Tenontosaurus
Tenontosaurus ("lagarto de tendón") es un género con dos especies de dinosaurios ornitópodos iguanodontianos, que vivieron a mediados del período Cretácico, entre aproximadamente 122 a 108 millones de años desde el Aptiense al Albiense, en lo que es hoy Norteamérica.
| Tenontosaurus | ||
|---|---|---|
| Rango temporal: 122 Ma - 108 Ma Cretácico Inferior | ||
 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Ornithischia | |
| Suborden: | Neornithischia | |
| Infraorden: | Ornithopoda | |
| Género: |
Tenontosaurus Ostrom, 1970 | |
| Especie tipo | ||
|
Tenontosaurus tilletti Ostrom, 1970 | ||
| Otras Especies | ||
| ||
| Sinonimia | ||
| ||
El género contiene dos especies, Tenontosaurus tilletti, descrito por John Ostrom en 1970[1] y Tenontosaurus dossi,descrito por Winkler, Murry y Jacobs en 1997.[2] Se han recolectado muchos especímenes de T. tilletti de varias formaciones geológicas en todo el oeste de América del Norte. T. dossi se conoce sólo por un puñado de especímenes recolectados en la Formación Twin Mountains del condado de Parker, Texas, Estados Unidos .
Descripción

Los tenontosaurios median entre 6,5 y 8 metros de longitud, 2,2 de altura y pesaba de 1 a 2 toneladas. En 2010, Gregory S. Paul le dio a T. tillettorum una longitud más corta de 6 metros con un peso de 600 kilogramos. T. dossi habría sido un poco más grande con siete metros y una tonelada. La longitud no es una buena indicación del peso en Tenontosaurus porque la cola es muy larga.[3] En 2016 dio valores de 6 a7 metros de largo y entre 600 a 1000 kilogramos.[4] Presentaban extremidades delanteras largas con manos provistas de cinco rollizos dedos, cráneo curvo y mediano, cuerpo robusto y corpulento, cola larga lo era más que los otros miembros de su familia y era el doble del largo del torso, que al igual que su espalda estaba rígida con una red de tendones óseos, cosa que lo habría obligado a andar mucho tiempo en cuatro patas, Su cabeza terminaba en pico con muelas alineadas a los lados y sus piernas eran largas con patas provistas de cuatro rollizos dedos.
Cráneo
El cráneo de Tenontosaurus es bajo y largo, muy alargado cuando se ve desde arriba, con la punta del hocico mostrando un ensanchamiento redondeado conspicuo y los lados inferiores de la parte posterior muy anchos. El hocico también es fuertemente redondeado en vista lateral con un perfil convexo que se extiende hasta las cuencas de los ojos. En la curva del hocico hay una enorme fosa nasal ovalada inclinada hacia atrás que se extiende oblicuamente hacia atrás en una gran depresión alargada que ahueca todo el lado superior del hocico. Debajo del extremo posterior de este, en el costado del hocico, solo hay una pequeña fenestra antorbital, un óvalo que también se inclina hacia atrás. La cuenca del ojo es relativamente pequeña y de inserción alta. Detrás de ella y un poco más abajo hay una pequeña ventana redondeada en la parte inferior de la sien que es más pequeña que la cuenca del ojo. Debajo, en muchos especímenes, hay una segunda abertura, más pequeña, en la pared lateral, en el borde del hueso cigomático y el cuadradoyugal. Esto se ve como una ventana de sueño inferior secundaria. Las ventanas temporales superiores emparejadas en el techo del cráneo tienen forma de riñón y son alargadas.[5]
El premaxilar abarca casi toda la fosa nasal desde abajo. La pared lateral del premaxilar también se profundiza profundamente por la depresión alrededor de la fosa nasal. Los premaxilares juntos forman un núcleo óseo del pico superior que cuelga por debajo del borde de la mandíbula. Estos son desdentados en T. tillettorum, esto es cierto para la mayoría de los Iguanodontia, pero si T. dossi realmente representa un ancestro directo con dientes de T. tillettorum, este rasgo debe haber evolucionado por separado en este último ya que es poco probable que sea el ancestro del resto del grupo. En la vista superior, esta mandíbula tiene forma de U. Sus paredes laterales son más anchas que los bordes de la mandíbula y están ligeramente hundidas en la parte posterior. Se superponen a las ramas anteriores de los huesos maxilares que, a su vez, superponen las partes anteriores planas de los huesos vomer, una configuración quizás única. Las alas internas de los premaxilares forman un paladar duro en el pico, que, sin embargo, todavía tiene muescas en los huesos vomer. La rama ascendente del premaxilar no se superpone al hueso nasal, sino que encaja en un surco en su costado, una característica basal. Ambas ramas ascendentes hacen una muesca en los huesos nasales aproximadamente a la mitad de su longitud. La rama larga del premaxilar debajo de la fosa nasal toca el hueso lagrimal, excluyendo el hueso maxilar del hueso nasal de Zalmoxes. La cuenca del ojo es