Acrocanthosaurus atokensis
Acrocanthosaurus atokensis (gr. "lagarto de espinas altas de Atoka") es la única especie conocida del género extinto Acrocanthosaurus de dinosaurio terópodo carcarodontosáurido que existió a mediados del período Cretácico, hace aproximadamente entre 125 y 99,6 millones de años, durante el Albiense,[1] en lo que hoy es Norteamérica. Sus restos fósiles han sido hallados en los estados de Oklahoma y Texas, mientras que dientes atribuidos a este género proceden de Maryland.
| Acrocanthosaurus atokensis | ||
|---|---|---|
| Rango temporal: 125 Ma - 99,6 Ma Cretácico inferior | ||
.jpg.webp) Esqueleto montado de un Acrocanthosaurus (NCSM 14345) en el Museo de Carolina del Norte de Ciencias Naturales. | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| Infraorden: | Carnosauria | |
| Superfamilia: | Allosauroidea | |
| Familia: | Carcharodontosauridae | |
| Género: |
Acrocanthosaurus Stovall & Langston, 1950 | |
| Especie: |
A. atokensis Stovall & Langston, 1950 | |
| Sinonimia | ||
| ||
El acrocantosaurio era un depredador bípedo, y como el nombre indica, se lo conoce principalmente por sus apófisis espinosa, ubicadas en muchas de sus vértebras, y que parecen haber sostenido una "joroba" encima del cuello, espalda y caderas del animal. El acrocantosaurio fue uno de los terópodos más grandes, con unos 11,5 metros de largo, y un peso de hasta 5.7 toneladas.[2] Grandes huellas fósiles descubiertas en Texas pudieron pertenecer al acrocantosaurio, aunque no se las ha asociado directamente con esqueletos.
Una serie de descubrimientos recientes han permitido clarificar muchos detalles de su anatomía, como también realizar estudios detallados de su estructura cerebral y la función de sus miembros delanteros. Sin embargo, persiste el debate sobre sus relaciones evolutivas, mientras que algunos científicos lo clasifican como un alosáurido, otros en cambio sostienen que es un carcarodontosáurido. El acrocantosaurio era el téropodo más grande de su ecosistema y posiblemente un superpredador que dio caza a grandes saurópodos y ornitópodos.
Descripción


Aunque era más pequeño que su pariente el Giganotosaurus, el Acrocanthosaurus fue uno de los terópodos más grandes que han existido. El individuo más grande y completo que se ha descubierto, NCSM 14345 se estima que su longitud fue de 11,5 metros y con un peso estimado de 5,7 a 6,2 toneladas, con un rango máximo de 7,25 toneladas posibles para el espécimen[2][3][4][5][6] y su cráneo medía aproximadamente 1,3 metros de largo.[4]
El cráneo del acrocantosaurio, al igual que el de otros alosáuridos, era largo, bajo y estrecho. Un agujero en frente de su cuenca ocular conocido como fenestra anteorbital, le permitía reducir un tanto su peso, dicha abertura era muy grande y medía más de un cuarto de la longitud craneal y dos tercios de su altura. La superficie externa del maxilar y el lado superior del hueso nasal, justo al tope del hocico, no poseían una textura rugosa como en el giganotosaurio o el carcarodontosaurio. De los huesos nasales partían largos y cortos bordes que recorrían ambos lados del hocico hasta detrás de los ojos, donde continuaban sobre los huesos lagrimales.[4] Esto es un rasgo característico de todos los alosauroides.[7] A diferencia del alosaurio, no se presentaba una prominente cresta sobre el hueso lagrimal que se ubicaba frente a los ojos. Los huesos lagrimales y postorbitales se encuentran formando un arco superciliar encima del ojo, al igual que en los carcarodontosáuridos y los no relacionados abelisáuridos. Diecinueve dientes curvos y aserrados se insertaban a cada lado de la mandíbula superior, pero no se ha publicado aún la cantidad que hay en la mandíbula inferior. Los dientes del acrocantosaurio eran más anchos que los del carcarodontosaurio y no tenían la textura rugosa que distinguía a los carcarodontosáuridos. El dental fue cuadrado fuera del borde frontal, como en el giganotosaurio, y poco profundo, a diferencia del resto de la mandíbula que es muy profunda. El acrocantosaurio y el giganotosaurio poseen un grueso borde horizontal en el lado externo del hueso surangular de la mandíbula inferior, debajo de la articulación temporomandibular.[4]
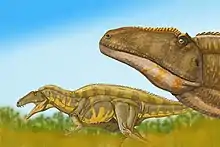
