Hypsilophodon foxii
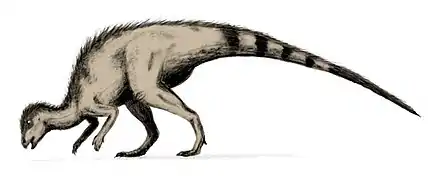
Hypsilophodon foxii es la única especie conocida del género extinto Hypsilophodon ("diente protuberante") de dinosaurio ornitópodo hipsilofodóntido, que vivió a finales del período Cretácico inferior, hace aproximadamente 124 millones de años, en el Barremiense, en lo que hoy es Europa. Los primeros restos de Hypsilophodon fueron encontrados en 1849. La especie tipo y única aceptada, Hypsilophodon foxii, fue nombrada en 1869. Se hicieron abundantes descubrimientos de fósiles en la Isla de Wight, dando una buena impresión de la estructura de la especie. Era un pequeño animal bípedo con una dieta herbívora o posiblemente omnívora. Hypsilophodon alcanzó hasta 1,8 metros de largo, pesaba unos 20 kilogramos y era un corredor ágil. Tenía una cabeza puntiaguda equipada con un pico afilado que se usaba para morder material vegetal, como los loros modernos. Los estudios más antiguos han dado lugar a una serie de ideas erróneas sobre el Hypsilophodon, que era capaz trepar a los árboles, que estaba blindado, que alcanzaba una longitud de 2,3 metros y su presencia fuera de Wight. Durante las últimas décadas, nuevas investigaciones han demostrado gradualmente que todo esto es incorrecto.
| Hypsilophodon foxii | ||
|---|---|---|
| Rango temporal: 124 Ma Cretácico inferior | ||
 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Ornithischia | |
| Suborden: | Neornithischia | |
| Infraorden: | Ornithopoda | |
| Familia: | Hypsilophodontidae | |
| Género: |
Hypsilophodon Huxley, 1869 | |
| Especie: |
H. foxii Huxley, 1869. | |
| Sinonimia | ||
| ||
Descripción
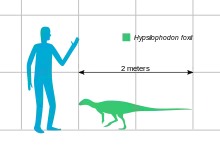
Hypsilophodon era un dinosaurio relativamente pequeño, aunque no tan pequeño como, por ejemplo, Compsognathus. Para el Hypsilophodon a menudo se indica una longitud máxima de 2,3 metros. Esto tiene su origen en un estudio de 1974 realizado por Galton, en el que extrapola una longitud de 2,28 metros en base al espécimen BMNH R 167, un hueso del muslo.[1] Sin embargo, en 2009, Galton concluyó que, de hecho, este fémur pertenecía a Valdosaurus y reducía el tamaño de Hypsilophodon a una longitud máxima conocida de 1,8 metros,[2] siendo la muestra más grande NHM R5829 con una longitud de fémur de 202 milímetros.[1] Los especímenes típicos miden alrededor de 1,5 metros de largo. Hypsilophodon habría alcanzado hasta medio metro de altura. En 2010, Gregory S. Paul estimó un peso de 20 kilogramos para un animal de dos metros de longitud.[3]
Como la mayoría de los dinosaurios pequeños, Hypsilophodon era bípedo, corría en dos piernas. Todo su cuerpo fue construido para correr. Un esqueleto liviano, bajo, con postura aerodinámica, patas largas y cola rígida, inmovilizada por tendones osificados, para mantener el equilibrio. Todo le habría permitido correr notablemente rápido por su tamaño. En 1974, Galton concluyó que habría estado entre los ornitisquio mejor adaptados para correr.[1]
A pesar de vivir en el último de los periodos en que los dinosaurios no avianos caminaban por la tierra, el Cretácico, el Hypsilophodon tenía una serie de características aparentemente "primitivas". Por ejemplo, había cinco dedos en cada mano y cuatro en cada pie. En Hypsilophodon, el quinto dedo había adquirido una función especializada, ser oponible y podía servir para agarrar alimentos.[1] Además, a pesar de que tenía un pico como la mayoría de los ornitisquios, Hypsilophodon todavía tenía cinco dientes triangulares puntiagudos en el frente de la mandíbula superior, el premaxilar. La mayoría de los dinosaurios herbívoros del Cretácico temprano tenían especializaciones en las que los dientes frontales se perdieron por completo, aunque existe cierto debate sobre si estos dientes pueden haber tenido una función especializada en el Hypsilophodon. Más hacia la parte posterior, la mandíbula superior llevaba hasta once dientes en el maxilar superior. La mandíbula inferior tenía hasta 16 dientes. El número fue variable, dependiendo del tamaño del animal.[2] Los dientes de la espalda tenían forma de abanico.[1]