aproximadamente triangular con el borde de ataque formando una línea recta vertical y el borde de salida redondeado. El hueso frontal es solo una pequeña parte del borde superior de la cuenca del ojo, una característica derivada. La rama posterior del maxilar se articula con el hueso cigomático. El quadratoyugal es grande y el hueso cigomático no toca el cuadrado, una característica basal. Igualmente básica es la ausencia de un agujero cuadrado y una muesca en el borde anterior del cuadrado; la ventana de sueño inferior secundaria puede ser un precursor de esto. El cuadrado es largo y transversalmente aplanado con un margen posterior ligeramente convexo. El escamoso toca internamente el pómulo. En la parte posterior de la cabeza se encuentran los procesos paroccipitales enganchados y doblados hacia abajo en sus extremos. En la pared lateral de la caja craneana, el hueso parietal no toca el proótico, una característica básica. El basioccipital toca los laterosfenoides.[5]
Las mandíbulas inferiores tienen un predentario común en sus frentes, el núcleo óseo del pico inferior, en Tenontosaurus esta es una estructura poco profunda en forma de herradura que encaja en la parte superior del pico desde abajo y está colocada con pseudodientes, protuberancias del margen óseo que se extendía por una capa córnea. Los dientes dentarios verdaderos de la mandíbula inferior tienen una cresta principal vertical prominente. Los dientes maxilares de la mandíbula superior no tienen ningún reborde principal, sino varios rebordes inferiores que no son del todo paralelos. El dentario con dientes tiene un margen superior e inferior paralelos, una característica derivada. Detrás del dentario está el surangular, que está atravesado por un solo agujero, no por dos como en los euornitópodos más basales. La mandíbula tiene una proyección retroarticular larga en la parte posterior a la que el músculo depresor de la mandíbula permitió que se abrieran las mandíbulas.[5]
Esqueleto axial
La columna vertebral consta de doce vértebras cervicales, quince vértebras dorsales, seis vértebras sacras y al menos sesenta y una vértebras de la cola. Algunos investigadores cuentan dieciséis vértebras dorsales y cinco vértebras sacras, sin contar una vértebra dorsosacral como parte del sacro, esta vértebra no tiene una costilla sacra ensanchada fusionada con el "yugo". La cola es muy larga y constituye al menos el 57% de la longitud del animal, en algunos casos hasta dos tercios. La columna vertebral está rígida por tendones osificados que recorren la espalda, el sacro y la cola anterior y media. En la cola también hay tales tendones sobre los chevrones en la parte inferior.[6]
El cuello es moderadamente largo con costillas largas y apófisis espinosas bajas. En comparación con la parte delantera baja del torso, el cuello se curva hacia arriba con bastante fuerza, lo que lleva la cabeza a la altura de la espalda. Las apófisis espinosas de las vértebras anteriores también son bajas, pero más atrás en la serie se vuelven algo más altas, llegando a ser dos veces más altas que anchas de adelante hacia atrás. En el centro de la espalda son bastante anchos en vista lateral con un perfil rectangular. Las apófisis espinosas de las vértebras sacras son aún más largas y se inclinan bastante hacia atrás. En tres vértebras sacras, las costillas se fusionan a la perfección en un yugo, en las dos posteriores las costillas, aunque ensanchadas, permanecen separadas. Estos pueden verse como vértebras caudosacral.[6]
La base de la cola se inclina un poco y los procesos espinosos continúan elongándose. Para mantener una conexión con las vértebras anteriores, las apófisis espinosas se curvan ligeramente hacia adelante con sus extremos superiores. Donde la cola se dobla hacia arriba nuevamente, esa curvatura termina. Las apófisis espinosas alcanzan aquí su mayor longitud en la columna vertebral, alrededor de la undécima vértebra. Eso lleva a una cresta en la base de la cola trasera. Los extremos superiores de los procesos espinosos también están algo ensanchados aquí. Más posteriormente, las apófisis espinosas pierden altura rápidamente y el tercio posterior de la cola, muy larga, es delgado. Las proyecciones emparejadas en la parte inferior de la cola, los cheurones o arcos hemales, son muy cortos al frente. Ganan longitud muy rápidamente y ya entre la quinta y la sexta vértebra el cheurón es más largo que las apófisis espinosas de esas vértebras. Así, la cresta en la base trasera de la cola tiene su contraparte en una depresión aún mayor de la mitad inferior de la cola. Esto hace que la cola sea casi tan alta como todo el fuselaje en este punto. Los cheurones aquí, teniendo en cuenta la curvatura de la cola, son casi verticales y se ensanchan gradualmente hacia sus extremos inferiores. Más hacia atrás, gradualmente se vuelven más cortos y más anchos en la parte inferior, y eventualmente desaparecen por completo en el extremo delgado de la cola. Los tendones osificados recorren las apófisis espinosas y los cheurones hasta llegar a este extremo, lo que debe haber convertido a la cola en una estructura muy rígida, presumiblemente para soportar los enormes músculos retractores que tiraban de los fémures hacia atrás al caminar. En comparación con estas proyecciones verticales, las proyecciones laterales de la base de la cola son bastante cortas. Sobresalen en ángulo recto y terminan completamente alrededor de la décima vértebra.[6]