Lo que más destaca al acrocantosaurio es su fila de altas espinas neurales que se localizan en las vértebras desde el cuello hasta el final de la cola, midiendo hasta 2,5 veces la altura de la vértebra.[8] Otros dinosaurios también poseían espinas similares en su espalda, las de algunos siendo mucho más altas que las del acrocantosaurio, como por ejemplo, las del espinosaurio, las cuales medían casi 2 metros de altura y eran 11 veces más altas que el cuerpo de la vértebra.[9] Estas se diferenciaban también de las del acrocantosaurio porque formaban una "vela" de piel, mientras en las bajas espinas del acrocantosaurio se ataban poderosos músculos, como es el caso de los bisontes modernos, que formaban una joroba alta y gruesa.[8] La función de estas espinas permanece desconocida, aunque probablemente pudieron servir para la comunicación, almacenamiento de grasa, o termorregulación. Todas las vértebras cervicales y dorsales tenían grandes depresiones (pleurocelas) a los lados, mientras las caudales se dotaban de unas más pequeñas. Esto es una característica más distintiva de los carcarodontosáuridos que de los alosáuridos.[10]
Sin contar las vértebras, el acrocantosaurio poseía el típico esqueleto de los alosauroides. Era bípedo, con una cola larga y poderosa que balanceaba al cuerpo y la cabeza, manteniendo el centro de gravedad sobre las caderas. Sus brazos eran más cortos y robustos que los del alosaurio pero se asemejaban en otros aspectos al estar provistos de tres dedos con garras en cada mano. A diferencia de muchos dinosaurios veloces más pequeños, el fémur era más largo que la tibia y el metatarso,[4][10] demostrando que el acrocantosaurio no era un rápido corredor.[11] No es de sorprender que los huesos de las patas del acrocantosaurio eran más robustos que los de su pequeño pariente el alosaurio. Cada pie poseía cuatro dedos, de los cuales tres contactaban con el suelo y uno se mantenía al aire.[4][10]
Descubrimiento e investigación
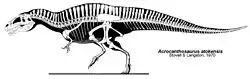
Acrocanthosaurus fue nombrado por sus altas espinas neurales, del griego ακρα/akra ("alta"), ακανθα/akantha ("espina") y σαυρος/sauros ("lagarto").[12] Solo hay una especie descrita (A. atokensis), nombrada por el condado de Atoka en Oklahoma, donde los primeros especímenes fueron encontrados. El nombre fue acuñado en 1950 por los paleontólogos estadounidenses J. Willis Stovall y Wann Langston, Jr.[8] Langston propuso el nombre "Acracanthus atokaensis" en su tesis inédita de maestría de 1947,[13][14] pero el nombre se cambió a Acrocanthosaurus atokensis en la publicación formal. Acracanthus es considerado un nombre de disertación, por lo tanto inválido.

El holotipo y paratipo, OMNH 10146 y OMNH 10147 respectivamente, fueron descubiertos a principios de la década de 1940 y descritos al mismo tiempo en 1950, consisten en dos individuos incompletos con restos del cráneo, encontrados en la Formación Antlers en Oklahoma.[8] Dos ejemplares mucho más completos fueron descritos en la década de 1990. El primero, SMU 74646 es un esqueleto parcial, que carece de gran parte del cráneo, que fue excavado en la Formación Twin Mountain en Texas y actualmente pertenece a la colección del Museo de Ciencia e Historia de Fort Worth.[10] Y el más completo de los esqueletos, NCSM 14345, apodado "Fran", fue hallado también en la Formación de Antlers por coleccionistas privados, el cual fue reconstruido por el Instituto de Black Hills en Dakota del Sur, y actualmente reside en el Museo de Ciencias Naturales de Carolina del Norte en Raleigh. Este es el espécimen más grande descubierto hasta ahora y posee el único cráneo y brazo completo.[4] Los elementos recobrados del paratipo son casi del mismo tamaño que los de NCSM 14345, sin embargo los del holotipo y SMU 74646 son considerablemente más pequeños.[4]
La presencia de Acrocanthosaurus en la Formación Cloverly se estableció en 2012 con la descripción de otro esqueleto parcial, UM 20796. Este ejemplar, que consta de partes de dos vértebras, huesos púbicos parciales, un fémur, peroné parcial, y fragmentos varios, representa un animal juvenil. Proviene de una cama de huesos en la cuenca Bighorn del centro-norte de Wyoming, y fue encontrado cerca del omóplato de un Sauroposeidon. Un surtido de otros fragmentyos de terópodos de la formación también pueden pertenecer a Acrocanthosaurus, que podría ser el único terópodo grande en la Formación Cloverly.[15] De Acrocanthosaurus se conocen especímenes menos completos que no proceden de Oklahoma y Texas. Un diente del sur de Arizona ha sido asignado a este género,[16] y marcas de dientes han sido halladas en huesos de un saurópodo de la misma área.[17] Varios dientes de la Formación de Arundel en Maryland han sido identificados con los del acrocantosaurio e indican una posible distribución hasta el extremo este de Norteamérica.[18] Muchos otros dientes y huesos, procedentes de varias formaciones geológicas alrededor del oeste estadounidense, se han asignado a Acrocanthosaurus, pero muchos han sido identificados erróneamente.[19] Sin embargo esto ha sido puesto en discusión después de la descripción de ejemplares en la formación Clovery.[15] En el 2022, varias huellas (icnofósil) de este terópodo fueron descubiertas en el Valle de los Dinosaurios, (Texas), como consecuencia de una intensa sequía en esta región de Estados Unidos, lo que provocó que el lecho de un río se secara y quedaran al descubierto cerca de 60 impresiones (estimándose un total de 140) en un trayecto de 30 metros, con una antigüedad de 113 millones de años, y que son consideradas cómo unas de las huellas de dinosaurio mejor conservadas del mundo.[20]
