Deinonychus antirrhopus
Deinonychus antirrhopus (gr. "Garra terrible con contrapeso") es la única especie del género extinto Deinonychus de dinosaurio celurosauriano dromeosáurido, que vivió hace unos 115 a 108 millones de años entre el Aptiense y el Albiense a mediados del período Cretácico, en lo que hoy es Norteamérica. Esta especie podría crecer hasta 3,4 metros de largo. Los fósiles se han recuperado de los estados de Estados Unidos de Montana, Utah, Wyoming y Oklahoma, en rocas de la formación Cloverly, formación Cedar Mountain y formación Antlers, aunque los dientes que pueden pertenecer a Deinonychus han sido encontrados mucho más al este, en Maryland.
| Deinonychus antirrhopus | ||
|---|---|---|
| Rango temporal: 115 Ma - 108 Ma Cretácico Inferior | ||
 Molde de esqueleto montado, Museo Field | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Archosauria | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| Infraorden: | Coelurosauria | |
| (sin rango): | Maniraptora | |
| Superfamilia: | Deinonychosauria | |
| Familia: | Dromaeosauridae | |
| Subfamilia: | Velociraptorinae | |
| Género: |
Deinonychus Ostrom, 1969 | |
| Especie: |
D. antirrhopus Ostrom, 1969 | |
| Sinonimia | ||
| ||
El estudio del paleontólogo John Ostrom de Deinonychus realizado a finales de la década de 1960 revolucionó la manera como los científicos pensaban acerca de los dinosaurios, lo que llevó al "renacimiento de los dinosaurios" e inició el debate sobre si los dinosaurios eran de sangre caliente o de sangre fría. Antes de esto, la concepción popular de dinosaurios había sido el de reptiles gigantes de andar con paso pesado. Ostrom observó el cuerpo pequeño, elegante, de postura horizontal, columna vertebral estrucioniforme, y sobre todo las garras rapaces agrandadas en los pies, lo que sugiere un depredador ágil y activo.[1]
El significado del nombre del género, "Garra terrible", se refiere a la garra inusualmente grande, en forma de hoz en el segundo dedo de cada pata trasera. El fósil YPM 5205 conserva una gran garra ungueal, fuertemente curvada. En vida, los arcosaurios presentaban una vaina córnea sobre este hueso, lo que extendía su longitud. Ostrom observó cocodrilos y garras de aves y reconstruyó la garra para YPM 5205 de más de 120 milímetros de largo.[1] El nombre de la especie antirrhopus significa "contrapeso", que se refiere a la idea de Ostrom sobre la función de la cola. Al igual que en otros dromeosáuridos, las vértebras de la cola tienen una serie de tendones osificados y apófisis súper alargadas en las vértebras. Estas características hacen suponer que la cola sea un contrapeso rígido, pero un fósil de Velociraptor mongoliensis muy estrechamente relacionado, el ejemplar IGM 100/986 tiene un esqueleto de cola articulado que se curva lateralmente en forma de una larga S. Esto sugiere que, en vida, la cola podría doblarse hacia los lados con un alto grado de flexibilidad.[2] Tanto en las formaciones de Cloverly como de Antlers, se han encontrado restos de Deinonychus estrechamente asociados con las del ornitópodo Tenontosaurus. Los dientes descubiertos asociados a especímenes de Tenontosaurus implica que estos eran cazados, o al menos formaban parte declaraciones dieta en forma de carroña, de Deinonychus.
Descripción
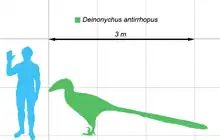
Basado en los pocos ejemplares completamente maduros,[3] Deinonychus podría alcanzar los 3,4 metros de largo, con un cráneo de alrededor de 410 milímetros, una altura de 87 centímetros a las caderas y un peso de 73 kilogramos aunque hay estimaciones más altas de hasta 100 kg.[4][5]
Cráneo
El cráneo estaba equipado con poderosas mandíbulas bordeadas de alrededor de setenta dientes como cuchillas curvas. Los estudios del cráneo han progresado mucho durante las últimas décadas. Ostrom lo reconstruyó a partir de ejemplares parciales, preservados imperfectamente, describiéndolos como triangulares, amplios, y bastante similares a los del Allosaurus. Material adicional del cráneo de Deinonychus y géneros estrechamente vinculados encontrados con una buena preservación tridimensional[6] muestran que el paladar estaba más abovedado volviéndolo más estrecho que lo que pensó Ostrom, haciendo el hocico más angosto, mientras que el yugal se expandía hacia lateral, daba una mayor visión estereoscópica. El cráneo de Deinonychus era diferente en la forma al de Velociraptor, con una bóveda craneana como la de Dromaeosaurus, y no tenía los nasales deprimidos de Velociraptor.[7] Tanto el cráneo como la mandíbula inferior tenían fenestras, aberturas de cráneo, que reducían el peso del cráneo. En Deinonychus la ventana anteorbital, una abertura del cráneo entre el ojo y las narinas, eran particularmente grandes.[6]
Extremidades

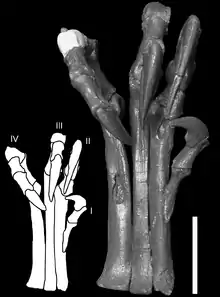
Deinonychus poseía grandes "manos" con tres garras en cada extremidad delantera. El primer dígito era más corto y el segundo era más largo. Cada pata trasera llevaba una garra en forma de hoz en el segundo dígito, lo que probablemente fuese utilizada durante la depredación.[8]
Recubrimiento de plumas
Ninguna impresión de piel han sido encontradas nunca en asociación con fósiles de Deinonychus. Sin embargo, la evidencia sugiere que los Dromaeosauridae, incluyendo Deinonychus, tenían plumas.[9] El género Microraptor es tanto filogenéticamente más primitivo como geológicamente más antiguo que Deinonychus, pese a estar dentro de la misma familia.[10] Múltiples fósiles de Microraptor preservan plumas pennáceas, o sea, plumas con álabes como las de las aves modernas en los brazos, las piernas y la cola, junto con plumas encubiertas y de contorno.[9] Velociraptor es geológicamente más joven que Deinonychus, pero aun así está más estrechamente relacionado, dentro de la subfamilia velociraptorinae, como se explica más abajo. Un espécimen de Velociraptor se ha encontrado con las perillas de inserción de plumas en el cúbito. Las perillas de inserción están donde se unen los ligamentos foliculares, y son un indicador directo de plumas de aspecto moderno.[11]
Descubrimiento e investigación

Los restos fosilizados de Deinonychus han sido recuperados de la formación Cloverly de Montana y Wyoming[1] y en la formación Antlers de Oklahoma,[12] en América del Norte. La formación Cloverly ha sido fechada a finales del Aptiano tomando como referencia las primeras etapas de Albiano del Cretácico temprano, alrededor de 115 a 108 Ma.[13][14] Además, dientes encontrados en las capas de arcilla de la formación Arundel del Aptiano medio y de la formación Potomac en la llanura costera Atlántica de Maryland pueden ser asignados a este género.[15]
Hallazgos iniciales
Los primeros restos en ser encontrados, se hallaron en 1931 en el sudeste de Montana cerca del pueblo de Billings. Un grupo liderado por el paleontólogo Barnum Brown, se concentró sobre todo en excavar y preparar los restos del ornitópodo Tenontosaurus, pero en su informe de campo sobre el sitio de excavación para el Museo Americano de Historia Natural, informaron el descubrimiento de un pequeño dinosaurio carnívoro cerca del esqueleto de Tenontosaurus, "pero muy encajado en la roca lo que lo hacía difícil de preparar".[16] Brown llamó informalmente al animal "Daptosaurus agilis" e hizo preparaciones para describirlo y lo colocó en la exhibición, bajo el número de catálogo AMNH 3015, pero nunca terminó este trabajo.[17] Posteriormente B. Brown volvió de la Formación Cloverly con el esqueleto de un terópodo más pequeño con aparentemente los dientes de gran tamaño, al que nombró informalmente como "Megadontosaurus". John Ostrom revisó el material varias décadas más tarde, identificando a los dientes como pertenecientes a un Deinonychus y el esqueleto proveniente de un animal totalmente distinto al que nombró Microvenator.[17]
Descubrimiento y descripción
Un poco más de treinta años más tarde, en agosto de 1964, el paleontólogo John Ostrom llevó a una expedición del Museo Peabody de la Universidad de Yale que descubrió más material esquelético. Las expediciones durante los dos veranos siguientes desenterraron más de 1000 huesos, entre los cuales estaban por lo menos tres individuos. Puesto que la asociación entre los varios huesos recuperados era débil, fue imposible determinar cual era el número exacto de animales individuales, el espécimen tipo, YPM 5205, de Deinonychus fue restringido a un pie izquierdo completo y un pie derecho parcial que pertenecieron definitivamente al mismo individuo.[8] Los especímenes restantes fueron catalogados en cincuenta entradas separadas en el Museo Peabody de Historia Natural de Yale.