Extremidades anteriores
El escapula tiene un borde de ataque recto y un borde de salida mucho más curvo. Esto da lugar a una extensión superior que sobresale más hacia atrás. El coracoides tiene una poderosa punta posterior y está perforado por un foramen coracoideo que no toca la costura ósea con la escápula. También es bastante bajo. Se han encontrado grandes esternones osificados en forma de media luna en varios especímenes.[6]
La extremidad anterior es larga y robusta en aproximadamente el 68% de la longitud de la extremidad posterior. Tenontosaurus es quizás el euornitópodo conocido más basal que casi con certeza podría caminar a cuatro patas, para Rhabdodon, esta habilidad a veces se cuestiona. El húmero robusto tiene una cresta deltopectoral bastante baja pero poderosa. El antebrazo es bastante largo y el cúbito en particular es robusto, notablemente más ancho que el radio , aunque los procesos olecranianos no está muy bien desarrollado en la parte superior trasera. Hay tres huesos carpos en la muñeca. La mano es corta y ancha con cinco dedos. El ancho es una vez y media el largo. Los dedos cuarto y quinto son bastante cortos. El pulgar tiene una primera falange ensanchada y está de lado pero no lleva una garra grande. Las garras de las manos son cortas pero no tienen forma de pezuña. La fórmula de las falanges es 2-3-3-2-2 o 2-3-3-1-1. Por lo tanto, el tercer dedo tiene tres falanges en lugar de cuatro, como todavía tenían los ancestros de Tenontosaurus.[6]
Extremidades posteriores
El ilion tiene una hoja anterior larga y baja que se curva hacia abajo. En la vista superior, las hojas delanteras se abren gradualmente en abanico ligeramente hacia un lado. El dorso tiene una llamativa protuberancia convexa en el borde superior que está rugosa pero no transversalmente engrosada ni curvada. Las hojas traseras se doblan moderadamente en vista en planta. El borde inferior de la hoja trasera tiene un perfil cóncavo en vista lateral. El surco en el margen inferior para la inserción del músculo caudofemoral breve es bastante estrecho transversalmente, es menos de una vez y media el grosor del borde inferior de la pared interna en este punto. El cuerpo principal del hueso púbico forma una barra recta, corta y delgada, aproximadamente una vez y media más larga que el proceso prepúbico y forma un ángulo de 100° con él. Las barras de ambos lados no están fusionadas en la parte inferior, una característica derivada. El proceso prepúbico aplanado transversalmente se extiende mucho más allá de la hoja anterior del ilion y se curva ligeramente hacia arriba, estrechándose anteriormente. En algunos especímenes, el extremo anterior está ligeramente ensanchado. El foramen obturador no toca la costura de la pierna con el isquion. El isquion es recto y muy ancho. Tiene un proceso obturatorio en forma de media luna a un tercio de la parte superior. Es transversalmente aplanado y tiene una cresta prominente en la parte superior exterior del eje. El extremo está ligeramente ensanchado y justo por encima de ese extremo hay una elevación baja en el borde de fuga.[6]
El fémur es bastante recto con solo una ligera protuberancia hacia adelante. En la parte superior del fémur, el trocánter menor tiene forma de dedo y está separado del trocánter mayor posterior por una fisura estrecha. El trocánter menor tiene aproximadamente un tercio del grosor del trocánter mayor y una quinta parte de la longitud del eje. El cuarto trocánter se coloca posteriormente en el centro del eje y está caído. En las cúspides articulares inferiores, el surco anterior es poco profundo y el posterior profundo. La tibia es ligeramente más corta que el fémur, lo que indica que Tenontosaurus no era un corredor veloz. El metatarso es bastante corto. El quinto hueso metatarsiano es un elemento torcido vestigial. El primer metatarsiano, por otro lado, se acerca a la longitud de los otros metatarsianos y tiene un dedo poderoso con una gran garra. Las garras de los pies son generalmente bastante largas. La fórmula de las falanges es 2-3-4-5-0.[6]
Descubrimiento e investigación