Clasificación
Acrocanthosaurus está clasificado en la superfamilia Allosauroidea, dentro del infraorden Tetanurae. Esta superfamilia se caracteriza por tener un par de crestas sobre los huesos nasales y lagrimales al tope del hocico y altas espinas neurales en las vértebras del cuello, entre otros rasgos.[7] Fue originalmente incluido en la familia Allosauridae junto con el alosaurio,[8] algo apoyado por algunos trabajos recientes.[4] Otros investigadores lo clasifican dentro de la cercana familia Carcharodontosauridae.[7][10][21][22][23]
Cuando fue descubierto, Acrocanthosaurus y muchos otros grandes terópodos se conocían solo por escasos restos, lo que llevó a este género a variar constantemente de clasificación. J. Willis Stovall y Wann Langston Jr. primero lo asignaron a "Antrodemidae", un equivalente de Allosauridae, pero fue transferido al taxón "reciclable" Megalosauridae por Alfred Sherwood Romer en 1956.[24] Para otros investigadores, la presencia de largas espinas sugieren un parentesco con Spinosaurus.[25][26] Esta interpretación del Acrocanthosaurus como un espinosáurido persistió hasta los 1980s,[27] y fue repetida en libros semi-técnicos de dinosaurios de ese entonces.[28][29]
Vértebras con largas espinas del Cretácico inferior de Inglaterra se consideraron muy parecidas a las del Acrocanthosaurus,[30] y en 1988, Gregory S. Paul las describió como una segunda especie del género, A. altispinax.[31] Estos huesos fueron originalmente asignados a Altispinax, un terópodo conocido solo a partir de dientes, y esta asignación llevó a siquiera un autor a proponer que el mismo Altispinax fue sinónimo de Acrocanthosaurus.[30] Estas vértebras fueron luego asignadas a un nuevo género, Becklespinax.[32]
Si el Acrocanthosaurus fue un alosáurido, tuvo que estar más relacionado con géneros del Jurásico como el Allosaurus y Saurophaganax.[4] Los análisis cladísticos que lo clasifican como carcarodontosáurido, comúnmente lo ubican en una posición basal al Carcharodontosaurus de África y Giganotosaurus de Sudamérica.[3][7][10] Sin embargo, el Neovenator, descubierto en Inglaterra, es usualmente considerado un carcarodontosáurido aún más basal,[7][10][11] aunque también como un neovenatórido.[33] En caso de ser un carcharodontosáurido, esto indicaría que la familia apareció en Europa y luego se dispersó a continentes sureños (en ese entonces unidos en el supercontinente Gondwana). Si el Acrocanthosaurus fue un carcarodontosáurido, entonces su distribución debió llegar incluso a Norteamérica.[10] Todos los carcarodontosáuridos existieron desde principios hasta mediados del Cretácico.[7]
Filogenia
El siguiente cladograma se basa en el estudio hecho por Novas et al., 2013, mostrando la posición de Acrocanthosaurus dentro de la familia Carcharodontosauridae.[34]
| ||||||||||||||||||||||||||||||||||||||||||||||
Nota: El análisis de Novas et al., 2013, incluyó al Neovenator como un miembro basal de Carcharodontosauridae. Pero aquí se omite a este género debido que actualmente se considera como miembro de la familia Neovenatoridae.[33]
Paleobiología
Desarrollo

De las características óseas de los ejemplares OMNH 10146 y NCSM 14345, se estima que un acrocantosaurio requería por lo menos 12 años de crecer completamente. Este número puede ser mucho mayor debido a que en el proceso de remodelación de los huesos y el crecimiento de la cavidad medular, algunas de las líneas de Harris se perdieron. Si se tienen en cuenta estas líneas, acrocantosaurio necesitaría 18-24 años para llegar a la madurez.[35]
Función de los brazos
Como otros terópodos no-avianos, los brazos del acrocantosaurio no tocaban el suelo y no se usaban para la locomoción, sirviendo en cambio para la caza. Al haberse descubierto un brazo completo (NCSM 14345), se realizó el primer estudio de la función y movimiento de los brazos del acrocantosaurio.[36] El estudio permitió conocer la amplitud de la articulación y el ángulo máximo de apertura. En muchas de las articulaciones las caras óseas no se contactaban directamente, evidenciando una gran presencia de cartílago en la uniones como en muchos arcosaurios. Otro descubrimiento es que llevaba los brazos en posición encogida contra el cuerpo, con los brazos colgando de los hombros con el húmero plegado levemente hacia medial, el codo doblado, y las garras mirando hacia adentro.[36]