Estudios posteriores de Ostrom y Meyer analizaron su propio material así como el "Daptosaurus" de B. Brown detalladamente y encontraron que era la misma especie. J. Ostrom publicó su descubrimiento en 1969, dándole a todos los restos referido al nombre de Deinonychus antirrhopus.[8] El nombre del género significa en griego: δεινός, 'terrible' y ὄνυξ, con el genitivo ὄνυχος 'garra'. El nombre específico "antirrhopus", del griego ἀντίρροπος significa "contrapeso" y se refiere a los probables efectos probables de una cola rígida.[18] En julio de 1969, Ostrom publicó una extensa monografía sobre Deinonychus.[8]
Aunque ya desde antes de 1969 ya estaban disponibles miles huesos, muchos de los huesos importantes faltaban o eran difíciles de interpretar. Había pocos elementos postorbitales del cráneo, no había ningún fémur, sacro, espoleta o esternón, faltaban vértebras y Ostrom pensó que solo tenía un pequeño fragmento de la coracoides. La reconstrucción esquelética de J. Ostrom de Deinonychus incluyó un hueso pélvico muy inusual, un pubis trapezoidal y plano, diferente al de los otros terópodos, pero que era de la misma longitud que el isquion y que se encontraba junto a este.[8]
Descubrimientos posteriores
En 1974 Ostrom publicó otra monografía sobre el hombro de Deinonychus, en la cual corrigió que el pubis que él había descrito era realmente un coracoides, un elemento del hombro.[19] En ese mismo año, otro espécimen de Deinonychus fue excavado en Montana por una expedición de la Universidad de Harvard dirigida por Farish Jenkins. Este descubrimiento agregó varios nuevos elementos como fémures bien preservados, pubis, un sacro y mejores iliones, así como los elementos del pie y del metatarso. Ostrom describió este espécimen y revisó la restauración esquelética de Deinonychus. Esta vez se vieron pubis muy largos, y Ostrom comenzó a sospechar que pudieron incluso haber estado orientados hacia atrás un poco como en las aves.[20]

Un esqueleto de Deinonychus que incluye los huesos del espécimen original y más completo puede verse en el Museo Americano de Historia Natural,[21] con otro ejemplar en el Museo de Zoología Comparada de la Universidad de Harvard. Los especímenes del Museo Americanos y de Harvard son de un lugar distinto que los de Yale. Incluso estos dos montajes esqueléticos carecen de elementos incluyendo los esternones, las costillas esternales, la espoleta, y gastralias.[22]
Incluso después de todo el trabajo de J. Ostrom, varios pequeños bloques de material incrustado en la roca seguía estando sin preparación en el almacén del AMNH. Estos consistieron sobre todo en huesos y fragmentos aislados de huesos, incluidos en la matriz original, o en la roca circundante en la cual los especímenes terminaron enterrados inicialmente. Un examen de estos bloques sin preparación de Gerald Grellet-Tinner y de Peter Makovicky en 2000 reveló una característica interesante, pasada por alto. Varios huesos largos y finos identificados en los bloques como tendones osificados, estructuras que ayudaron a volver rígida la cola de Deinonychus.w resultaron ser realmente gastralias, costillas abdominales. Pero lo más significativo , una gran cantidad de cáscaras de huevo fosilizadas previamente inadvertidas fueron descubiertas en la matriz de la roca que había rodeado el espécimen original de Deinonychus.[23]
En un informe posterior, más detallado sobre las cáscaras de huevo, Grellet-Tinner y Makovicky concluyeron que los huevos pertenecieron casi ciertamente a Deinonychus, representando el primer huevo del dromeosáurido que se identificara.[16] Por otra parte, la superficie externa de una cáscara de huevo fue encontrada en contacto cercano con las gastralias que sugeriría que este Deinonychus pudo haber estado incubando sus huevos. Esto implica que Deinonychus realizaba un traspaso térmico del cuerpo como mecanismo para la incubación del huevo, e indica endotermia similar a las aves actuales.[24] El estudio adicional de Gregory Erickson y colegas encuentra que este individuo tenía entre 13 o 14 años a la muerte y su crecimiento se había detenido. A diferencia de otros terópodos, en su estudio de los especímenes encontrados asociado con los huevos o nidos, había acabado el crecimiento a la hora de su muerte.[25]
Implicaciones
La descripción de 1969 hecha por Ostrom de Deinonychus ha sido descrito como uno de los hechos más importantes de la paleontología de mediados de siglo XX.[26] El descubrimiento de este depredador ágil y claramente activo hizo mucho para cambiar la concepción científica y popular de los dinosaurios y abrió la puerta a la especulación de que los dinosaurios pueden haber sido de sangre caliente. Este cambio de paradigma ha sido llamado el renacimiento de los dinosaurios. Varios años después, Ostrom notó similitudes entre las patas delanteras de Deinonychus y las de los pájaros, una observación que lo llevó a revivir la hipótesis de que las aves descienden de los dinosaurios.[27] Cuarenta años después, esta idea es casi universalmente aceptada.
