de las células de la cresta neural es indicada por la existencia de mecanismos del desarrollo del arco neural independientes de estas.[4] El primero arco faríngeo, más anterior, da origen a la mandíbula oral. El segundo da lugar Hioides.[3] En los peces el resto de arcos posteriores contribuyen al esqueleto branquial, que da soporte a las branquias; en los tetrápodos los arcos anteriores dan lugar a componentes de los oídos, amígdalas y el timo.[5] La base genética del desarrollo de los arcos está bien caracterizada. Se ha comprobado que los genes HOX y otros relacionados con el desarrollo embrionario, como los DLX son importantes para el desarrollo de los patrones anterior/posterior y dorsal/ventral de los arcos faríngeos.[6] De forma interesante, algunos peces tienen sus mandíbulas en su garganta, conocidas como mandíbulas faríngeas, que se desarrollan usando los mismos caminos genéticos involucrados en la formación de las mandíbulas orales.[7]
Evolución
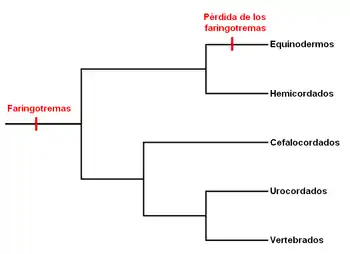
La presencia de faringotremas en hemicordados llevó a debates sobre si éstas eran homólogas a las hendiduras encontradas en los cordados, o eran el resultado de evolución convergente.[8] Con el posicionamiento de los hemicordados y equinodermos como grupos hermanos de los cordados han surgido hipótesis que sugieren que los faringotremasse encontraban presentes sus antepasados deuteróstomos.[9] Extrañamene algunos taxones extintos de equinodermos no poseen estructuras faríngeas, aunque la evidencia fósil revela que los antepasados de os equinodermos poseían estructuras similares a las agallas.[10] En análisis genético y comparativo del desarrollo de estas estructuras faríngeas entre los hemicordados y urocordados han revelado datos importantes sobre la evolución del plan corporal del deuteróstomo.[11] La biología molecular comparativa ha revelado que los genes PAX1/9 (que codifican los factores de trascripción) son expresados en patrones similares entre hemicordados y urocordados; En los vertebrados Pax 1 y Pax 9 son expresados en las bolsas faríngeas y son importantes en el desarrollo del timo.[12] De manera interesante, la aplicación de un exceso de ácido retinoico en vertebrados resulta en anormalidades faríngeas, y en el cefalocordado Branchiostoma lanceolatum resulta en la ausencia de faringotremas, sugiriendo que el ácido retinoico actúa con el mismo mecanismoen ambos.[13] Estos estudios indican que los faringotremas encontrados en hemicordados y cordados son homólogos desde un punto de vista molecular.
Referencias
- Gerhart J, Lowe C, Kirschner M (2005). «Hemichordates and the origin of chordates». Current Opinions in Genetics & Development 15 (4): 461-467. PMID 15964754. doi:10.1016/j.gde.2005.06.004.
- Gould, S.J. (1977). Ontogeny and Phylogeny. Cambridge, Massachusetts: The Belknap Press of Harvard University Press. pp. vii-viii. ISBN 0-674-63940-5.. Also ISBN 0-674-63941-3 (paperback)
- Graham A (2003). «Development of the pharyngeal arches». Am J Med Genet A 199 (3): 251-256. PMID 12784288. doi:10.1002/ajmg.a.10980.
- Graham A, Smith A (2001). «Patterning the pharyngeal arches». Bioessays 23 (1): 54-61. PMID 11135309. doi:10.1002/1521-1878(200101)23:1<54::AID-BIES1007>3.0.CO;2-5.
- Kardong KV (2003). «Vertebrates: Comparative Anatomy, Function, Evolution». Third edition. New York (McGraw Hill).
- Depew MJ, Lufkin T, Rubenstein JLR (2002). «Specification of jaw subdivisions by Dlx genes». Science 298 (5592): 381-385. PMID 12193642. doi:10.1126/science.1075703.
- Fraser GJ, Hulsey D, Bloomquist RF, Uyesugi K, Manley NR, Streelman T (2009). «An ancient Gene Network is co-opted for teeth on old and new jaws». Plos Biology 7 (2): 0233-0247. PMC 2637924. PMID 19215146. doi:10.1371/journal.pbio.1000031.
- Gee H (1996). Before the backbone. Views on the origin of vertebrates. London (Chapman & hall).
- Winchell CJ, Sullivan J, Cameron CB, Swalla BJ, Mallatt J (2002). «Evaluating hypotheses of deuterostome phylogeny and chordate evolution with new LSU and SSU ribosomal DNA data». Mol Biol Evol 19 (5): 762-776. PMID 11961109.
- Shu D, Morris SC, Zhang ZF, Liu JN, Han J, Chen L, Zhang XL, Yasui K, Li Y (2003). «A new species of yunnanozoan with implications for deuterostome evolution». Science 299 (5611): 1380-1384. PMID 12610301. doi:10.1126/science.1079846.
- Brown FD, Prendergast A, Swalla BJ (2008). «Man is but a worm: chordate origins». Genesis 46 (11): 605-613. PMID 19003926. doi:10.1002/dvg.20471.
- Ogasawara M, Wada H, Peters H, Satoh N (1999). «Developmental expression of Pax 1/9 genes in urochordate and hemichordate gills: insight into function and evolution of the pharyngeal epithelium». Development 126 (11): 2539-2550. PMID 10226012.
- Holland LZ, Holland N (1996). «Expression of AmphiHox-1 and AmphiPax-1 in amphioxus embryos treated with retinoic acid: insights into evolution and patterning of the chordate nerve cord and pharynx». Development 122 (6): 1829-1838. PMID 8674422.
| Control de autoridades |
|
|---|
 Datos: Q3378614
Datos: Q3378614