El hombro del acrocantosaurio poseía un rango de movimiento menor que el de los humanos. Los brazos no podían hacer un círculo completo pero podía retraerlo 109° grados de la línea vertical. La protracción estaba limitada solo a 24° de la vertical. Y una abducción de alrededor del 9° en horizontal. El radio y la ulna estaban unidos no permitiendo la pronación o supinación.[36] El movimiento del codo también fue limitado comparado a los seres humanos, con una amplitud total de movimiento de solamente 57°. El brazo no podía extenderse totalmente, ni podría flexionarse muy lejos, con el húmero incapaz incluso de formar un ángulo recto con el antebrazo.[36]
Ningunos de los carpales fueron encontrados juntos, sugiriendo un montón de cartílago en la muñeca. Cuando todos los dedos estaban hiperextendidos casi llegaban a tocar la muñeca y cuando se flexionaban convergían sobre el primer dedo. El primer dedo poseía una garra más grande, permanecía flexionado casi contra la palma, el segundo la flexión era menor y el tercero podía estar extendido o flexionado.[36]
Antes del estudio de los miembros delanteros se suponía que eran usados para la caza, pero estos no podían abrirse lo suficiente como para atrapar el cuello de su presa. Es probable que no usaran los miembros superiores al comienzo del ataque, por lo que un acrocantosaurio debía iniciar el ataque con la boca. Cuando ya había atrapado a su presa, los brazos lo aferraban fuertemente contra el cuerpo impidiendo que la presa se escapase. Mientras que la presa luchaba por escapar, se habrían clavado más profundamente las garras permanentemente dobladas de los primeros dos dígitos. El hiperextensibilidad extremo de los dedos pudo haber sido una adaptación permitiendo al acrocantosaurio sostener la presa durante la lucha sin el miedo de una dislocación. Una vez que la presa fue atrapada contra el cuerpo, el acocantosaurio podría haberla matado a dentelladas. Otra posibilidad es que el acrocantosaurio sostuviese su presa con la boca, mientras que en varias ocasiones rasgaba grandes incisiones con sus garras de las manos.[36]
Cerebro y estructura interna del oído
En 2005, científicos reconstruyeron una réplica de la bóveda craneal de un acrocantosaurio usando tomografía computarizada para analizar los espacios intracraneales del holotipo (OMNH 10146). En vida, gran parte de este espacio estaba ocupado por el cerebro, las meninges y el fluido cerebroespinal. La mayoría de los rasgos del cerebro y los nervios craneales se pueden determinar de una réplica endocraneal y compararla a las de otros terópodos. Así se demostró que el cerebro es parecido al de muchos otros terópodos, pero guarda más semejanza con el de los alosauroides tales como el carcarodontosaurio y el giganotosaurio, y no con los del alosaurio y del sinraptor, siendo soporte para la hipótesis de que el acrocantosaurio fue un carcarodontosáurido.[37]

El cerebro tenía forma de banana, sin mucha expansión en sus hemisferios, más parecido al de un cocodrilo que al de un pájaro, lo que sostiene una idea sobre el cerebro de los téropodos no-coelurosaurianos. El acrocantosaurio poseía grandes bulbos olfativos que indican un buen sentido del olfato. Al reconstruirse los canales semicirculares del oído, los cuales controlan el equilibrio, demuestran que la cabeza se mantenía a un ángulo horizontal debajo de los 25°. Esto se determinó al orientar la réplica endocraneal para que el canal semicircular lateral sea paralelo al suelo, en la postura común de un animal alertado.[37]
Posibles icnitas

La Formación de Glen Rose del centro de Texas preserva muchas huellas fósiles (icnitas), incluyendo marcas de tres dedos de terópodos. La más famosa serie de icnitas fue hallada en el lecho del río Paluxy en el Parque nacional del Valle del Dinosaurio, siendo hoy exhibida en el Museo Americano de Historia Natural en Nueva York,[38] pero varios otros sitios con huellas han sido descritos.[39][40] Es imposible asegurar a que animal pertenece estas huellas. Pero se considera que fueron echas por acrocantosaurio.[41]

En 2001 un estudio comparativo de las huellas de Glen Rose no se han podido asigna a ningún terópodo en particular. Pero el tamaño del animal debió ser cercano al del acrocantosaurio. Y debido a la cercanía de Glen Rose a la formaciones de Antlers y Twin Mountain donde el único gran terópodo es el acrocantosaurio, es el más posible autor de las huellas.[42]