El hallazgo de los primeros restos de muchos de Deinonychus por los paleontólogos John Ostrom y Grant E. Meyer en 1964, en el sur de Montana, llevaron a Ostrom a suponer que el Deinonychus vivía en manadas, algo impensable para la época, que juzgaba a los dinosaurios como animales de limitada inteligencia que sólo podrían llevar una vida solitaria. También dio con las primeras evidencias de que Deinonychus depredaban sobre los Tenontosaurus; algo que, se supone, no podrían hacer en solitario.

Deinonychus estaba seguramente emplumado como su cercano pariente Velociraptor, demostrado en 2007 con base en las marcas de las plumas en los brazos. Tales marcas demuestran donde se anclan las plumas en pájaros actuales. Este descubrimiento reciente agregó el peso a las teorías ya fuertes que todos los dromeosáuridos estaban emplumados.[11]
Debido a su anatomía extremadamente similar a ave y estrecha relación con otros dromeosáuridos, los paleontólogos suponen que probablemente Deinonychus estuviera recubierto de plumas.[28][29][30] Existe clara evidencia fósil de plumas del estilo de las aves modernas para varios dromeosáuridos relacionados, incluyendo Velociraptor y Microraptor, aunque no haya evidencia directa conocida para Deinonychus.[9][11] Cuando se realizaban estudios de áreas tales como la amplitud de movimiento en las extremidades anteriores, los paleontólogos como Phil Senter asumieron como probable presencia de plumas de vuelo, que se presentan en todos dromaeosaurios conocidos con impresiones de piel.[31]
Clasificación
Deinonychus es uno de los mejor conocidos entre los Dromaeosauridae,[32] junto al más pequeño y relativamente cercano Velociraptor, del Campaniano de Asia Central.[33][34] El clado que ellos forman se llama Velociraptorinae. El nombre Velociraptorinae fue acuñado por primera vez por Rinchen Barsbold en 1983[35] y originalmente contenía solo al Velociraptor. Posteriormente, Phil Currie incluyó a más dromeosáuridos.[36] Del Cretácico superior a Tsaagan de Mongolia[33] y al Saurornitholestes norteamericano,[4] pueden también estar entre los familiares cercanos, pero este último es mal conocido y difícil de clasificar.[33] Velociraptor y sus parientes usaron sus garras más que sus cráneos como herramientas de caza, en comparación con dromeosáuridos como Dromaeosaurus de cráneos más masivos.[4] Junto con Troodontidae, los dromeosáuridos del clado Deinonychosauria son taxones hermanos de las aves. Filogenéticamente, Deinonychosauria es el grupo de dinosaurios no avianos más cercano a las aves.[37] En un análisis de 2015 de los paleontólogos Robert DePalma, David Burnham, Larry Martin, Peter Larson y Robert Bakker, utilizando datos actualizados del Grupo de Trabajo Teróxido coloca a Deinonychus como basal en Dromaeosaurinae.[38]
Filogenia
El cladograma muestra la posición filogenética de Deinonychus dentro de Eudromaeosauria de acuerdo con el análisis de Evans y sus colegas en 2013.[39]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Cladograma basado en el estudio de 2015.

| Dromaeosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiología
Comportamiento predador
.jpg.webp)
Los dientes de Deinonychus encontrados en asociación con fósiles del dinosaurio ornitópodo Tenontosaurus son muy comunes en la formación Cloverly. Se han descubierto dos sitios que preservan fósiles bastantes completos de Deinonychus cerca a Tenontosaurus. El primero, el sitio de Yale en Cloverly, Montana, incluye numerosos dientes, cuatro fósiles de Deinonychus adultos y un juvenil. La asociación de este número de esqueletos de Deinonychus en un solo lugar sugiere que Deinonychus puede haberse alimentado de ese animal, y quizás haberlo cazado. Ostrom y Maxwell incluso utilizaron esta información para especular que Deinonychus pudo haber vivido y cazado en manadas.[40] El segundo sitio es en la formación Antlers de Oklahoma. El sitio contiene seis esqueletos parciales de Tenontosaurus de varios tamaños, junto con un esqueleto parcial y muchos dientes de Deinonychus. Un húmero del Tenontosaurus incluso tiene lo que pueden ser marcas de dientes de Deinonychus. D. Brinkman et al.. En 1998 precisan que Deinonychus tenía un peso adulto de 70 a 100 kilogramos, mientras que los Tenontosaurus adultos pesaban entre 1 a 4 toneladas. Un solitario Deinonychus no podría matar un Tenontosaurus adulto, sugiriendo que la caza en manadas sería posible.[41]
Un estudio de 2007 por B. T. Roach y D. L. Brinkman han aportado a la cuestión sobre el comportamiento cooperativo durante la caza en manada de Deinonychus, basados en lo que se sabe de la caza de carnívoros modernos y la tafonomía de sitios de Tenontosaurus. Los arcosaurios modernos, aves, cocodrilos y el dragón de Komodo exhiben poca o ninguna forma de caza cooperativa, en su lugar, son generalmente cazadores solitarios, o se agrupan en torno a cadáveres de animales muertos anteriormente, donde ocurre mucho conflictos entre los individuos de la misma especie. Por ejemplo, en las situaciones donde los grupos de dragones de Komodo están comiendo juntos, los individuos más grandes comen primero y atacarán a dragones de Komodo más pequeños que intenten alimentarse. Si se mata al animal más pequeño, este es canibalizado. Cuando esta información se aplica a los sitios de Tenontosaurus, parece que es consistente con lo encontrado entre los dragones de Komodo y cocodrilos. Los restos esqueléticos de Deinonychus encontrados en estos sitios son de subadultos y las piezas que faltan en ellos son consistentes con la predación por otros Deinonychus.[42] Por otra parte, un artículo de Li et al. describen sitios de pista con una separación pie similares y rastros paralelos, lo que implica un comportamiento gregario de manada en lugar de la conducta alimentaria no coordinada.[43]
En 2011, un estudio encabezado por Denver Fowler sugirió un nuevo método de caza de Deinonychus y otros dromeosaurios para alimentarse de presas pequeñas.[44] Este modelo propone que Deinonychus se abalanzaba sobre sus presas como los modernos accipítridos, dejando caer el peso sobre su presa y sujetándola con las garras. Como estas aves, el dromeosaurio empezaría a devorar a su presa aún viva, mientras esta moría por hemorragia. Esta proposición está basada en la comparación de la anatomía de dromeosáuridos con aves depredadoras actuales. Las proporciones de las patas y pies son similares a las de águilas y halcones, mientras que los pies fuertes y los metatarsos cortos de Deinonychus recuerdan a los de los búhos. El método de depredación propuesto sería coherente con otros aspectos de la anatomía de Deinonychus, tales como su morfología inusual de mandíbula y brazo. Los brazos estuvieron probablemente cubiertos de plumas largas, y pudieron haber sido utilizados como estabilizadores de aleteo para el equilibrio mientras el alto de un animal de presa luchaba, junto con la cola de contrapeso rígida. Sus mandíbulas, que se cree que tuvieron una fuerza de mordida comparativamente débil,[45] podrían haber sido utilizadas para las mordeduras de movimiento de sierra, como el dragón de Komodo moderno que también tiene una fuerza de mordida débil, para acabar con su presa, si sus patadas no fueran lo suficientemente potentes.[46]
