Galactógeno
El galactógeno es un polisacárido de galactosa que sirve para el almacenamiento de energía en los caracoles pulmonados y algunos Caenogastropoda.[1] Este polisacárido es exclusivo de la reproducción, y solo se encuentra en la glándula del albumen del sistema reproductor femenino de estos caracoles y en el fluido perivitelino de los huevos.
| Galactógeno | ||
|---|---|---|
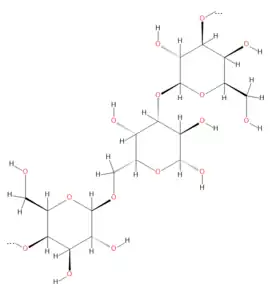 Estructura general del galactógeno. | ||
| General | ||
| Fórmula molecular | C24H42O21 | |
| Familia | Polisacárido | |
El galactógeno sirve como reserva de energía tanto para los embriones en desarrollo como para los juveniles recién eclosionados, y es luego reemplazado por el glucógeno en juveniles de estadios avanzados y en adultos.[2] Aún no se ha identificado la ventaja de acumular galactógeno en vez de glucógeno en los huevos de caracoles, pero se han propuesto algunas hipótesis al respecto (ve abajo).[3]
Ocurrencia y distribución
El galactógeno ha sido encontrado en la glándula del albumen de caracoles pulmonados como Helix pomatia, Limnaea stagnalis, Oxychilus cellarius, Lissachatina fulica, Aplexa nitens,Otala lactea, Bulimnaea megasoma, Ariolimax columbianis, Ariophanta, Biomphalaria glabrata, y Strophochelius oblongus.[4][5][6][7][8][9][10][11][12][13] Este polisacárido fue también identificado en los Caenogastropoda Pila virens y Viviparus, Pomacea canaliculata, y Pomacea maculata.[14][15]
En los gasterópodos adultos, el galactógeno está limitado a la glándula del albumen, mostrando una gran variación en contenido durante el año, alcanzando su pico máximo en la época reproductiva.[2] Durante la estación reproductiva, este polisacárido es rápidamente restaurado en la glándula del albumen después de ser transferido a los huevos, disminuyendo su cantidad total solo después de varias ovoposiciones repetidas.[16][17] En la especie Pomacea canaliculata, el galactégeno actuaría, junto con las perivitelinas, como factor limitante principal de la reproducción. Este polisacárido ha sido identificado en la zona del aparato de Golgi en las células secretoras de la glándula del albumen en la forma de gránulos discretos de 200 Å de diámetro.[18][19][20] El aspecto de los gránulos de galactógeno dentro de los glóbulos secretores sugiere que éste es el sitio de síntesis del polisacárido.[1]
Además de la glándula del albumen, el galactógeno es también encontrado como un componente importante del fluido perivitelino de los huevos de caracol, comprendiendo la fuente de energía principal para el embrión en desarrollo.[4][5][14][15]
Estructura
El galactógeno es un polímero de galactosa con variaciones estructurales especie-específicas. En este polisacárido, la D-galactosa esta predominantemente unida por enlaces glicosídicos β (1→3) y β (1→6); sin embargo, en algunas especie también puede esta unida por enlaces β (1→2) y β (1→4).[3] El galactógeno de los caracoles acuáticos Basommatophora (por ejemplo Lymnaea, Biomphalaria) esta altamente ramificado con solo 5-8 % de los residuos de azúcar en secciones lineales, y los enlaces β(1→3) y β(1→6) se alternan más o menos regularmente. En los caracoles terrestres Stylommatophora (como por ejemplo Helix, Arianta, Cepaea, Achatina), hasta un 20% de los residuos de azúcar están unidos por enlaces β(1→3) de forma lineal. El galactógeno de la especie Ampullarius sp tiene una proporción inusualmente grande de residuos de azúcar en disposición lineal, con 5% β(1→3), 26% β(1→6), y 10% β(1→2). Otros análisis en Helix pomatia sugieren una estructura dicotómica, donde cada una de las unidades de galactopiranose lleva un residuo lateral.[21][22]
El peso molecular del galactógeno extraído de los huevos de Helix pomatia y Limnaea stagnalis fue estimado en 4x106 y 2.2x106, respectivamente.[23][24] En estos caracoles, el galactógeno contiene solo D-galactosa.[25] Según el origen del galactógeno, otros componentes como L-galactosa, L-fucosa, D-glucosa, L-glucosa y residuos de fosfato también pueden estar presentes en la estructura de este polisacárido; por ejemplo, el galactógeno de Ampullarius sp. contiene un 98% de D-galacotosa y un 2% de L- fucosa, mientras que el galactógeno aislado de los huevos de Pomacea maculata consisten en un 68% de D-galactosa y un 32% de D-glucosa.[3][26][15] Residuos de galactosa sustituidos con fosfato han sido encontrados en el galactógeno de algunas especies dentro de varios géneros de caracoles como Biomphalaria, Helix y Cepaea.[27] Por lo tanto, el conocimiento actual indica que el galactógeno podría ser considerado como un homopolisacarido o un heteropolisacarido dominado por galactosa.