La famosa vía de Glen Rose mostrada en Nueva York incluye huellas de varios terópodos moviéndose en la misma dirección que doce saurópodos. La huellas de los terópodos se encuentran sobre la de los saurópodos. Esta podría ser la mejor evidencia de caza cooperativa entre los acrocantosaurios ya que estarían acechando a los saurópodos.[38] Aunque es posible también hay otras probables explicaciones, como la de terópodos moviéndose en forma independientes, en distinto momento de tiempo, que las huellas se hayan hecho con varios días de diferencia.[43] En un punto donde las huellas se cruzan, uno de los rastros de un terópodo, desaparece, lo que por muchos es evidencia de un ataque.[44] Otros científicos no apoyan la teoría ya que el saurópodo no cambia de marcha como sería de esperar luego de un ataque.[43]
Otro caso fue reportado en Arkansas, el 2018, en la formación De Queen. Se halló un largo patrón de huellas de un gran depredador, posiblemente de Acrocanthosaurus, o de otro depredador no identificado, estas huellas son datadas del Albiense. Fueron encontradas en unos lodazales de la formación De Queen, algunas huellas que permanecen en el lodazal, tienen 0.5 m de espesor, en el patrón de huellas tienen ángulos con las huellas que van en línea recta, algunas huellas de los morfotipos miden de 44 a 68 cm de largo y 33 a 56 cm de ancho, otros morfotipos miden más de 3.50 cm de ancho y largo. También, se descubrieron huellas de saurópodos, no se pudo determinar el tipo de saurópodo, al igual que las huellas del terópodo, estas tienen ángulos, un poco más de 90 grados, además, se puede ver el arrastre del dedo con el pie, estas fueron comparadas con las huellas del elefante africano (Loxodonta africana), la huella del mamífero, es similar a unas huellas del saurópodo. Las huellas de estas ambas especies, se compararon con las del saurópodo y terópodo de la formación Glen Rose, la conclusión final, fue que los saurópodos y terópodos de ambas formaciones, eran atribuibles a ser el mismo género de icnitas, un género indeterminado de huellas, pero no se sabe si eran las mismas especies de dinosaurios, o al menos, especies distintas pero del mismo género de icnitas. El tema de que estas especies vivieran juntas (saurópodo y terópodo), sería "depredador y presa" en muchos sentidos.[45]
Patología
El cráneo del holotipo de Acrocanthosaurus atokensis muestra una ligera exostosis en el material de la escamoso. La espina neural de la undécima vértebra estuvo fracturada y terminó sanando, mientras que la espina neural de la tercera vértebra de su cola tenía una inusual estructura en forma de gancho.[46]
Paleoecología

Acrocanthosaurus ha sido hallado en los estratos Albiense y Aptiense, hace 120 millones de años en la Formación Twin Mountain del norte de Texas, la Formación de Antlers del sudeste de Oklahoma. Estas formaciones no han sido datadas por radiometría, pero la bioestratigrafía fue usada para datar su edad. Con base en los cambios en los taxones de amonitas, el límite entre el Aptieno y Albiano del Cretácico Inferior se ha situado dentro de la Formación Glen Rose de Texas, que puede contener huellas Acrocanthosaurus y se encuentra justo por encima de la Formación Twin Mountain. Esto indica que la Formación Twin Mountain se encuentra en su totalidad dentro del etapa Aptiano, que duró desde 125 hasta 112 millones de años. También las pisadas de la cañada Rose pertenecen a esta época[47] La Formación Antlers contiene fósiles de Deinonychus y Tenontosaurus, que también se han encontrado en la Formación Cloverly de Montana, que ha sido radiométricamente fechada entre el Aptiano y Albiano, lo que sugiere una edad similar para la Formación Antlers.[48][7] Por lo tanto Acrocanthosaurus lo más probable es que haya existido entre 125 y 100 millones de años atrás.[7] Un diente del sudeste de Arizona ha sido referido a este género,[16] y además hay marcas de dientes en huesos de saurópodos de la misma área.[17] Varios dientes de Maryland han sido descritos como pertenecientes a Acrocanthosaurus y representan su registro más hacia al este.
Durante esta época, el área donde fue hallado fue una zona pantanosa que drenaba hacia el mar. Millones de años después el mar invadiría la zona creando el Mar de Niobrara que dividió Norteamérica en dos hasta finales del Cretácico La Formación Glen Rose representa un ambiente de costa donde el acrocantosaurio dejó sus huellas en las marismas de la antigua línea de maras. Acrocanthosaurus fue el mayor depredador de su zona, y es de esperar que necesitara un gran territorio y que estos incluyan distintos terrenos de caza.[42] Las presas potenciales son saurópodos como Paluxysaurus[49] o posiblemente el enorme Sauroposeidon,[50] así como grandes ornitópodos como Tenontosaurus.[51] El pequeño terópodo Deinonychus que solo alcanzaba 3 metros de largo era poca competencia para el gigante Acrocanthosaurus.[48]
Referencias
- Holtz, Thomas (2012). Genus List for Holtz (2007) Dinosaurs:The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. Consultado el 17 de octubre de 2015.