El cráneo de Hypsilophodon era corto y relativamente grande. El hocico era triangular y puntiagudo, terminando en un pico superior cuyo borde de corte era notablemente más bajo que la fila de dientes superiores. La cuenca del ojo era muy grande. Un palpebral con una longitud igual a la mitad del diámetro de la cuenca del ojo ensombreció su sección superior. Un anillo esclerótico de quince pequeñas placas óseas apoyaba la superficie externa del ojo. La parte posterior del cráneo era bastante alta, con una yugal y cuadradoyugal muy grande y alta que cerraba una pequeña fenestra infratemporal altamente posicionada.[1]
La columna vertebral consistía en 9 vértebras cervicales, 15 o 16 vértebras dorsales, 6 a 5 vértebras sacras y aproximadamente 48 vértebras caudales. Gran parte de la espalda y la cola estaban rígidas por largos tendones osificados que conectaban las espinas en la parte superior de las vértebras. Los procesos en la parte inferior de las vértebras de la cola, los cheurones, también estaban conectados por tendones osificados, que, sin embargo, tenían una forma diferente, eran más cortos y estaban rotos y desgastados en un extremo, con la punta del otro extremo afilado dentro del extremo divergente del tendón posterior. Además, hubo varias filas contradireccionales de estas, lo que resultó en un patrón de espina de pescado que completamente inmovilizó el extremo de la cola.[1]
Un concepto erróneo de larga data sobre la anatomía de Hypsilophodon ha sido que estaba blindado. Esto fue sugerido por primera vez por Hulke en 1874, después del hallazgo de una placa ósea en la región del cuello.[4] Si es así, Hypsilophodon habría sido el único ornitópodo blindado conocido.[1] Como señaló Galton en 2008, la supuesta armadura parece ser del torso, un ejemplo de placas intercostales internas asociadas con la caja torácica. Consiste en placas circulares mineralizadas finas que crecen desde el extremo posterior del eje de la costilla central y se superponen al borde frontal de la costilla posterior. Tales placas son más conocidas de Talenkauen y Thescelosaurus , y probablemente fueron de origen cartilaginoso.[5]
Descubrimiento e investigación
Los primeros restos de Hypsilophodon se recuperaron en los primeros días de la paleontología en 1849, cuando los trabajadores de la Isla de Wight desenterraron el llamado bloque Mantell-Bowerbank. Una pieza se vendió a Gideon Mantell, la otra al naturalista James Scott Bowerbank. Sin embargo, en ese momento, se pensaba que los huesos pertenecían a un Iguanodon joven, primero Mantell en 1849,[6] y luego Richard Owen en 1855, describiendo el bloque como tal.[7]

No fue hasta 1870 que el paleontólogo Thomas Henry Huxley pudo publicar una descripción más completa de Hypsilophodon como la conocemos hoy, entendiendo que representaba una especie diferente de Iguanodon. En 1868 tuvo acceso a un espécimen encontrado en enero de ese año por el Reverendo William Fox,[8] que también había descubierto algunos fósiles del animal.[9] Este espécimen incluía el cráneo, que había faltado con el bloque Mantell-Bowerbank. Huxley anunció por primera vez la nueva especie en 1869 en una conferencia. El texto de este, publicado el mismo año, forma el artículo oficial de nombramiento, porque contenía una descripción suficiente.[9] En 1870, Huxley expandió esto en un artículo completo, en el que se convirtió en el primer investigador en informar que los Ornithischia, que aún no se conocía como tal en ese momento, tenía un hueso púbico que apuntaba hacia atrás como pájaros.[10]