El primer fósil de Tenontosaurus, algunas vértebras de un euornitópodo, el espécimen AMNH 5854, fue encontrado en el condado de Big Horn, Montana, por una expedición del Museo Americano de Historia Natural en 1903. Regresó al área en 1927, 1931 y 1933, desenterrando numerosos fósiles de Tenontosaurus , en el sitio de Middle Dome en el condado de Wheatland , Montana . Eran parte de toda una fauna que Brown pretendía describir pero, como tantos de sus proyectos, nunca llegó a hacerlo. Sin embargo, Charles Langen 1938 se instaló un esqueleto en el Museo Americano de Historia Natural, AMNH 3034, pero eso era muy inexacto, los tendones osificados de la base de la cola se habían eliminado para que la cola no sobresaliera hacia atrás sino que descansara ondulada en el suelo. El cráneo perdido se complementó con un modelo imaginario que tenía poco en común con la realidad. Estas excavaciones desenterraron 18 especímenes más, y cuatro especímenes fueron encontrados durante la década de 1940. A pesar de la gran cantidad de especímenes fósiles, el animal no fue nombrado ni descrito científicamente durante este tiempo, aunque Barnum Brown del AMNH le dio el nombre informal "Tenantosaurus", "lagarto tendón", en referencia al extenso sistema de rigidez de los tendones en su espalda y cola.[7]
Durante la década de 1960, la Universidad de Yale comenzó una excavación extensa a largo plazo en el área de la cuenca Big Horn de la Formación Cloverly de Montana y Wyoming. La expedición estuvo dirigida por John Ostrom, cuyo equipo descubrió más de 40 nuevos especímenes. Después de su expedición, Ostrom se convirtió en el primero en describir y nombrar al animal, llamándolo Tenontosaurus, una ligera variación en la ortografía del nombre informal de Brown. El nombre del género se deriva del griego clásico τένων, tetoon , "tendón", una referencia a los tendones osificados que endurecen la columna vertebral en la pelvis y la base de la cola. Brown ya había planeado nombrar al animal "Tenantosaurus kaiseni" en honor a su colaborador Peter Kaisen. La otra ortografía probablemente se basó en un error tipográfico. El nombre específico honra a la familia de Lloyd Tillett que siempre había ofrecido una gran hospitalidad a las expediciones de Ostrom.[1] Porque este estaba formado por varias personas, George Olshevsky en 1991 propuso cambiar el nombre de la especie por el de Tenontosaurus tillettorum.[8] Fue una cosa que no prosperó en la comunidad científica. El holotipo, AMNH 3040, se ha encontrado en el depósito Himes de la Formación Cloverly que data del Albiense, hace unos 110 millones de años, consiste en un esqueleto parcial sin cráneo. Los paratipos son YPM 5456, un cráneo y YPM-PU 16338, un segundo esqueleto parcial.
Desde 1970, se han reportado muchos más especímenes de Tenontosaurus, Las piezas, al menos mínimamente definidas, incluyen AMNH 5854, vértebras y un posible hueso coracoides. AMNH 3010, un esqueleto parcial. AMNH 3013, un tronco con extremidades. AMNH 3020, huesos del pie. AMNH 3012, la mitad izquierda del torso y las extremidades de un animal joven. AMNH 3014, un esqueleto parcial encontrado junto con un espécimen y cáscaras de huevo de Deinonychus, AMNH 3015. AMNH 3011, un esqueleto parcial. AMNH 3017 un esqueleto parcial. AMNH 3031 un esqueleto parcial. AMNH 3034 una mandíbula inferior, la columna vertebral desde la parte delantera trasera hasta la base de la cola y una pata trasera. AMNH 3043 un esqueleto parcial. AMNH 3044, fragmentos óseos de todas las partes del esqueleto. AMNH 3045, un esqueleto fragmentario. AMNH 3050, piezas de mandíbula inferior y tres garras. AMNH 3062, una serie de treinta y tres vértebras caudales; AMNH 3061, un esqueleto parcial. AMNH 3063, un esqueleto parcial. OMNH 10132, un esqueleto parcial. YPM 16514, un esqueleto parcial. BB #1, un esqueleto parcial. YPM 5459, un esqueleto parcial. YPM 5460, un esqueleto parcial. YPM 5458, un esqueleto parcial. YPM 5471, una calavera. YPM 5472, una calavera. YPM 5461, un esqueleto parcial. YPM 5478, un esqueleto parcial. YPM 5466, un esqueleto parcial. YPM 5457, un esqueleto parcial. YPM 5462, un esqueleto parcial.[9]
Junto con hallazgos posteriores, se asignaron a la especie unos 80 especímenes solo de la Formación Cloverly, tanto adultos como juveniles. Un espécimen importante es OU 10144, un esqueleto sin cráneo de un animal recién nacido, informado en 1990 por Catherine Forster como parte de una descripción completa del esqueleto poscraneal de Tenontosaurus.[6] Además, se han informado hallazgos más grandes de la Formación Paluxy de Texas y más restos de Idaho, Utah, Maryland, Arizona y Oklahoma, todos del Cretácico Inferior. Se conoce todo el esqueleto, pero aún no se ha descrito la mayor parte del material.