Fuerza de la mordedura

En 2005 se elaboraron por primera vez las estimaciones de la fuerza de mordida para Deinonychus, basándose en la reconstrucción de la musculatura de la mandíbula. Este estudio concluyó que era probable que Deinonychus haya tenido una fuerza máxima de mordedura de solo el 15% de la del caimán americano moderno.[45] Un estudio de 2010 de Paul Gignac y colegas intentó estimar la fuerza de mordida basada directamente en marcas de punción de dientes de Deinonychus recién descubiertos en los huesos de un Tenontosaurus. Estas marcas provenían de un gran individuo, y proporcionaron la primera evidencia de que los grandes Deinonychus podrían morder hasta los huesos. En cuanto al uso de las marcas de los dientes, el equipo de Gignac fue capaz de determinar que la fuerza de la mordedura de Deinonychus fue significativamente mayor que lo que habían estimado estudios anteriores solo por estudios biomecánicos. Descubrieron que la fuerza de mordida de Deinonychus alcanzada entre 4100 y 8200 newtons de fuerza, mayor que mamíferos carnívoros vivos como la hiena, y equivalente a un cocodrilo de tamaño similar.[47]
Sin embargo, Gignac y sus colegas también observaron que las marcas de punción ósea de Deinonychus son relativamente raras, y a diferencia de los terópodos más grandes con muchas marcas de punciones conocidas, como el Tyrannosaurus, Deinonychus probablemente no mordiera frecuentemente a través de los huesos o los comiera. En cambio, es probable que usaron su gran fuerza de mordida con propósitos de defensa o en la captura de presas, en lugar de alimentarse.[47]
Función de los miembros

A pesar de ser la característica más distintiva de Deinonychus, la forma y la curvatura en hoz de la garra varía entre los especímenes. El espécimen tipo descrito por Ostrom en 1969 tiene una garra fuertemente curvada en hoz, mientras que un espécimen más nuevo descrito en 1976 tenía una garra con una curvatura mucho más débil, más similar en perfil con el de las garras normales en los restantes dedos del pie.[20] Ostrom ha sugerido que esta diferencia en el tamaño y la forma de las garras en hoz podría ser debido a la variación individual, sexual, o relativa a la edad, pero admitió que no podía asegurarlo.
Esta es evidencia anatómica[1] y dactilar[48] de que este talón se levantaba del suelo, mientras que el dinosaurio caminaba sobre el tercer y cuarto dedo del pie.
Ostrom sugirió que Deinonychus podía patear con la garra falciforme para cortar y desgarrar a su presa.[1] Inclusive, algunos investigadores sugirieron que el talón fue utilizado para destripar grandes dinosaurios ceratopsianos.[49] Otros estudios han sugerido que no se utilizaron las garras en hoz para desgarrar, sino para asestar pequeñas puñaladas a su víctima.[50] En 2005, Manning y colegas realizaron pruebas en una réplica robótica que coincide exactamente con la anatomía de Deinonychus y Velociraptor, que utiliza pistones hidráulicos para hacer que el robot golpee un cadáver de cerdo. En estas pruebas, las garras hacen solamente pinchazos superficiales y no pudieron cortar o recortar. Los autores sugirieron que las garras habrían sido más eficaces para escalar que para matar asestando golpes.[51]
Ostrom comparó a Deinonychus con el avestruz y el casuario. Señaló que las especies de aves pueden causar una lesión grave con la gran garra en el segundo dedo del pie.[1] El casuario tiene garras de hasta 125 milímetros de largo.[52] Ostrom citó a Gilliard de 1958 cuando dijo que lo que pueden romper un brazo o destripar a un hombre.[53] Kofron entref 1999 y 2003 estudió 241 ataques documentados de casuarios y encontró que dejó un saldo de un humano y dos perros muertos, pero no hay evidencia de que los casuarios pueden destripar o desmembrar a otros animales.[54][55] Los casuarios usan sus garras para defenderse, atacar y amenazar a los animales y en exhibiciones intraespecíficas de exhibición.[56] El seriema también tiene una segunda garra de pies ampliada, y la utiliza para desgarrar pequeñas presas antes de tragar.[56] En 2011, un estudio sugirió que la garra falciforme probablemente se haya utilizado para mantener fija la presa, mientras la mordía, y no como un arma cortante.[44]

Estudios biomecánicos de Ken Carpenter en 2002 confirmaron que la función más probable de los brazos era la de agarrar la presa, pues sus grandes longitudes habrían permitido un alcance más largo que para la mayoría de los otros terópodos. El grande y alargado coracoides hace suponer la existencia de músculos de gran fuerza en los brazos, lo que consolida esta teoría.[57] Los estudios biomecánicos de Carpenter usando moldes de los huesos también demostraron que los Deinonychus no podían doblar sus brazos contra su cuerpo como un pájaro, contrariamente a lo que fue deducido en 1985 de las descripciones anteriores por Jacques Gauthier[58] y Gregory S. Paul en 1988.[4]
Los estudios de Phil Senter en 2006 indicaron que los brazos de Deinonychus se podían utilizar no sólo para agarrar sino también para llevar los objetos hacia el pecho. La presencia probable de un ala grande emplumada en Deinonychus, sin embargo, habría limitado en un cierto grado la gama de movimiento de los brazos. Por ejemplo, cuando Deinonychus llevaba sus brazos hacia adelante, la palma de la mano giraba automáticamente a una posición hacia arriba. Esto habría hecho que un ala bloqueara a la otra si ambos brazos estuviesen extendidos al mismo tiempo, llevando a Senter a la concluir el hecho de atraer objetos al pecho habría sido logrado solamente con un brazo a la vez. La función de los dedos también habría estado limitada por las plumas, por ejemplo, en actividades tales escarbar en las grietas para buscar pequeñas presa solamente se habría podido emplear el tercer dígito de la mano, y solamente en perpendicular a la posición del ala.[31] Alan Gishlick, en un 2001 estudio de la mecánica del brazo de Deinonychus, encontró que incluso si las plumas grandes del ala estaban presentes, la capacidad de atrape de la mano no habría sido obstaculizada perceptiblemente. Esta habría sido perpendicular al ala, y sería sostenida probablemente por ambas manos simultáneamente en un "abrazo de oso", hecho apoyado por los estudios posteriores del brazo por Carpenter y Senter.[59] En un estudio de 2001 realizado por Bruce Rothschild y otros paleontólogos, se examinaron 43 huesos de la mano y 52 huesos del pie que se refieren a Deinonychus, para detectar signos de fractura por estrés, no encontrándose ninguna.[60] La segunda falange del segundo dedo del pie en el espécimen YPM 5205 presenta una fractura curada.[61]