La especie tipo fue nombrada Hypsilophodon foxii.[9] Existe una incomprensión persistente en cuanto al significado del nombre genérico, que a menudo se traduce directamente del griego como "diente alto".[2] En realidad, Huxley, de forma análoga a la forma en que se formó el nombre del género relacionado Iguanodon, "diente de iguana", tenía la intención de nombrar al animal como un lagarto herbívoro existente, eligiendo para este papel el Hypsilophus y combinando su nombre con Griego ὀδών, odon , "diente".[11] Hypsilophodon significa "diente de Hypsilophus". El griego ὑψίλοφος, Hipsilofos, significa "de cresta alta" y se refiere al volante trasero del lagarto, no a los dientes de Hypsilophodon en sí, que en ningún caso son muy altos.[2] El nombre específico de H. foxii honra a Fox.[9]
El holotipo del animal, que hoy tiene el número de inventario NHM R197, es el cráneo encontrado por Fox. El bloque Mantell-Bowerbank es el paratipo. Sus dos piezas se han combinado en la colección del Museo de Historia Natural como especímenes NHM 28707, 39560-1.[2] Owen no estaba convencido por la conclusión de Huxley de que Hypsilophodon representaba un género diferente y en 1874 cambió el nombre de la especie Iguanodon foxii.[12] Sin embargo, esto fue nuevamente rechazado por John Whitaker Hulke, quien había obtenido algunos especímenes adicionales de Fox,[13] en 1874 describió algunas presuntas armaduras dérmicas,[4] y en 1882 publicó una osteología completa de la especie.[4] Más tarde, el número de especímenes se incrementó por Reginald Walter Hooley . En 1905, el barón Franz Nopcsa dedicó un estudio a Hypsilophodon,[14] y en 1936 William Elgin Swinton hizo lo mismo,[15] con motivo del montaje de dos esqueletos restaurados en el Museo Británico de Historia Natural.[16]

La investigación moderna de Hypsilophodon comenzó con los estudios de Peter Malcolm Galton, comenzando con su tesis de 1967.[17] Aunque se siguen haciendo nuevos hallazgos,[2] la mayoría de los especímenes conocidos de Hypsilophodon se descubrieron entre 1849 y 1921 y están en posesión del Museo de Historia Natural que adquirió las colecciones de Mantell, Fox, Hulke y Hooley.[1] Estos representan unos veinte animales individuales. Aparte del holotipo y el paratipo, los ejemplares más significativos son, NHM R5829, el esqueleto de un animal grande, NHM R5830 y NHM R196/196a, ambos esqueletos de animales juveniles y NHM R2477, un bloque con un cráneo junto con dos columnas vertebrales separadas. Los fósiles de otros lugares, especialmente de la parte continental de Britania, Portugal y España, se han referido una vez a Hypsilophodon. Sin embargo, en 2009, Galton llegó a la conclusión de que los especímenes de Gran Bretaña propiamente dichos eran indeterminables o pertenecían al Valdosaurus y que los fósiles del resto de Europa eran de especies relacionadas pero diferentes. Esto deja los hallazgos en la Isla de Wight , en la costa sur de Inglaterra, como los únicos fósiles de Hypsilophodon auténticos conocidos.[2] Los fósiles se han encontrado en el lecho de Hypsilophodon, una capa de marga de un metro de grosor que emerge en una franja de 1200 metros de largo a lo largo de Cowleaze Chine paralela a la costa suroeste de Wight,[1] parte de la Formación Wessex superior y data del tardío Barremian, de unos 126 millones de años. Informes de que Hypsilophodon estaría presente en la última Formación Vectis , Galton en 2009 considerada como no demostrada.[2]
Solo hay una especie conocida de Hypsilophodon, la H. foxii original de Huxley. Galton y James Jensen nombraron otra especie en 1978/1979, Hypsilophodon wielandi, que se basa en un hueso del muslo de Dakota del Sur, Estados Unidos, ejemplar AMNH 2585 encontrado en 1891 por George Reber Wieland.[18] Esta especie fue vista como un indicativo de un puente terrestre tardío entre América del Norte y Europa,[19] pero ahora se considera un ornitópodo basal indeterminado[20] o un ornitisquio no ornitópodo.[21] En 2009, Galton lo consideró un nomen dubium , el fémur en vista de la procedencia que posiblemente pertenezca a Zephyrosaurus.[2]
Clasificación