Los restos fósiles de este dinosaurio han sido descubiertos en Estados Unidos, exactamente en Montana, Oklahoma, Wyoming, Texas y Arizona. La primera especie, T. tilletti, fue encontrada en Wyoming y Montana en la Formación Cloverly y en la Formación Antlers en el sureste Oklahoma, , la Formación Paluxy de Texas , la Formación Wayan de Idaho, la Formación Cedar Mountain de Utah y la Formación Arundel de Maryland. Años después se descubrió la segunda especie, T. dossi, en la Formación Twin Mountains que data del Aptiense, del condado Parker de Texas, la cual fue nombrada por Winkler, Murray y Jacobs en 1997. El nombre específico honra al ranchero James Doss en cuyo rancho se encontró el holotipo, FWMSH 93B1. Doss donó el esqueleto al Museo de Ciencia e Historia de Fort Worth. Consiste en un esqueleto parcial.[2] Se ha asignado a la especie un segundo esqueleto, FWMSH 93B2. La validez de la especie y su identidad con Tenontosaurus son controvertidas.
En 2013, Jonathan Tennant describió el esqueleto poscraneal utilizando el espécimen LL.12275. En 2015, el cráneo se describió con gran detalle utilizando un nuevo espécimen, OMNH 58340, encontrado en 2000.[5] En 2019, se describieron anomalías debidas a infecciones de este espécimen.[10] En 2021, se describió en detalle la mano de este espécimen.[11]
Clasificación
La posición de Tenontosaurus en el árbol genealógico evolutivo fue muy problemática y controvertida. Faltaba un análisis cladístico adecuado. La estructura ligera de Tenontosaurus recuerda a un hipsilofodonte, pero un examen más detallado ha revelado que es muy probable que sea un miembro basal de Iguanodontia.[12]
El siguiente cladograma sigue un análisis de Butler et al, 2011.[12]
| Ornithopoda |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiología
Dieta
Los Tenontosaurus, al igual que otros géneros relacionados, presentaban comportamientos gregarios. Durante el Cretácico, las plantas con flores estaban limitadas geográficamente, por lo que es probable que este dinosaurio se alimentara de las plantas predominantes de la era, tales como helechos, cicadáceas y coníferas. La vida vegetal en el ecosistema de Tenontosaurus probablemente estuvo dominada por helechos, helechos arborescentes, cícadas y, posiblemente, angiospermas primitivas. Las plantas y árboles más grandes consistieron en gimnospermas, como coníferas y ginkgos. Tenontosaurus era un ramoneador bajo, y un adulto habría tenido una altura máxima de alimentación de unos 3 metros si adoptara una postura bípeda. Esto restringió a Tenontosaurus, especialmente los juveniles, a comer helechos y arbustos de bajo crecimiento. Sin embargo, su potente pico en forma de U y las superficies de corte en ángulo de sus dientes significaban que no estaba limitado a consumir una parte concreta de la planta. Las hojas, la madera e incluso la fruta pudieron haber formado parte de su dieta.[7]
Reproducción
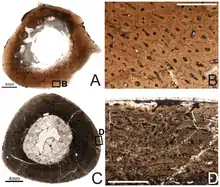
La presencia de tejido óseo medular en el fémur y la tibia de un espécimen indica que los tenontosaurios usaron este tejido, que hoy solo se encuentra en aves que están poniendo huevos, en la reproducción. Además, al igual que Tyrannosaurus y Allosaurus, otros dos dinosaurios que se sabe que produjeron hueso medular, el individuo no alcanzó el tamaño adulto completo tras su muerte a los 8 años. Debido a que la línea de dinosaurios terópodos que incluye a Allosaurus y Tyrannosaurus divergió de la línea que condujo a Tenontosaurus muy temprano en la evolución de los dinosaurios, esto sugiere que los dinosaurios en general produjeron tejido medular y alcanzaron la madurez reproductiva antes de alcanzar su tamaño máximo.[13] Un estudio histológico mostró que T. tilletti creció rápidamente durante sus primeros años de vida y en su etapa de sub-adulto, tras lo cual ralentizava su ritmo de crecimiento en los años que se aproximaban a la madurez, a diferencia de otros iguanodontes.[14]
Depredadores