Parsons y Parsons han demostrado que los especímenes juveniles y adultos jóvenes de Deinonychus muestran algunas diferencias morfológicas con los adultos. Por ejemplo, los brazos de los ejemplares más jóvenes fueron proporcionalmente más largos que los de los adultos, una posible indicación de diferencia de comportamiento entre los jóvenes y los adultos.[62] Otro ejemplo de esto podría ser la función de las garras del pie. Parsons y Parsons han sugerido que la curvatura de la garra, que Ostrom [1976] ya había mostrado que fue diferente entre los especímenes,[20] tal vez fue mayor para Deinonychus juveniles, ya que esto podría ayudar a subir a los árboles, y que las garras se hicieron más rectas a medida que el animal aumentaba de tamaño y comenzaba a vivir exclusivamente de la tierra.[63] Esto se basaba en la hipótesis de que algunos pequeños dromeosáuridos usaban sus garras del pie para escalar.[51] En un documento de 2015, tras un nuevo análisis de fósiles inmaduros se informó que el carácter abierto y móvil de la articulación del hombro podría haber significado que los Deinonychus jóvenes eran capaces de realizar alguna forma de vuelo.[64]
Velocidad

Los dromeosáuridos, especialmente Deinonychus, son representados por lo general como animales inusualmente de rápidos en los medios masivos y Ostrom mismo especuló que Deinonychus era veloz en su descripción original.[8] Sin embargo, cuando el primer Deinonychus fue descrito, carecía de una pierna completa, y Ostrom especuló sobre la longitud del fémur, que más adelante se comprobó que estaba sobredimensionado. En un estudio posterior, Ostrom observó que el cociente del fémur a la tibia no es tan importante en la determinación de velocidad como la longitud relativa del pie y de la parte inferior de la pierna. En las aves modernas, como el avestruz, el cociente entre el pie y la tibia es 0,95. En dinosaurios inusualmente de rápidos como el Struthiomimus, el cociente es 0,68, pero en Deinonychus, el cociente es de 0,48. Ostrom indicó que "solamente una conclusión es razonable, el Deinonychus no era particularmente rápido comparado con otros dinosaurios, ni con las aves corredoras modernas".[20]
El bajo cociente del miembro posterior distal en Deinonychus es debido en parte a un inusualmente corto metatarso. El cociente es realmente más grande en individuos más pequeños que en los más grandes. J. Ostrom sugirió que el metatarso corto se pueda relacionar con la función de la garra falciforme, y utilizó el hecho de que parece hacerse más corto mientras que los individuos envejecen como soporte de esto. Interpretó todas estas características, el segundo dedo del pie corto con la garra agrandada, el metatarso corto, etc. como ayuda para el uso de la pierna trasera como arma ofensiva, donde la garra giraría hacia abajo y atrás, y de la pierna empujaba hacia abajo y detrás al mismo tiempo, desgarrando en la presa. Ostrom sugirió que el metatarso corto reducía la tensión total en los huesos de la pierna durante tal ataque, e interpretó el arreglo inusual de las inserciones musculares en la pierna del Deinonychus como apoyo para su idea que un sistema de músculos distinto fuera utilizado en el movimiento rapaz y en caminar. Por lo tanto, Ostrom concluyó que las piernas de Deinonychus representaban un equilibrio entre las adaptaciones corrientes necesarias para un depredador ágil, y las características para compensar ser su arma única del pie.[20]
En su estudio de 1981 de las huellas de dinosaurio encontrada en Canadá, Richard Kool realizó estimaciones burdas sobre la velocidad que caminan basadas en varios rastros de huellas hechas por diversas especie en la Formación Gething de la Columbia Británica. R. Kool estimó para uno de estos rastros de la icnospecies, Irenichnites gracilis, que pudo haber sido de hecho Deinonychus, una velocidad al caminar de 2,8 metros por segundo, alrededor de 10,1 kilómetros por hora.[65]
Huevos

La identificación en 2000 de un probable huevo de Deinonychus asociado a uno de los especímenes originales permitió la comparación con otros dinosaurios terópodos en términos de estructura del huevo, nido, y reproducción. En su estudio de 2006 del espécimen, Grellet-Tinner y Makovicky examinaron la posibilidad que los dromeosáuridos se había estado alimentando de los huevos, o que fragmentos había sido asociado al esqueleto de Deinonychus por coincidencia. Descartaron la idea que el huevo había sido una comida para el terópodo, observando que los fragmentos fueron intercalados entre las gastralias y los huesos del brazo, haciéndolo imposible que representaran el contenido estomacal del animal. Además, la manera de la cual el huevo había sido machacado y hecho fragmento indicando que había estado intacto a la hora del entierro, y estaba quebrado por el proceso de fosilización. La idea que el huevo fuera asociado aleatoriamente al dinosaurio también fue encontrada como inverosímil, los huesos que rodeaban el huevo no habían sido dispersados ni desarticulados, sino seguían estando bastante intactos concerniente a sus posiciones en vida, indicando que el área alrededor al huevo no fueron disturbadas durante la preservación. El hecho de que estos huesos fueran gastralias, que muy raramente se encuentran articulados, apoyó esta interpretación. Toda la evidencia, según Grellet-Tinner y Makovicky, indican que el huevo estaba intacto debajo del cuerpo del Deinonychus cuando fue enterrado. Es posible que este representa el empollamiento o comportamiento de nido en Deinonychus similar a lo visto en trodóntidos y ovirraptóridos, o que el huevo estaba de hecho en el interior del oviducto cuando murió el animal.[16]
El examen de la microestructura del huevo de Deinonychus confirma que perteneció a un terópodo, puesto que comparte características con otros huevos conocidos de terópodos y muestra que diferencias con huevos de ornitisquios y saurópodos. Comparado con otros terópodos maniraptores, el huevo de Deinonychus es más similar a los de oviraptórido que a los de troodóntidos, a pesar de los estudios que demuestran que estos últimos son lo más estrechamente vinculados a los dromeosáuridos como Deinonychus. Mientras que el huevo estaba muy roto para determinar exactamente su tamaño, Grellet-Tinner y Makovicky estimaron un diámetro alrededor de 7 centímetros basado en la anchura del canal pélvico a través del cual el huevo tuvo que haber pasado. Este tamaño es similar al diámetro de 7,2 centímetros del más grande huevo de Citipati, un oviraptórido. Citipati y Deinonychus también compartían el mismo tamaño de cuerpo total, apoyando esta estimación. Además, el grosor de las cáscaras de huevo de Citipati y Deinonychus son casi idénticos, y puesto que el grueso de cáscara se correlaciona con el volumen del huevo, estos datos apoyan la idea que los huevos de estos dos animales eran de tamaño casi igual.[16]