Metabolismo
El galactógeno es sintetizado por las células secretoras de la glándula del albumen de los caracoles hembra y más tarde transferidos a los huevos. Este proceso esta bajo control neurohormonal, notablemente por la hormona cerebral galactogenina.[9][28][29] Las rutas metabólicas para la síntesis del glucógeno y el galactógeno están estrechamente relacionadas. Ambas utilizan glucosa como precursor común y su conversión a residuos activados de galactosa está catalizada por la UDP-glucosa 4-epimerasa y la galactose-1-P uridil-transferasa. Estas reacciones permite a la glucosa ser el precursor común tanto para la glucogenogénesis como para la galactogenogénesis.[30] De hecho, ambos polisacáridos se encuentra en las mismas células secretoras de la glándula del albumen y están sujetos a variaciones estacionales independientes.[19] El glucógeno se acumula en otoño como reserva de energía general para la hibernación, mientras que el galactógeno es sintetizado durante la primavera en preparación para la producción de los huevos.[31] Esta generalmente aceptado que la producción del galactógeno está restringida a la nutrición del embrión y, por tanto, es principalmente transferido a los huevos.
Se conoce poco sobre las enzimas sintetizadoras del galactógeno. Una D-galactosiltransferasa fue descripta en la glándula del albumen de Helix pomatia.[32] Esta enzima cataliza la transferencia de una D-galactose a otra con unión (1→6) y es dependiente de la presencia del aceptor galactógeno. De modo parecido, la actividad β-(1→3)-galactosiltransferasa ha sido detectada en extractos de la glándula del albumen de Limnaea stagnalis.[33]
En embriones y juveniles recién eclosionados del huevo, el galactógeno es, probablemente, la fuente de intermediarios metabólicos más importante (vía galactosa). En los caracoles que se alimentan por su cuenta, la dieta principal consiste en los polisacáridos almidón y celulosa, constituido por glucosa. Estos polímeros son digeridos y contribuyen con la glucosa para las rutas del metabolismo intermediario.[1] El consumo del galactógeno comienza en la etapa de gástrula y continúa durante todo el desarrollo. Hasta un 46-78 % del galactógeno del huevo desaparece durante el desarrollo del embrión. El resto es utilizado dentro de los primeros días después de la eclosión.[9]
Solo los embriones y los caracoles recién eclosionados son capaces de degradar el galactógeno, mientras que otros animales, e incluso los caracoles adultos, no cuentan con dicha capacidad. La β-galactosidasa puede ser importante en la liberación de residuos de galactosa desde el galactógeno; sin embargo, aún se desconoce la mayoría de las rutas catabólicas de este polisacárido.[9][34][35][1]
Otras funciones
Además de ser una fuente de energía, otras funciones han sido descriptas para el galactógeno de los huevos de los caracoles, y todas ellas están relacionadas con la defensa y protección del embrión. Dado que los carbohidratos retienen agua, la cantidad alta de este polisacárido protegería a los huevos contra la desecación de aquellos caracoles que tiene oviposición aérea.[36][37] Además, la alta viscosidad que este polisacárido puede conferir al fluido perivitelino ha sido sugerido como una potencial defenza antimicrobiana.
Dado que el galactógeno es un polisacáridos con enlaces β, como la celulosa o la hemicelulosa, se necesitan adaptaciones bioquímicas especiales para explotar éste polisacárido como nutriente, tales como la presencia de enzimas específicas. Sin embargo, aparte de los embriones de caracol y los caracoles recién eclosionados, ningún otro animal parece ser capaz de catabolizar el galactógeno, incluyendo los caracoles adultos. Este hecho llevó a considerar al galactógeno como parte de un sistema de defensa antidepredación exclusivo de los gasterópodos, disuadiendo a los depredadores por medio de bajar el valor nutritivo de huevos.[15]
Referencias
- «Carbohydrates and carbohydrate metabolism in Mollusca.». Chemical Zoology. VII Mollusca. New York: Academic Press. 1972. pp. 219-244.
- «Beitrag zur Kenntnis des Glykogen und Galaktogengehaltes bei Helix pomatia.». Z. Biol. 92: 319-324. 1932.
- Urich, Klaus (1994). Comparative Animal Biochemistry. Springer Berlin Heidelberg. pp. 1-8. ISBN 978-3-642-08181-1. doi:10.1007/978-3-662-06303-3_1.