Huxley originalmente asignó Hypsilophodon a los Iguanodontidae.[10] En 1882, Louis Dollo erigío un Hypsilophodontidae separado para incluirlo.[22] A mediados del siglo XX, que se había convertido en la clasificación aceptada, pero a principios del siglo XXI se hizo evidente a través del uso del análisis cladístico que los hipsilopodóntidos formaban un grupo parafiletico de brotes sucesivos de todo Neornithischia.[20] Hypsilophodon en la visión moderna, por lo tanto, simplemente es un ornitópodo basal. En 2017, Daniel Madzia, Clint Boyd y Martin Mazuch eliminaron Hypsilophodon de Ornithopoda en conjunto, ubicándolo como el grupo hermano de Cerapoda , en una posición más basal, varios otros "hipsilopodóntidos" han sufrido reclasificaciones similares.[21]
Filogenia
El siguiente cladograma se reproduce a partir del estudio de Madzia, Boyd y Mazuch de 2017[21]
| Clypeodonta |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiología
Debido a su pequeño tamaño, el Hypsilophodon se alimenta de vegetación de baja, dado que el hocico puntiagudo probablemente prefiere material vegetal de alta calidad, como brotes jóvenes y raíces, a la manera de los ciervos modernos. La estructura de su cráneo, con los dientes colocados muy atrás en la mandíbula, sugiere fuertemente que tenía mejillas, una característica avanzada que habría facilitado la masticación de alimentos. Había 23 a 27 dientes maxilares y dentarios con crestas verticales en las mandíbulas superior e inferior del animal que, debido al hecho de que la hilera de dientes de la mandíbula inferior, con sus dientes curvados hacia fuera, encajaban dentro de la mandíbula superior , con sus dientes curvados hacia adentro, parecen haberse autoafilado, la oclusión desgastaba los dientes y proporcionaba un simple mecanismo de masticación. Como en casi todos los dinosaurios y ciertamente en todos los ornistisquios , los dientes fueron reemplazados continuamente en un arreglo alternativo, con las dos ondas de reemplazo moviéndose desde la parte posterior hacia la parte frontal de la mandíbula. El espaciado de Z, la distancia promedio en la posición del diente entre los dientes en la misma etapa de erupción, fue bastante bajo con Hyspilophodon , aproximadamente 2,3. Dicha dentición habría permitido procesar plantas relativamente duras.

Los primeros paleontólogos modelaron el cuerpo de este dinosaurio herbívoro pequeño, bípedo y pequeño de varias maneras. En 1882, Hulke sugirió que el Hypsilophodon era cuadrúpedo pero, en vista de su mano, podía trepar rocas y árboles para buscar refugio.[14] En 1912 esta línea de pensamiento fue perseguida por el paleontólogo austríaco Othenio Abel. Concluyendo que el primer dedo del pie del pie podía funcionar como un hallux oponible, Abel declaró que el Hypsilophodon era un animal totalmente arbóreo e incluso que un estilo de vida arbóreo era primitivo para los dinosaurios en su conjunto.[23] Aunque esta hipótesis fue puesta en duda por Nopcsa,[24] fue adoptada por el investigador danés Gerhard Heilmann, quien en 1916 propuso que un Hypsilophodon cuadrúpedo viviera como el moderno canguro arborícola Dendrolagus.[25] En 1926, Heilmann volvió a cambiar de opinión, negando que el primer dedo fuera opuesto porque el primer metatarsiano estaba conectado firmemente al segundo,[26] pero en 1927 Abel se negó a aceptar esto.[27] En 1936, fue apoyado por Swinton, quien afirmó que incluso un primer metatarsiano apuntando hacia adelante podría llevar un dedo movible.[16] Como Swinton fue un popularizador de dinosaurios muy influyente, este fue el punto de vista aceptado durante más de tres décadas, y la mayoría de los libros ilustran al Hypsilophodon sentado en la rama de un árbol. Sin embargo, Peter M. Galton en 1969 realizó un análisis más preciso de la estructura musculoesquelética, mostrando que la postura del cuerpo era horizontal.[28] En 1971, Galton refutó detalladamente los argumentos de Abel, demostrando que el primer dedo del pie se había reconstruido incorrectamente y que ni la curvatura de las garras, ni el nivel de movilidad del hombro o la cola podían verse como adaptaciones para escalar,[28] concluyendo que Hypsilophodon tenía una forma bípeda de moverse.[29] Esto convenció a la comunidad paleontológica de que Hypsilophodon permanecía firmemente en el suelo.