Paleoecología
La evidencia geológica sugiere que Deinonychus habitaba una llanura aluvional o zonas pantanosas.[32] El paleoambiente de ambas formaciones, la Cloverly y la de Antlers, en donde los restos de Deinonychus han sido encontrados, consisten de bosques, deltas fluviales y lagos, no muy diferente a lo que es hoy Luisiana.[66][67] Otros animales que compartieron este mundo con Deinonychus al herbívoro armado Sauropelta, el más grande de los raptores Utahraptor y los ornitópodos Zephyrosaurus y Tenontosaurus. En Oklahoma, el ecosistema de Deinonychus también incluyó al terópodo Acrocanthosaurus, el gigantesco Sauroposeidon, el cocodriliano Goniopholis y peces Lepisosteiformes como Lepisosteus.[67] Si los dientes encontrados en Maryland son de Deinonychus, entonces sus vecinos incluirían a Astrodon, Dryptosaurus y un nodosáurido, llamado Priconodon conocido solamente por los dientes. La porción media de la Formación Cloverly varía en antigüedad desde los 115 ± 10 millones de años cerca de la base[13] a 108,5 ± 0,2 millones de años cerca de la parte superior.[14]
En la cultura popular
Deinonychus ocupó un lugar destacado de la novela Parque Jurásico de 1990 de Michael Crichton y su adaptación cinematográfica en 1993, dirigida por Steven Spielberg. Sin embargo, Crichton y Spielberg optaron por utilizar el nombre del Velociraptor para estos dinosaurios, en lugar de Deinonychus. Durante el proceso de escritura Crichton se reunió, en varias ocasiones, con John Ostrom para discutir los detalles de la posible gama de comportamientos y apariencia viva de Deinonychus. En cierto momento Crichton se disculpó contándole a Ostrom que había decidido utilizar el nombre del Velociraptor en lugar de Deinonychus para su libro, porque sentía que el nombre anterior era "más dramático". A pesar de esto, según Ostrom, Crichton declaró que el Velociraptor de la novela fue basado en Deinonychus en casi todos los aspectos, y que solo el nombre fue cambiado.[68] Los realizadores de Jurassic Park siguieron el ejemplo, diseñando los modelos de la película basándose casi enteramente en Deinonychus en lugar del Velociraptor real, y según los informes solicitados todos los trabajos publicados de Ostrom sobre Deinonychus durante la producción.[68] Como resultado, estos dinosaurios de la película fueron retratados con el tamaño, proporciones y forma de hocico de Deinonychus.[69]
Deinonychus ha aparecido en la franquicia The Land Before Time en la película de 2000 The Land Before Time VII: La piedra del fuego frío, pero solo en una historia durante la película en la que atacan una manada de dinosaurios herbívoros que consiste en Parasaurolophus, 'Ouranosaurus, Styracosaurus y 'Ankylosaurus. Deinonychus también jugó un papel protagónico en documentales sobre dinosaurios, como Clash of the Dinosaurs y Jurassic Fight Club.
Old Lace, compañera de Gertrude Yorkes en el cómic de Marvel y programa de televisión Runaways fue identificada correctamente por Víctor Mancha como Deinonychus después de que el grupo asumió que ella era un Velociraptor.
Véase también
Referencias
- Ostrom, John Harold (1970). «Stratigraphy and paleontology of the Cloverly Formation (Lower Cretaceous) of the Bighorn Basin area, Wyoming and Montana». Bulletin of the Peabody Museum of Natural History (en inglés) 35: 1-234.
- Norell, Mark A.; Makovicky, Peter J. (1999). «Important features of the dromaeosaurid skeleton II: information from newly collected specimens of Velociraptor mongoliensis». American Museum Novitates 3282: 1-45.
- Parsons, William L.; Parsons, Kristen M. (2009). «Further descriptions of the osteology of Deinonychus antirrhopus (Saurischia, Theropoda)». Bulletin of the Buffalo Society of Natural Sciences (en inglés) 38.
- Paul, Gregory Scott (1988). Predatory Dinosaurs of the World (en inglés). Nueva York: Simon and Schuster. pp. 358, 366–369. ISBN 0-671-61946-2.
- Campione, Nicolas E.; Evans, David C.; Brown, Caleb M.; Carrano, Matthew T. (2014). «Body mass estimation in non-avian bipeds using a theoretical conversion to quadruped stylopodial proportions». Methods in Ecology and Evolution (en inglés) 5 (9): 913-923. doi:10.1111/2041-210X.12226.
- Maxwell, W.D.; &Witmer, L.M. (1996). «New Material of Deinonychus (Dinosauria, Theropoda)». Journal of Vertebrate Paleontology. 16(3): 51A.
- Witmer, Lawrence M.; Maxwell, William D. (1996). «The skull of Deinonychus (Dinosauria:Theropoda): New insights and implications». Journal of Vertebrate Paleontology (en inglés) 16 (3): 73A.
- Ostrom, J. H. (1969). «Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana». Peabody Museum of Natural History Bulletin 30: 1-165.
- Xu, X.; Zhou, Z.; Wang, X.; Kuang, X.; Zhang, F.; Du, X. (2003). «Four-winged dinosaurs from China». Nature (en inglés) 421 (6921): 335-340. PMID 12540892. doi:10.1038/nature01342.
- Hwang, S. H.; Norell, M. A.; Ji, Q.; Gao, K. (2002). «New Specimens of Microraptor zhaoianus (Theropoda: Dromaeosauridae) from Northeastern China». American Museum Novitates (en inglés) 3381: 44. doi:10.1206/0003-0082(2002)381<0001:nsomzt>2.0.co;2.
- Turner, Alan H.; Makovicky, Peter J.; Norell, Mark A. (2007). «Feather quill knobs in the dinosaur Velociraptor». Science (en inglés) 317 (5845): 1721. PMID 17885130. doi:10.1126/science.1145076.
- Brinkman, D. L.; Cifelli, R. L.; Czaplewski , N. J. (1998). «First occurrence of Deinonychus antirrhopus (Dinosauria: Theropoda) from the Antlers Formation (Lower Cretaceous: Aptian–Albian) of Oklahoma». Oklahoma Geological Survey Bulletin (en inglés) 146: 1-27.
- Chen, Z.-Q.; Lubin, S. (1997). «A fission track study of the terrigenous sedimentary sequences of the Morrison and Cloverly Formations in northeastern Bighorn Basin, Wyoming». The Mountain Geologist (en inglés) 34: 51-62.
- Burton, D.; Greenhalgh, B.W.; Britt, B.B.; Kowallis, B.J.; Elliott, W.S.; Barrick, R. (2006). «New radiometric ages from the Cedar Mountain Formation, Utah and the Cloverly Formation, Wyoming: implications for contained dinosaur faunas». Geological Society of America Abstracts with Programs (en inglés) 38 (7): 52.
- Lipka, T. R. (1998). «The Affinities of the Enigmatic Theropods of the Arundel Clay Facies (Aptian), Potomac Formation, Atlantic Coastal Plain of Maryland». En Lucas, S.G., Kirkland, J.I. and Estep, J.W., ed. Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History and Science Bulletin, 14 (en inglés). Albuquerque: New Mexico Museum of Natural History and Science. pp. 229-234. OCLC 40283894.