- «Ober den Galactogengehalt der Eier von Heilix pomatia.». Z. Biol. 92: 325-330. 1932.
- «Studies on the galactogen metabolism in the snail (Helix pomatia L.). 3. Catabolism of galactogen in young animals.». Z. Biol. 115 (2): 133-155. 1965. PMID 5847702.
- Rigby, Joyce E. (20 de agosto de 2009). «Alimentary and Reproductive Systems of Oxychilus Cellarius (Müller) (Stylommatophora)». Proceedings of the Zoological Society of London (en inglés) 141 (2): 311-359. doi:10.1111/j.1469-7998.1963.tb01615.x.
- Ghose, Krishna Chandra (1963). «Reproductive System of the Snail Achatina Fulica». Proceedings of the Zoological Society of London (en inglés) 140 (4): 681-695. ISSN 1469-7998. doi:10.1111/j.1469-7998.1963.tb01993.x.
- «Observations on the polysaccharides of aquatic snails». Journal of Cellular and Comparative Physiology 50 (2): 219-40. October 1957. PMID 13513668. doi:10.1002/jcp.1030500206.
- «Galactogen catabolism by embryos of the freshwater snails, Bulimnaea megasoma and Lymnaea stagnalis». Comparative Biochemistry and Physiology. B, Comparative Biochemistry 53 (4): 439-42. January 1976. PMID 4280. doi:10.1016/0305-0491(76)90194-2.
- «Regulation of galactogen synthesis in the slug Ariolimax columbianus». Comparative Biochemistry and Physiology 29 (2): 841-845. May 1969. ISSN 0010-406X. doi:10.1016/0010-406x(69)91636-3.
- «Galactogen in some common south Indian gastropods with special reference to Pila.». Current Science 23 (9): 301-2. September 1954.
- «Structure of a galactan found in the albumen gland of Biomphalaria glabrata». Carbohydrate Research 3 (4): 445-452. February 1967. ISSN 0008-6215. doi:10.1016/s0008-6215(00)81676-6.
- «Some structural studies on the galactan from the albumen glands of the snail, Strophocheilus oblongus». Carbohydrate Research 16 (2): 327-335. February 1971. ISSN 0008-6215. doi:10.1016/s0008-6215(00)81168-4.
- «Biochemical composition and energy sources during embryo development and in early juveniles of the snail Pomacea canaliculata (Mollusca: Gastropoda)». Journal of Experimental Zoology 280 (6): 375-383. 1998. ISSN 1097-010X. doi:10.1002/(SICI)1097-010X(19980415)280:6<375::AID-JEZ1>3.0.CO;2-K.
- «The eggs of the apple snail Pomacea maculata are defended by indigestible polysaccharides and toxic proteins». Canadian Journal of Zoology 94 (11): 777-785. 12 de septiembre de 2016. ISSN 0008-4301. doi:10.1139/cjz-2016-0049.
- «Biochemical Composition of the Eggs of the Freshwater Snail Lymnaea stagnalis and Oviposition-induced Restoration of Albumen Gland Secretion». International Journal of Invertebrate Reproduction and Development 12 (2): 199-212. 1 de septiembre de 1987. ISSN 0168-8170. doi:10.1080/01688170.1987.10510317.
- «Biosynthesis in the Albumen Gland-Capsule Gland Complex Limits Reproductive Effort in the Invasive Apple Snail Pomacea canaliculata». The Biological Bulletin 235 (1): 1-11. August 2018. PMID 30160995. doi:10.1086/699200.
- «Histochemical observations on galactogen». Stain Technology 27 (2): 81-5. March 1952. PMID 14931082. doi:10.3109/10520295209105064.
- «Ultrastructure of galactogen in the albumen gland of Helix pomatia». Journal of Ultrastructure Research 29 (1–2): 119-40. October 1969. PMID 4310741. doi:10.1016/s0022-5320(69)80059-6.
- «Pallial oviduct of Pomacea canaliculata (Gastropoda): ultrastructural studies of the parenchymal cellular types involved in the metabolism of perivitellins». Cell and Tissue Research 324 (3): 523-33. June 2006. PMID 16453107. doi:10.1007/s00441-005-0132-x.
- «The application of the Barry degradation to snail galactogen.». Proceedings of the Royal Irish Academy. Section B: Biological, Geological, and Chemical Science (Royal Irish Academy) 55: 165-170. January 1952.
- «278. A preliminary investigation of galactogen from the albumin glands of Helix pomatia». Journal of the Chemical Society (Resumed): 1461-1465. 1 de enero de 1938. ISSN 0368-1769. doi:10.1039/JR9380001461. Consultado el 17 de agosto de 2020.