- Bates, K.T., Manning, P.L., Hodgetts, D. and Sellers, W.I. (2009). «Estimating Mass Properties of Dinosaurs Using Laser Imaging and 3D Computer Modelling». En Beckett, Ronald, ed. PLoS ONE 4 (2): e4532. Bibcode:2009PLoSO...4.4532B. PMC 2639725. PMID 19225569. doi:10.1371/journal.pone.0004532. «We therefore suggest 5750–7250 kg represents a plausible maximum body mass range for this specimen of Acrocanthosaurus. »
- Eddy, Drew R.; Clarke, Julia A. (2011). «New Information on the Cranial Anatomy of Acrocanthosaurus atokensis and Its Implications for the Phylogeny of Allosauroidea (Dinosauria: Theropoda)». En Farke, Andrew, ed. PLoS ONE 6 (3): e17932. Bibcode:2011PLoSO...6E7932E. doi:10.1371/journal.pone.0017932.
- Currie, Philip J.; Carpenter, Kenneth. (2000). «A new specimen of Acrocanthosaurus atokensis (Theropoda, Dinosauria) from the Lower Cretaceous Antlers Formation (Lower Cretaceous, Aptian) of Oklahoma, USA». Geodiversitas 22 (2): 207-246.
- Campione, Nicolas; Evans, David; Brown, Caleb; Carrano, Matthew (21 de julio de 2014). «Body mass estimation in non-avian bipeds using a theoretical conversion to quadruped stylopodial proportions». Methods in Ecology and Evolution 5 (9): 913-923. doi:10.1111/2041-210X.12226. Consultado el 29 de octubre de 2015.
- Therrien, F.; Henderson, D.M. (2007). «My theropod is bigger than yours...or not: estimating body size from skull length in theropods». Journal of Vertebrate Paleontology 27 (1): 108-115. doi:10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2.
- Holtz, Thomas R.; Molnar, Ralph E.; & Currie, Philip J. (2004). «Basal Tetanurae». En Weishampel, David B.; Dodson, Peter; & Osmólska, Halska (eds.), ed. The Dinosauria (Second Edition edición). Berkeley: University of California Press. pp. 71-110. ISBN 0-520-24209-2.
- Stovall, J. Willis; Langston, Wann. (1950). «Acrocanthosaurus atokensis, a new genus and species of Lower Cretaceous Theropoda from Oklahoma». American Midland Naturalist 43: 696-728.
- Molnar, Ralph E.; Kurzanov, Sergei M.; & Dong Zhiming. (1990). «Carnosauria». En Weishampel, David B.; Dodson, Peter; & Osmólska, Halska (eds.), ed. The Dinosauria (First Edition edición). Berkeley: University of California Press. pp. 169-209. ISBN 0-520-06727-4.
- Harris, Jerald D. (1998). «A reanalysis of Acrocanthosaurus atokensis, its phylogenetic status, and paleobiological implications, based on a new specimen from Texas». New México Museum of Natural History and Science Bulletin 13: 1-75.
- Naish, Darren; Hutt, Stephen; & Martill, David M. (2001). «Saurischian Dinosaurs 2: Theropods». Dinosaurs of the Isle of Wight. London: The Palaeontological Association. pp. 242-309. ISBN 978-0901702722.
- Liddell, Henry George; Robert Scott (1980). Greek-English Lexicon, Abridged Edition. Oxford: Oxford University Press. ISBN 0-19-910207-4.
- Langston, Wann R. (1947). A new genus and species of Cretaceous theropod dinosaur from the Trinity of Atoka County, Oklahoma. Unpublished M.S. thesis. University of Oklahoma.
- Czaplewski, Nicholas J.; Cifelli, Richard L.; Langston, Wann R., Jr. (1994). «Catalog of type and figured fossil vertebrates. Oklahoma Museum of Natural History». Oklahoma Geological Survey Special Publication 94 (1): 1-35.
- D'Emic, Michael D.; Melstrom, Keegan M.; Eddy, Drew R. (2012). «Paleobiology and geographic range of the large-bodied Cretaceous theropod dinosaur Acrocanthosaurus atokensis». Palaeogeography, Palaeoclimatology, Palaeoecology. 333–334: 13-23. doi:10.1016/j.palaeo.2012.03.003.
- Ratkevich, Ronald P. (1997). «Dinosaur remains of southern Arizona». En Wolberg, Donald L.; Stump, Edward; & Rosenberg, Gary, ed. Dinofest International: Proceedings of a Symposium Held at Arizona State University. Philadelphia: Academy of Natural Sciences. ISBN 978-0935868944.
- Ratkevich, Ronald P. (1998). «New Cretaceous brachiosaurid dinosaur, Sonorasaurus thompsoni gen. et sp. nov., from southern Arizona». Journal of the Arizona-Nevada Academy of Science 31 (1): 71-82. Archivado desde el original el 22 de agosto de 2007.