- Grellet-Tinner, G.; and Makovicky, P. (2006). «A possible egg of the dromaeosaur Deinonychus antirrhopus: phylogenetic and biological implications». Canadian Journal of Earth Sciences 43 (6): 705-719. doi:10.1139/E06-033.
- Norell, M. A.; Gaffney, E. S.; Dingus, L. (1995). Discovering Dinosaurs in the American Museum of Natural History. New York: Knopf. pp. 126–130. ISBN 0-679-43386-4.
- Ostrom, John Harold (1969). «A new theropod dinosaur from the Lower Cretaceous of Montana». Postilla (en inglés) 128: 1-17.
- Ostrom, John H.,. (1974). " The Pectoral Girdle and Forelimb Function of Deinonychus (Reptilia: Saurischia) : A Correction". Postilla, Peabody Museum of Natural History Bulletin , 165:1-11.
- Ostrom, J.H. (1976). «On a new specimen of the Lower Cretaceous theropod dinosaur Deinonychus antirrhopus». Breviora 439: 1-21.
- American Museum of Natural History (2007). «Deinonychus». http://www.amnh.org. American Museum of Natural History. Archivado desde el original el 12 de agosto de 2007. Consultado el 13 de julio de 2007.
- Parsons, W.L. & K.M. (2009). "Further Descriptions of the Osteology of Deinonychus antirrhopus (Saurischia, Theropoda)". Bulletin of the Buffalo Society of Natural Sciences. 38.
- Makovicky, P. J.; Grellet-Tinner, G. (2000). «Association between a specimen of Deinonychus antirrhopus and theropod eggshell». En Bravo, A.M. and T. Reyes, ed. First international symposium on dinosaur eggs and babies,Isona i Conca Dellà Catalonia, Spain, 23–26 September 1999. pp. 123-128.
- Grellet-Tinner, Gerard (2006). «Oology And The Evolution Of Thermophysiology In Saurischian Dinosaurs: Homeotherm And Endotherm Deinonychosaurians?». Papeis Avulsos de Zoologia 46 (1): 1-10. Consultado el 7 de julio de 2007.
- Erickson, Gregory M.; Curry Rogers, Kristina; Varricchio, David J.; Norell, Mark A.; Xu, Xing (2007). «Growth patterns in brooding dinosaurs reveals the timing of sexual maturity in non-avian dinosaurs and genesis of the avian condition» (pdf). Biology Letters. published online: 558. doi:10.1098/rsbl.2007.0254. Consultado el 26 de julio de 2007. (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- Fastovsky, D.E., Weishampel, D.B. (2005). «Theropoda I: Nature Red in Tooth and Claw». En Fastovsky, D.E., Weishampel, D.B., ed. The Evolution and Extinction of the Dinosaurs (2nd edition edición). Cambridge: Cambridge University Press. pp. 265-299. ISBN 0-521-81172-4.
- Ostrom, J. H. (1976). «Archaeopteryx and the origin of birds». Biological Journal of the Linnean Society 8 (2): 91-182. doi:10.1111/j.1095-8312.1976.tb00244.x.
- Bakker, R.T. (1986). The Dinosaur Heresies (en inglés). Kensington Publishing. p. 310. ISBN 0-8065-2260-7.
- Long, J.A.; Schouten, P. (2008). «Deinonychus». Feathered Dinosaurs: The Origin of Birds (en inglés). Oxford University Press. pp. 142–143. ISBN 0-19-537266-2.
- Dixon, Dougal (2007). «Fast Hunters». The Illustrated Encyclopedia of Dinosaurs (en inglés). Lorenz Books. pp. 160-161. ISBN 0-7548-1573-0.
- Senter, Phil (2006). «Comparison of Forelimb Function Between Deinonychus And Bambiraptor (Theropoda: Dromaeosauridae)». =Journal of Vertebrate Paleontology (en inglés) 26 (4): 897-906. doi:10.1671/0272-4634(2006)26[897:COFFBD]2.0.CO;2.
- Norell, M.A., Makovicky, P.J. (2004). «Dromaeosauridae». En Weishampel, D.B., Dodson, P., Osmólska, H., ed. The Dinosauria (2a edición). Berkeley: University of California Press. pp. 196–210. ISBN 0-520-24209-2.
- Norell, M.A.; Clark, J.M., Turner, A.H., Makovicky, P.J., Barsbold, R., y Rowe, T. (2006). «A new dromaeosaurid theropod from Ukhaa Tolgod (Ömnögov, Mongolia)» (PDF). American Museum Novitates 3545: 1-51. doi:10.1206/0003-0082(2006)3545[1:ANDTFU]2.0.CO;2. Consultado el 7 de julio de 2007.
- Turner, A.S.; Hwang, S.H.; Norell, M.A. (2007). «A small derived theropod from Öösh, Early Cretaceous, Baykhangor Mongolia» (PDF). American Museum Novitates 3557: 1-27. doi:10.1206/0003-0082(2007)3557[1:ASDTFS]2.0.CO;2. Consultado el 29 de marzo de 2007.
- Barsbold, R. (1983). «Carnivorous Dinosaurs from the Cretaceous of Mongolia». The Joint Soviet-Mongolian Palaeontological Expedition, Transactions 19: 1-117.
- Currie, P. J. (1995). «New information on the anatomy and relationships of Dromaeosaurus albertensis (Dinosauria: Theropoda)». Journal of Vertebrate Paleontology 15 (3): 576-591. (abstract)
- Benton, M.J. (2004). Vertebrate Palaeontology (Third Edition). Blackwell Publishing. p. 472. ISBN 9780632056378. Archivado desde el original el 19 de octubre de 2008. Consultado el 26 de diciembre de 2008.
- DePalma, Robert A.; Burnham, David A.; Martin, Larry D.; Larson, Peter L.; Bakker, Robert T. (2015). «The First Giant Raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation.». Paleontological Contributions (14).
- Evans, D. C.; Larson, D. W.; Currie, P. J. (2013). "A new dromaeosaurid (Dinosauria: Theropoda) with Asian affinities from the latest Cretaceous of North America". Naturwissenschaften. doi:10.1007/s00114-013-1107-5.
- Maxwell, W. D.; Ostrom, J.H. (1995). «Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations». Journal of Vertebrate Paleontology 15 (4): 707-712. (abstract)
- Brinkman, Daniel L.; Cifelli, Richard L., Czaplewski, Nicholas J. (1998). «"First Occurrence of Deinonychus antirrhopus (Dinosauria: Theropoda) from the Antlers Formation (Lower Cretaceous: Aptain-Albian) of Oklahoma"». Oklahoma Geological Survey (164): 27.
- Roach, B. T.; D. L. Brinkman (2007). «A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs». Bulletin of the Peabody Museum of Natural History 48 (1): 103-138. doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2.
- Li, Rihui; Lockley, Martin G.; Makovicky, Peter J.; Matsukawa, Masaki; Norell, Mark A.; Harris, Jerald D.; Liu, Mingwei (2007). «Behavioral and faunal implications of Early Cretaceous deinonychosaur trackways from China». Naturwissenschaften (en inglés) 95 (3): 185-91. PMID 17952398. doi:10.1007/s00114-007-0310-7.