El nivel de cuidado parental en este dinosaurio no se ha definido, no se han encontrado nidos, aunque se conocen nidos ordenados de especies relacionadas, lo que sugiere que se tomó cierto cuidado antes de la eclosión. Aunque los fósiles de Hypsilophodon no formaban parte de una sola manada, los huesos se habían ido acumulando gradualmente, a menudo se consideraba probable que los animales se movieran en grandes grupos. Por estas razones, los hipsilofodóntidos, particularmente Hypsilophodon, a menudo se han referido como el "ciervo del Mesozoico". Algunas indicaciones sobre los hábitos reproductivos son proporcionadas por la posibilidad de dimorfismo sexual, Galton consideró probable que los ejemplares con cinco en lugar de seis vértebras sacras, con algunos especímenes, la vértebra que normalmente debería contar como la primera parte del sacro con una costilla que no toca la pelvis, representan a hembras.[1]
En la cultura popular
Los hipsilofodontes aparecen en las novelas del escritor estadounidense Michael Crichton Parque Jurásico y El mundo perdido como una de las especies de dinosaurio recreados mediante ingeniería genética,[30][31] pero fueron omitidos en la versión cinematográfica.
Referencias
- Galton, P.M., 1974, The ornithischian dinosaur Hypsilophodon from the Wealden of the Isle of Wight. British Museum (Natural History), Bulletin, Geology, London, 25: 1‑152c
- P.M. Galton, 2009, "Notes on Neocomian (Lower Cretaceous) ornithopod dinosaurs from England - Hypsilophodon, Valdosaurus, "Camptosaurus", "Iguanodon" - and referred specimens from Romania and elsewhere", Revue de Paléobiologie, Genève 28(1): 211-273
- Paul, G.S., 2010, The Princeton Field Guide to Dinosaurs, Princeton University Press p. 275
- Hulke, J.W., 1874, "Supplemental note on the anatomy of Hypsilophodon foxii", Geological Society of London, Quarterly Journal, 30: 18-23
- Butler, Richard J.; Galton, Peter M. (2008). «The 'dermal armour' of the ornithopod dinosaur Hypsilophodon from the Wealden (Early Cretaceous: Barremian) of the Isle of Wight: a reappraisal». Cretaceous Research 29 (4): 636-642. doi:10.1016/j.cretres.2008.02.002.