- Lipka, Thomas R. (1998). «The affinities of the enigmatic theropods of the Arundel Clay facies (Aptian), Potomac Formation, Atlantic Coastal Plain of Maryland». En Lucas, Spencer G.; Kirkland, James I.; & Estep, J.W. (eds.), ed. Lower and Middle Cretaceous Terrestrial Ecosystems. New México Museum of Natural History and Science Bulletin 14. pp. 229-234.
- Harris, Jerald D. (1998). «Large, Early Cretaceous theropods in North America». En Lucas, Spencer G.; Kirkland, James I.; & Estep, J.W. (eds.), ed. Lower and Middle Cretaceous Terrestrial Ecosystems. New México Museum of Natural History and Science Bulletin 14. pp. 225-228.
- «El asombroso hallazgo de las huellas de un dinosaurio que quedaron expuestas por la sequía en Estados Unidos». BBC News Mundo. Consultado el 25 de agosto de 2022.
- Sereno, Paul C.; Dutheil, Didier B.; Iarochene, M.; Larsson, Hans C.E.; Lyon, Gabrielle H.; Magwene, Paul M.; Sidor, Christian A.; Varricchio, David J. et al. (1996). «Predatory dinosaurs from the Sahara and Late Cretaceous faunal differentiation». Science 272 (5264): 986-991. doi:10.1126/science.272.5264.986.
- Brusatte, Stephen L.; Benson, Roger B. J.; Chure, Daniel J.; Xu, Xing; Sullivan, Corwin; Hone, David W. E. (2009). «The first definitive carcharodontosaurid (Dinosauria: Theropoda) from Asia and the delayed ascent of tyrannosaurids». Naturwissenschaften 96 (9): 1051-8. Bibcode:2009NW.....96.1051B. PMID 19488730. doi:10.1007/s00114-009-0565-2.
- Benson, Roger B. J.; Carrano, Matthew T.; Brusatte, Stephen L. (2009). «A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic». Naturwissenschaften 97 (1): 71-8. Bibcode:2010NW.....97...71B. PMID 19826771. doi:10.1007/s00114-009-0614-x.
- Romer, Alfred S. (1956). Osteology of the Reptiles. Chicago: University of Chicago Press. pp. 772pp. ISBN 0-89464985-X.
- Walker, Alick D. (1964). «Triassic reptiles from the Elgin area: Ornithosuchus and the origin of carnosaurs». Philosophical Transactions of the Royal Society of London, Series B, Biological Sciences 248: 53-134.
- Romer, Alfred S. (1966). Vertebrate Paleontology (Third Edition edición). Chicago: University of Chicago Press. pp. 468pp. ISBN 0-7167-1822-7.
- Carroll, Robert L. (1988). Vertebrate Paleontology and Evolution. W.H. Freeman and Company. ISBN 0716718227.
- Lambert, David; and the Diagram Group (1983). «Spinosaurids». A Field Guide to Dinosaurs. Nueva York: Avon Books. pp. 84-85. ISBN 0-380-83519-3.
- Norman, David B. (1985). «Carnosaurs». The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom. Nueva York: Crescent Books. pp. 62-67. ISBN 0-517-468905.
- Glut, Donald F. (1982). The New Dinosaur Dictionary. Secaucus, NJ: Citadel Press. pp. 39, 48. ISBN 0-8065-0782-9.
- Paul, Gregory S. (1988). «Genus Acrocanthosaurus». Predatory Dinosaurs of the World. Nueva York: Simon & Schuster. pp. 314–315. ISBN 0-671-61946-2.
- Olshevsky, George (1991). A Revision of the Parainfraclass Archosauria Cope, 1869, Excluding the Advanced Crocodylia. San Diego: Publications Requiring Research. pp. 196pp.
- Benson, Roger; Carrano, Matthew; Brusatte, Stephen (2010). «A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic». Naturwissenschaften 97 (1): 71-78. doi:10.1007/s00114-009-0614-x. Consultado el 29 de octubre de 2015.
- Novas, Fernando; Agnolín, Federico; Ezcurra, Martín; Porfiri, Juan; Canale, Juan (2013). «Evolution of the carnivorous dinosaurs during the Cretaceous: The evidence from Patagonia». Cretaceous Research (elSevier) 45: 174-215. doi:10.1016/j.cretres.2013.04.001. Consultado el 29 de octubre de 2015.
- D'Emic, Michael; Melstrom, Keegan; Eddy, Drew (15 de mayo de 2012). «Paleobiology and geographic range of the large-bodied Cretaceous theropod dinosaur Acrocanthosaurus atokensis». Palaeogeography, Palaeoclimatology, Palaeoecology. 333-334: 13-23. doi:10.1016/j.palaeo.2012.03.003. Consultado el 2 de agosto de 2015.