Se han descubierto algunos dientes de Deinonychus en especímenes de Tenontosaurus tilletti, indicando relaciones tróficas ya seana de depredación o carroñeo entre dichos dinosaurios.[15][16][17]

A menudo se han descubierto dientes y varios esqueletos pertenecientes al terópodo carnívoro Deinonychus asociado con Tenontosaurus tilletti. Se han encontrado especímenes de tenontosaurus en más de 50 sitios, y 14 de ellos también contienen restos de Deinonychus. De acuerdo con un estudio de 1995, solo seis sitios que contienen fósiles de Deinonychus no contienen rastro de Tenontosaurus, y los restos de Deinonychus raramente se encuentran asociados con otras presas potenciales, como Sauropelta.[15] En total, el 20% de los fósiles de Tenontosaurus se encuentran muy cerca de Deinonychus, y varios científicos han sugerido que esto implica que Deinonychus era el principal depredador de Tenontosaurus. Sin embargo, el Deinonychus adulto era mucho más pequeño que el Tenontosaurus adulto, y es poco probable que un solo Deinonychus hubiera sido capaz de atacar a un Tenontosaurus completamente desarrollado. Si bien algunos científicos han sugerido que Deinonychus debe haber sido un cazador gregario, esta opinión se ha cuestionado en función de la falta de evidencia para la caza coordinada (en lugar del comportamiento de acoso como en la mayoría de las aves y reptiles modernos), así como evidencia de que Deinonychus podría mostrar un cierto grado de canibalismo entre individuos, así como también con el Tenontosaurus, en un frenesí alimenticio.[17] Es probable que Deinonychus cazase al Tenontosaurus juvenil, y que cuando este alcanzó un cierto tamaño, pasó fuera del rango como fuente de alimento para los pequeños terópodos, aunque es posible que hayan eliminado a individuos más grandes. El hecho de que la mayoría de los tenontosaurus se encuentran con Deinonychus son individuos medio crecidos que admiten esta visión.[7][16] También vivía en la misma área que el gran dinosaurio carnívoro Acrocanthosaurus.[18]
Paleoecología

En la Formación Cloverly, el Tenontosaurus es, con mucho, el vertebrado más común; cinco veces más abundante que el siguiente más común, el ankylosaurio Sauropelta. En el árido Miembro Little Sheep Member, Tenontosaurus es el único dinosaurio herbívoro, y compartió su ambiente con el depredador Deinonychus, así como una especie indeterminada de terópodo alosauroideo y cocodrilos goniofolídidos. Después del gran cambio climático a mediados de la era albiense, varios dinosaurios ingresaron a la región, incluyendo el ornitópodo Zephyrosaurus, el oviraptorosaurio Microvenator y una especie indeterminada de saurópodo titanosauriformes y un ornitomímido sin identificar. La comunidad ecológica en el estadio tropical también incluyó al pequeño mamífero Gobiconodon, tortugas como Glyptops y diversas especies de peces pulmonados.[7]
La comunidad ecológica fue similar en otras regiones, con dinosaurios como Tenontosaurus y Deinonychus como los grandes vertebrados más comunes. La Formación Antlers se extiende desde el suroeste de Arkansas, a través del sureste de Oklahoma, hasta el noreste de Texas. Esta formación geológica no ha sido fechada radiométricamente. Los científicos han utilizado datos bioestratigráficos y el hecho de que comparte varios de los mismos géneros que el Grupo Trinity de Texas, para suponer que esta formación se estableció durante la etapa albiense del período Cretácico temprano, aproximadamente 110 millones de años. El área preservada en esta formación era una gran llanura de inundación que drenaba en un mar interior poco profundo. Varios millones de años más tarde, este mar se expandiría hacia el norte, convirtiéndose en el Mar Interior Occidental y dividiendo a Norteamérica en dos durante casi todo el período del Cretácico Superior. El paleoambiente de la Formación Antlers consistía en bosques tropicales o subtropicales, llanuras aluviales, deltas fluviales, pantanos costeros, pantanos y lagunas, probablemente similar a la Luisiana moderna.