- Mantell, G.A., 1849, "Additional observations on the osteology of the Iguanodon and Hylaeosaurus", Royal Society of London, Philosophical Transactions, 139: 271-305
- Owen, R., 1855, Monograph on the fossil Reptilia of the Wealden and Purbeck formations. Part II. Dinosauria (Iguanodon). [Wealden]. Palaeontographical Society Monographs, London, 7: 1-54
- Fox, W.D., 1868, "On the skull and bones of an Iguanodon", British Association for the Advancement of Science, Annual Report for 1867, London, 38: 64-65
- Huxley, T.H., 1869, "On Hypsilophodon, a new genus of Dinosauria", Geological Society of London, Abstracts of Proceedings, 204: 3-4
- Huxley, T.H., 1870, Plantilla:Ws, Geological Society of London, Quarterly Journal, 26: 3-12
- Darren Naish, 2009, The Great Dinosaur Discoveries, A & C Black, London, p. 46
- Owen, R., 1874, Monograph on the fossil Reptilia of the Wealden and Purbeck formations. Supplement No. 5. Dinosauria (Iguanodon). [Wealden and Purbeck]. Palaeontographical Society Monographs 27, pp 1-18
- Hulke, J.W., 1873, Plantilla:Ws, Geological Society of London, Quarterly Journal, 29: 522-532
- Hulke, J.W., 1882, "An attempt at a complete osteology of Hypsilophodon foxii, a British Wealden dinosaur", Royal Society of London, Philosophical Transactions, 172: 1053-1062
- Nopcsa, F., 1905, "Notes on British dinosaurs. Part I. Hypsilophodon", Geological Magazine, London, (5) 2: 203-208
- Swinton, W.E., 1936, "Notes on the osteology of Hypsilophodon, and on the family Hypsilophodontidae", Zoological Society of London, Proceedings, 1936: 555-578
- Galton, P.M., 1967, On the anatomy of the ornithischian dinosaur Hypsilophodon foxii from the Wealden (Lower Cretaceous) of the Isle of Wight, England. unpublished Ph.D. Thesis, University of London, King’s College, UK, 513 pp
- Galton, P.M. & J.A. Jensen, 1978, "Remains of ornithopod dinosaurs from the Lower Cretaceous of North America", Brigham Young University, Geology Studies, Provo, 25(3): 1-10
- Galton, P.M. & J.A. Jensen, 1975, "Hypsilophodon and Iguanodon from the Lower Cretaceous of North America", Nature 257: 668-669
- Norman, David B.; Sues, Hans-Dieter; Witmer, Larry M.; Coria, Rodolfo A. (2004). «Basal Ornithopoda». En Weishampel, David B.; Dodson, Peter; Osmólska, Halszka, eds. The Dinosauria (2nd edición). Berkeley: University of California Press. pp. 393–412. ISBN 978-0-520-24209-8.
- Madzia, D.; Boyd, C.A.; Mazuch, M. (2017). «A basal ornithopod dinosaur from the Cenomanian of the Czech Republic». Journal of Systematic Palaeontology 16 (11): 967-979. doi:10.1080/14772019.2017.1371258.
- L. Dollo, 1882, "Première note sur les dinosaures de Bernissart", Bulletin du Musée Royale d'Histoire Naturelle de Belgique 1: 1-18
- Abel, O., 1912, Grundzüge der Palaeobiologie der Wirbeltiere, E. Schweizerbart'sche Verlagsbuchhandlung Nägele und Dr Sproesser, Stuttgart
- V. Pietschinann (ed), 1914, Verhandlungen der Kaiserlich-Königlichen Zoologisch-Botanischen Gesellschaft in Wien, Vienna, pp 380
- Heilmann., G., 1916, Vor nuværende Viden om Fuglenes Afstamning, Copenhagen
- Heilmann, Gerhard, 1926, The Origin of Birds. London, Witherby
- Abel, O., 1927, Lebensbilder aus der Tierwelt der Vorzeit, Fisher, Jena
- Galton, P.M., 1971, "Hypsilophodon, the cursorial nonarboreal dinosaur", Nature, 231: 159-161
- Galton, P.M., 1971, "The mode of life of Hypsilophodon, the supposedly arboreal ornithopod dinosaur", Lethaia, 4: 453-465
- Chrichton, Michael (1995) [1990]. Parque Jurásico (5 edición). Barcelona: Plaza & Janés. ISBN 84-01-49202-5.
- Chrichton, Michael (1997) [1995]. El Mundo Perdido (2 edición). Barcelona: Plaza & Janés. ISBN 84-01-49202-5.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikimedia Commons alberga una galería multimedia sobre Hypsilophodon foxii.
Wikimedia Commons alberga una galería multimedia sobre Hypsilophodon foxii. Wikispecies tiene un artículo sobre Hypsilophodon foxii.
Wikispecies tiene un artículo sobre Hypsilophodon foxii.- Más datos sobre el Hypsilophodon
- Excelente diseño e interesantes características del Hypsilophodon en Yahooligans!
- Rendición de un artista (en inglés)
| Control de autoridades |
|
|---|
 Datos: Q131086
Datos: Q131086- Multimedia: Hypsilophodon / Q131086