- «[Studies on Galactogen Metabolism of Snails (Helix Pomatia L.) I. The Preparation and Properties of Native Galactogen From Eggs]». Biochemische Zeitschrift 340: 548-51. November 1964. PMID 14331584.
- «Über das native Galaktogen aus den Eiern der Schlammschnecke Lymnaea stagnalis». Biological Chemistry (en inglés) 348 (Jahresband): 1301-1306. 1 de enero de 1967. ISSN 1437-4315. doi:10.1515/bchm2.1967.348.1.1301. Consultado el 17 de agosto de 2020.
- «[Research on metabolism in air-breathing snails. III. Galactogen in the eggs of Lymnaea stagnalis L]». Hoppe-Seyler's Zeitschrift Fur Physiologische Chemie 325 (Jahresband): 251-9. September 1961. PMID 14449055. doi:10.1515/bchm2.1961.325.1.251. Consultado el 17 de agosto de 2020.
- «Some structural studies on the fucogalactan from egg masses of the snail ampullarius sp». Carbohydrate Research 44 (2): 241-249. 1 de noviembre de 1975. ISSN 0008-6215. doi:10.1016/S0008-6215(00)84167-1.
- «Structural Studies on the Galactan from the Albumin Gland of Achatina fulica». Zeitschrift für Naturforschung C 39 (11–12): 1063-1065. 1 de diciembre de 1984. ISSN 1865-7125. doi:10.1515/znc-1984-11-1211.
- «Calcium-dependent release of a neurochemical messenger from the brain of the land snail, Helix pomatia». Brain Research 151 (2): 418-23. August 1978. PMID 28169. doi:10.1016/0006-8993(78)90900-9.
- «Stimulation of Helix pomatia albumen gland galactogen synthesis by putative neurohormone (galactogenin) and by cyclic AMP analogues». Comparative Biochemistry and Physiology Part B: Comparative Biochemistry 71 (3): 417-422. January 1982. ISSN 0305-0491. doi:10.1016/0305-0491(82)90403-5.
- Metabolic Biochemistry and Molecular Biomechanics. Elsevier. 1983. pp. 177-242. ISBN 978-0-12-751401-7. doi:10.1016/b978-0-12-751401-7.50012-0.
- May, Friedrich Julius (1934). Chemische und biologische Untersuchungen über Galaktogen. Lehmann. OCLC 256810367.
- «Biosynthesis of galactogen: identification of a beta-(1----6)-D-galactosyltransferase in Helix pomatia albumen glands». Biochimica et Biophysica Acta 992 (3): 289-97. September 1989. PMID 2505854. doi:10.1016/0304-4165(89)90087-1.
- «Identification of a UDP-Gal:beta-galactoside beta 1----3-galactosyltransferase in the albumen gland of the snail Lymnaea stagnalis». FEBS Letters 221 (1): 139-44. August 1987. PMID 3113997. doi:10.1016/0014-5793(87)80368-x.
- «[In vitro galactogen decomposition by enzymes; studies on Helix pomatia. I. Orientation on occurrence and effect of the galactogen-splitting enzyme]». Biochemische Zeitschrift 324 (1): 19-31. 1953. PMID 13093716.
- «A Survey of the Enzymes from the Gastro-Intestinal Tract of Helix Pomatia». Journal of Experimental Biology (en inglés) 35 (3): 639-648. 1 de septiembre de 1958. ISSN 0022-0949. Consultado el 17 de agosto de 2020.
- «Characterization of the major egg glycolipoproteins from the perivitellin fluid of the apple snail Pomacea canaliculata». Molecular Reproduction and Development 68 (3): 359-64. July 2004. PMID 15112330. doi:10.1002/mrd.20078.
- «Egg carotenoproteins in neotropical Ampullariidae (Gastropoda: Arquitaenioglossa)». Comparative Biochemistry and Physiology. Toxicology & Pharmacology. Fourth Special Issue of CBP dedicated to The Face of Latin American Comparative Biochemistry and Physiology organized by Marcelo Hermes-Lima (Brazil) and co-edited by Carlos Navas (Brazil), Rene Beleboni (Brazil), Rodrigo Stabeli (Brazil), Tania Zenteno-Savín (Mexico) and the Editors of CBP - This issue is dedicated to the memory of two exceptional men, Peter L. Lutz, one of the pioneers of comparative and integrative physiology, and Cicero Lima, journalist, science lover and Hermes-Lima's dad 146 (1–2): 158-67. 1 de julio de 2007. PMID 17320485. doi:10.1016/j.cbpc.2006.10.013.
| Control de autoridades |
|
|---|
 Datos: Q82191590
Datos: Q82191590