[19][20] En la formación Antlers, en lo que hoy es Oklahoma, Tenontosaurus y Deinonychus compartieron su paleoambiente con otros dinosaurios, como los saurópodos Astrodon (Pleurocoelus) y Sauroposeidon proteles, y el carnosaurio Acrocanthosaurus atokensis, que probablemente fue el superdepredador de esta región.[21][16] El dinosaurio más común en el paleoambiente de esta formación es Tenontosaurus. Otros vertebrados presentes incluyen al anfibio Albanerpeton arthridion, los reptiles Atokasaurus metarsiodon y Ptilotodon wilsoni, el reptil crurotarso Bernissartia, los peces cartilaginosos Hybodus buderi y Lissodus anitae, el actinopterigios Gyronchus dumblei, el cocodrilo Goniopholis y las tortugas Glyptops y Naomichelys.[22][23] Posibles restos indeterminados de aves también se conocen en la Formación Antlers. La evidencia fósil sugiere que el pejelagarto Lepisosteus fue el vertebrado más común en esta región. Los primeros mamíferos conocidos de esta región incluyen Atokatherium boreni y Paracimexomys crossi.[24]
Medio ambiente
En la Formación Cloverly de Montana y Wyoming, los restos de Tenontosaurus son comunes en dos unidades de roca distintas: el miembro de Little Sheep Member (unidad de formación Cloverly V) y el más reciente Miembro Himes (unidades VI y VII). La parte más antigua de la formación, el Conglomerado Pryor, no contiene fósiles de Tenontosaurus, y solo aparecen en la parte más alta y más reciente del Miembro Little Sheep Member. Catherine Forster, en un artículo de 1984 sobre la ecología de Tenontosaurus, usó esto como evidencia para sugerir que las poblaciones de Tenontosaurus no llegaron al área de la Cuenca de Bighorn hasta el momento del Miembro Little Sheep Member tardío.[7]
En el momento en que Tenontosaurus apareció por primera vez en Wyoming y Montana (albiense temprano), el clima de las regiones era entre árido y semiárido, seco, con períodos de lluvias estacionales y sequías ocasionales. Sin embargo, durante unos pocos millones de años, el clima en la región cambió a uno de mayor precipitación, y el ambiente se volvió subtropical o tropical, con deltas fluviales, llanuras inundables y bosques con ensenadas pantanosas que recuerdan a la actual Luisiana,[19] si bien las marcadas estaciones secas también persistieron para crear ambientes similares a la sabana. El cambio en los niveles de precipitaciones probablemente se deba a la costa que avanza en la "Skull Creek Seaway", una etapa de la formación del Mar de Niobrara.[7]
Este cambio dramático en el clima coincidió con un aumento aparente, en lugar de una disminución, en la abundancia de Tenontosaurus. Esto muestra que Tenontosaurus ha sido un animal notablemente adaptable, que persistió durante un largo período de tiempo en un área a pesar de los cambios en su entorno.[7]
Referencias
- J. H. Ostrom. 1970. Stratigraphy and paleontology of the Cloverly Formation (Lower Cretaceous) of the Bighorn Basin area, Wyoming and Montana. Peabody Museum Bulletin 35:1-234
- Winkler, Dale A, Phillip A Murry and Louis L Jacobs. (1997). "A New Species of Tenontosaurus (Dinosauria: Ornithopoda) from the Early Cretaceous of Texas". Journal of Vertebrate Paleontology.
- Paul, G.S., 2010, The Princeton Field Guide to Dinosaurs, Princeton University Press p. 286
- Paul, Gregory S. (2016). The Princeton Field Guide to Dinosaurs. Princeton University Press. p. 311. ISBN 978-1-78684-190-2. OCLC 985402380.
- Thomas, D. Andrew, 2015, "The cranial anatomy of Tenontosaurus tilletti Ostrom, 1970 (Dinosauria, Ornithopoda)", Palaeontologia Electronica 18.2.37A: 1-99
- Catherine A. Forster, 1990, "The Postcranial Skeleton of the Ornithopod Dinosaur Tenontosaurus tilletti", Journal of Vertebrate Paleontology 10(3): 273-294
- Forster, C.A. (1984). "The paleoecology of the ornithopod dinosaur Tenontosaurus tilletti from the Cloverly Formation, Big Horn Basin of Wyoming and Montana." The Mosasaur, 2: 151–163.