- Senter, Phil; Robins, James H. (2005). «Range of motion in the forelimb of the theropod dinosaur Acrocanthosaurus atokensis, and implications for predatory behaviour». Journal of Zoology 266 (3): 307-318. doi:10.1017/S0952836905006989.
- Franzosa, Jonathan; Rowe, Timothy. (2005). «Cranial endocast of the Cretaceous theropod dinosaur Acrocanthosaurus atokensis». Journal of Vertebrate Paleontology 25 (4 páginas=859–864).
- Bird, Roland T. (1941). «A dinosaur walks into the museum». Natural History 43: 254-261.
- Rogers, Jack V. (2002). «Theropod dinosaur trackways in the Lower Cretaceous (Albian) Glen Rose Formation, Kinney County, Texas». Texas Journal of Science 54 (2): 133-142.
- Hawthorne, J. Michael; Bonem, Rena M.; Farlow, James O.; & Jones, James O. (2002). Ichnology, stratigraphy and paleoenvironment of the Boerne Lake Spillway dinosaur tracksite, south-central Texas revista=Texas Journal of Science 54 (4). pp. 309-324.
- Langston, Wann (1974). «Non-mammalian Comanchean tetrapods». Geoscience and Man 3: 77-102.
- Farlow, James O.. «Acrocanthosaurus and the maker of Comanchean large-theropod footprints». En Tanke, Darren; & Carpenter, Ken. (eds.), ed. Mesozoic Vertebrate Life. Bloomington: Indiana University Press. pp. 408-427. ISBN 978-0253339072.
- Lockley, Martin G. (1991). Tracking Dinosaurs: A New Look at an Ancient World. Cambridge: Cambridge University Press. pp. 252pp. ISBN 978-0521394635.
- Thomas, David A.; & Farlow, James O. (1997). «Tracking a dinosaur attack». Scientific American 266 (6): 48-53.
- Brian F. Platt; Celina A. Suarez; Stephen K. Boss; Malcolm Williamson; Jackson Cothren; Jo Ann C. Kvamme (2018). "LIDAR-based characterization and conservation of the first theropod dinosaur trackways from Arkansas, USA". PLoS ONE. 13 (1): e0190527. doi:10.1371/journal.pone.0190527.
- Molnar, R. E., 2001, Theropod paleopathology: a literature survey: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 337-363.
- Jacobs, Louis L.; Winkler, Dale A.; & Murry, Patrick A. (1991). «On the age and correlation of Trinity mammals, Early Cretaceous of Texas, USA». Newsletter of Stratigraphy 24: 35-43.
- Brinkman, Daniel L.; Cifelli, Richard L.; & Czaplewski, Nicholas J. (1998). «First occurrence of Deinonychus antirrhopus (Dinosauria: Theropoda) from the Antlers Formation (Lower Cretaceous: Aptian - Albian) of Oklahoma». Oklahoma Geological Survey Bulletin 146: 1-27.
- Rose, Peter J. (2007). «A new titanosauriform sauropod (Dinosauria: Saurischia) from the Early Cretaceous of central Texas and its phylogenetic relationships». Palaeontologia Electronica 10 (2): 65pp. [published online]
- Wedel, Matthew J. (2000). «Sauroposeidon proteles, a new sauropod from the Early Cretaceous of Oklahoma». Journal of Vertebrate Paleontology 20 (1): 109-114. (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- Winkler, Dale A.; Murry, Patrick A.; & Jacobs, Louis L. «A new species of Tenontosaurus (Dinosauria: Ornithopoda) from the Early Cretaceous of Texas». Journal of Vertebrate Paleontology 17 (2): 330-348. Archivado desde el original el 27 de septiembre de 2007.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikispecies tiene un artículo sobre Acrocanthosaurus atokensis.
Wikispecies tiene un artículo sobre Acrocanthosaurus atokensis. Wikimedia Commons alberga una galería multimedia sobre Acrocanthosaurus atokensis.
Wikimedia Commons alberga una galería multimedia sobre Acrocanthosaurus atokensis.- "Fran", el Acrocanthosaurus atokensis en el Museo Mundial de Historia Natural (en inglés)
- Acrocanthosaurus FAQ del Museo de Ciencias Naturales de Carolina del Norte (en inglés)
- Acrocanthosaurus en la Dinosaur Encyclopaedia de Dino Russ's Lair (en inglés)
- Acrocanthosaurus en Dinodata (en inglés, se requiere registrarse en forma gratuita)
- Acrocanthosaurus dentro de Carnosauria en The Thescelsosaurus (en inglés)
- Ficha e imagen de Acrocanthosaurus en Dinosaurier-web (en alemán)
- Significado y pronunciación del nombre (en inglés)
- Acrocanthosaurus en the Theropod Database (en inglés)
| Control de autoridades |
|
|---|
 Datos: Q131021
Datos: Q131021- Multimedia: Acrocanthosaurus / Q131021
- Especies: Acrocanthosaurus