- Fowler, D. W.; Freedman, E. A.; Scannella, J. B.; Kambic, R. E. (2011). «The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds». En Farke, Andrew Allen, ed. PLoS ONE 6 (12): e28964. PMID 22194962. doi:10.1371/journal.pone.0028964.
- Therrien, F.; Henderson, D.M.; Huff, C.B. (2005). «Bite me: biomechanical models of theropod mandibles and implications for feeding behavior». En Carpenter, Kenneth, ed. The Carnivorous Dinosaurs. Indianapolis: Indiana University Press. pp. 179–237. ISBN 978-0-253-34539-4.
- Sakamoto, M. (2010). "Jaw biomechanics and the evolution of biting performance in theropod dinosaurs". Proceedings of the Royal Society B: Biological Sciences 277 (1698): 3327. doi:10.1098/rspb.2010.0794.
- Gignac, P.M.; Makovicky, P.J.; Erickson, G.M.; Walsh, R.P. (2010). «A description of Deinonychus antirrhopus bite marks and estimates of bite force using tooth indentation simulations». Journal of Vertebrate Paleontology (en inglés) 30 (4): 1169-1177. doi:10.1080/02724634.2010.483535.
- Li, Rihui; Lockley, Martin G.; Makovicky, Peter J.; Matsukawa, Masaki; Norell, Mark A.; Harris, Jerald D.; Liu, Mingwei (2007). «Behavioral and faunal implications of deinonychosaur trackways from China». Naturwissenschaften (en inglés) 95 (3): 185-91. PMID 17952398. doi:10.1007/s00114-007-0310-7.
- Adams, Dawn (1987) "The bigger they are, the harder they fall: Implications of ischial curvature in ceratopsian dinosaurs" pp. 1–6 in Currie, Philip J. and Koster, E. (eds) Fourth symposium on mesozoic terrestrial ecosystems. Tyrrell Museum, Drumheller, Canada
- Carpenter, Kenneth (1998). «Evidence of predatory behavior by carnivorous dinosaurs». Gaia 15: 135-144.
- Manning, Phil L.; Payne, David; Pennicott, John; Barrett, Paul M.; Ennos, Roland A. (2006). «Dinosaur killer claws or climbing crampons?». Biology Letters (en inglés) 2 (1): 110-112. PMC 1617199. PMID 17148340. doi:10.1098/rsbl.2005.0395.
- Davies, S. J. J. F. (2002). Ratites and Tinamous. Nueva York: Oxford University Press. ISBN 0-19-854996-2.
- Gilliard, E. T. (1958). Living birds of the world. Garden City, NY: Doubleday.
- Kofron, Chhristopher P. (1999) "Attacks to humans and domestic animals by the southern cassowary (Casuarius casuarius johnsonii) in Queensland, Australia
- Kofron, Christopher P. (2003). «Case histories of attacks by the southern cassowary in Queensland». Memoirs of the Queensland Museum (en inglés) 49 (1): 335-338.
- Redford, Kent H.; Peters, Gustav. «Notes on the biology and song of the red-legged seriema (cariama cristata)». Journal of Field Ornithology (en inglés) 57 (4): 261-269.
- Carpenter, K. (2002). «Forelimb biomechanics of nonavian theropod dinosaurs in predation» (PDF). — 82: 59-76. Archivado desde el original el 27 de septiembre de 2007. Consultado el 7 de julio de 2007.
|revista=y|publicación=redundantes (ayuda) - Gauthier, J.; Padian, K. (1985). "Phylogenetic, Functional, And Aerodynamic Analyses Of The Origin Of Birds And Their Flight" in The Beginnings Of Birds. M.K. Hecht, J.H. Ostrom, G. Viohl, & P. Wellnhofer (eds) proceedings of the International Archaeopteryx Conference, Eichstätt, 1984: 185-197, Eischtatt: Freunde des Jura-Museums Eichstätt. isbn 3-9801178-0-4.
- Gishlick, A.D. (2001). «The function of the manus and forelimb of Deinonychus antirrhopus and its importance for the origin of avian flight». En Gauthier, J. and Gall, L.F., ed. New Perspectives on the Origin and Early Evolution of Birds. New Haven: Yale Peabody Museum. pp. 301-318.
- Rothschild, B., Tanke, D. H., and Ford, T. L., 2001, Theropod stress fractures and tendon avulsions as a clue to activity: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, pp. 331-336.
- Molnar, R. E., 2001, Theropod paleopathology: a literature survey: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, pp. 337-363.
- Parsons, W.; Parsons, K. (2006). «Morphology and size of an adult specimen of Deinonychus antirrhopus, (Saurischia, Theropoda)». Journal of Vertebrate Paleontology 26 (3 sup.): 109A.
- Parsons, W. L.; Parsons, K. M. (2009). «Further descriptions of the osteology of Deinonychus antirrhopus (Saurischia, Theropoda)» (pdf). Bulletin of the Buffalo Society of Natural Sciences 38: 43-54. Archivado desde el original el 3 de julio de 2010. Consultado el 14 de noviembre de 2010.
- Parsons, William L.; Parsons, Kristen M. (2015). «Morphological Variations within the Ontogeny of Deinonychus antirrhopus (Theropoda, Dromaeosauridae)». PLoS ONE 10 (4). doi:10.1371/journal.pone.0121476. e0121476.
- Kool, R. (1981). "The walking speed of dinosaurs from the Peace River Canyon, British Columbia, Canada." Canadian Journal of Earth Science, 18: 823-825.
- Forster, C. A. (1984). «The paleoecology of the ornithopod dinosaur Tenontosaurus tilletti from the Cloverly Formation, Big Horn Basin of Wyoming and Montana». The Mosasaur (en inglés) 2: 151-163.
- Wedel, M. J.; Cifelli, R. L. (2005). «Sauroposeidon: Oklahoma’s Native Giant» (PDF). Oklahoma Geology Notes 65 (2): 40-57. Archivado desde el original el 5 de julio de 2008. Consultado el 7 de julio de 2007.
- Cummings, M. "Yale’s legacy in Jurassic World." Yale News, 18 de junio de 2015.
- Duncan, Jody (2006). The Winston Effect (en inglés). Londres: Titan Books. p. 175. ISBN 1-84576-365-3.
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikimedia Commons alberga una categoría multimedia sobre Deinonychus antirrhopus.
Wikimedia Commons alberga una categoría multimedia sobre Deinonychus antirrhopus. Wikispecies tiene un artículo sobre Deinonychus antirrhopus.
Wikispecies tiene un artículo sobre Deinonychus antirrhopus.- Deinonychus en Dinodata (en inglés, se requiere registrarse en forma gratuita)
- Deinonychus dentro de Dromaeosauridae en The Thescelsosaurus (en inglés)
- Significado y pronunciación del nombre (en inglés)
| Control de autoridades |
|
|---|
 Datos: Q20717749
Datos: Q20717749- Especies: Deinonychus antirrhopus