- G. Olshevsky. 1991. A revision of the parainfraclass Archosauria Cope, 1869, excluding the advanced Crocodylia. Mesozoic Meanderings 2:1-196
- http://www.paleofile.com/Dinosaurs/Ornithopods/Tenotosaurus.asp
- T.C. Hunt; J.E. Peterson; J.A. Frederickson; J.E. Cohen; J.L. Berry, 2019, "First documented pathologies in Tenontosaurus tilletti with comments on infection in non-avian dinosaurs", Scientific Reports 9: Article number 8705
- Hunt T.C., Cifelli R.L. & Davies K.L. 2021. "The hand of Tenontosaurus tilletti (Dinosauria, Ornithopoda)". Journal of Vertebrate Paleontology. e1938591
- Richard J. Butler, Jin Liyong, Chen Jun, Pascal Godefroit (2011). «The postcranial osteology and phylogenetic position of the small ornithischian dinosaur Changchunsaurus parvus from the Quantou Formation (Cretaceous: Aptian–Cenomanian) of Jilin Province, north-eastern China». Palaeontology 54 (3): 667-683. doi:10.1111/j.1475-4983.2011.01046.x. Parámetro desconocido
|doi-access=ignorado (ayuda) - Lee, Andrew H.; Werning, Sarah (2008). «Sexual maturity in growing dinosaurs does not fit reptilian growth models». Proceedings of the National Academy of Sciences 105 (2): 582-587. doi:10.1073/pnas.0708903105.
- Werning, S. (2012). «The Ontogenetic Osteohistology of Tenontosaurus tilletti». En Farke, Andrew A, ed. PLoS ONE 7 (3): e33539. PMC 3314665. PMID 22470454. doi:10.1371/journal.pone.0033539.
- Maxwell, W. D.; Ostrom, J.H. (1995). «Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations». Journal of Vertebrate Paleontology 15 (4): 707-712. (abstract)
- Brinkman, Daniel L.; Cifelli, Richard L., Czaplewski, Nicholas J. (1998). «"First Occurrence of Deinonychus antirrhopus (Dinosauria: Theropoda) from the Antlers Formation (Lower Cretaceous: Aptain-Albian) of Oklahoma"». Oklahoma Geological Survey (164): 27.
- Roach, B. T.; D. L. Brinkman (2007). «A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs». Bulletin of the Peabody Museum of Natural History 48 (1): 103-138. doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2.
- D'Emic, Michael D.; Melstrom, Keegan M.; Eddy, Drew R. (2012). «Paleobiology and geographic range of the large-bodied Cretaceous theropod dinosaur Acrocanthosaurus atokensis». Palaeogeography, Palaeoclimatology, Palaeoecology. 333–334: 13-23. doi:10.1016/j.palaeo.2012.03.003.
- Wedel, M. J.; Cifelli, R. L. (2005). «Sauroposeidon: Oklahoma’s Native Giant» (PDF). Oklahoma Geology Notes 65 (2): 40-57. Archivado desde el original el 5 de julio de 2008. Consultado el 7 de julio de 2007.
- Forster, C. A. (1984). «The paleoecology of the ornithopod dinosaur Tenontosaurus tilletti from the Cloverly Formation, Big Horn Basin of Wyoming and Montana». The Mosasaur 2: 151-163.
- Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; and Noto, Christopher R. (2004). "Dinosaur Distribution", in The Dinosauria (2nd), p. 264.
- Nydam, R.L. and R. L. Cifelli. 2002a. Lizards from the Lower Cretaceous (Aptian-Albian) Antlers and Cloverly formations. Journal of Vertebrate Paleontology. 22:286–298.
- Cifelli, R. Gardner, J.D., Nydam, R.L., and Brinkman, D.L. 1999. Additions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous), southeastern Oklahoma. Oklahoma Geology Notes 57:124-131.
- Kielan-Jarorowska, Z., and Cifelli, R.L. 2001. Primitive boreosphenidan mammal (?Deltatheroida) from the Early Cretaceous of Oklahoma. Acta Palaeontologica Polonica 46: 377-391.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikispecies tiene un artículo sobre Tenontosaurus.
Wikispecies tiene un artículo sobre Tenontosaurus. Wikimedia Commons alberga una categoría multimedia sobre Tenontosaurus.
Wikimedia Commons alberga una categoría multimedia sobre Tenontosaurus.- Muchos datos acerca del Tenontosaurus
- Algo de información sobre el Tenontosaurus
- Excelente diseño e interesantes características del Tenontosaurus en Yahooligans!
| Control de autoridades |
|
|---|
 Datos: Q131103
Datos: Q131103- Multimedia: Tenontosaurus / Q131103