Mapusaurus roseae
Mapusaurus roseae la única especie conocida del género extinto Mapusaurus (arn. "lagarto de la tierra") de dinosaurio terópodo carcarodontosáurido que vivieron a principios del período Cretácico, entre hace 97 y 93,5 millones de años, durante el Cenomaniense,[1] en lo que hoy es Suramérica.
| Mapusaurus roseae | ||
|---|---|---|
| Rango temporal: 97 Ma - 93,5 Ma Cretácico Superior | ||
 Restauración de Mapusaurus roseae | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| Infraorden: | Carnosauria | |
| Superfamilia: | Allosauroidea | |
| Familia: | Carcharodontosauridae | |
| Subfamilia: | Carcharodontosaurinae | |
| Tribu: | Giganotosaurini | |
| Género: |
Mapusaurus Coria y Currie, 2006 | |
| Especie: |
M. roseae Coria y Currie, 2006 | |
Descripción
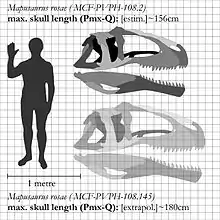
Basándose en las proporciones del espécimen más grande encontrado, se ha estimado que Mapusaurus roseae medía de 12,2 a 12,6 metros de longitud y que pesaba hasta 6,8 toneladas.[1][2][3] Dimensiones que resultan bastante grandes hasta para un terópodo, pero aun así, es de menor tamaño que Giganotosaurus. Los restos de Mapusaurus fueron exhumados entre 1997 y 2001 por la expedición argentino-canadiense Proyecto Dinosaurio, de una zona expuesta de la Formación de Huincul, parte del grupo de Río Limay, del Cenomaniense, en la provincia argentina de Neuquén, en un lecho que contenía por lo menos siete individuos de varias etapas ontogenéticas. Los responsables de la extracción fueron el profesor paleontólogo argentino Rodolfo Coria y su colega canadiense Philip Currie, quienes lo bautizaron en honor a la etnia mapuche que habita la región e hicieron público el descubrimiento en 2006.[2]

Mapusaurus era un gran terópodo y era más o menos similar en tamaño a su pariente cercano Giganotosaurus, con los individuos más grandes conocidos de aproximadamente 10,2 metros de longitud o más y un peso de aproximadamente 3 toneladas. El individuo más largo para el cual Coria y Currie en 2006 proporcionaron una estimación concreta en la Tabla 1, apéndice III, es el animal al que pertenecía el fémur MCF-PVPH-208.203. Este individuo se estima en 10,2 metros de largo.[2]
Coria y Currie notan la presencia de huesos aislados de al menos un individuo más largo, pero no proporcionan una figura, sino que encuentran los huesos más grandes coherentes con un individuo de tamaño comparable al holotipo de Giganotosaurus estimado en 12,7 metros de longitud, aunque no con las mismas proporciones exactas, con espinas neurales más altas y anchas, un peroné más alargado, 86 centímetros en comparación con 83.5 centímetros, pero más delgado, 81-89% del ancho que en Giganotosaurus, así como un eje púbico más ancho en dimensiones mínimas, 10% más ancho como lo indica un fragmento largo de 7,8 centímetros, catalogado como MCF-PVPH-108.145 y con un cráneo de proporciones diferentes, más corto en longitud que Giganotosaurus porque el maxilar no es alargado, 12 dientes en comparación con 14 en Carcharodontosaurus, pero es más profundo en proporción debido a esto, así como más estrecho, debido a los estrechos nasales. Considerando esto, un maxilar fragmentario es coherente con el tamaño del individuo del tamaño de Giganotosaurus, MCF-PVPH-108.169'. Un arco neural desde un eje, MCF-PVPH-108.83 y un fragmento de cuchilla escapular también tienen el mismo tamaño exacto que los elementos en Giganotosaurus . El peso estimado de 6,800 kilogramos es de un fémur de 1300 milímetros de largo con una circunferencia de 455 milímetros, de MCF-PVPH-208.234.[2]
Holtz estimó el tamaño máximo del animal en 12,6 metros.[4] Esta estimación ha sido citada en Drew Eddy y Julia Clarke en 2011,[5] y nuevamente citada en una tabla filogenética en un análisis de 2014 por Canale et al.[6] Otros autores sugirieron que medía 12.5 metros de largo y 7.3 toneladas de peso.[7]
Coria y Currie han especulado que el nicho del hallazgo puede representar una acumulación de restos de largo plazo (posiblemente debido a una cierta clase de trampa natural), y podría proporcionar pistas sobre el comportamiento de Mapusaurus. Se han encontrado otros lechos de terópodos con similares características, como la denominada mina del Dinosaurio de Cleveland Lloyd en Utah con Allosaurus, los yacimientos de Albertosaurus en Alberta (Canadá) y de Daspletosaurus, en Montana, Estados Unidos.[2]
Coria, titular del Museo municipal Carmen Funes, anunció en una conferencia de prensa realizada a comienzos del 2006, la posibilidad de que la alta concentración de huesos fósiles pudiera indicar que Mapusaurus habría cazado en grupo, con algún esquema de coordinación mutua, lo que le habría posibilitado dominar a inmensas criaturas como Argentinosaurus.[8] Esto contradice la hipótesis central de su último trabajo, aunque de ser plausible constituiría la primera prueba substantiva de comportamiento gregario entre terópodos grandes aparte de Tyrannosaurus, aunque el mecanismo concreto de caza permanece desconocido, ya que no queda claro si lo hacían en grupos organizados o si simplemente atacaban en banda.[2]

Coria y Currie en 2006 diagnostican Mapusaurus como un terópodo carcarodontosáurido cuyo cráneo difiere de Giganotosaurus en que tiene huesos nasales gruesos y rugosos, más angostos en la porción anterior a la unión nasal/maxilar/lagrimal, posee una mayor extensión de la fosa anteorbital hacia el maxilar y menores fenestras maxilares, una barra interfenestral de mayor grosor entre las fenestras anteorbital y maxilar y un lacrimal más bajo y aplanado con un pequeño cuerno. Transversalmente el prefrontal es más ancho en lo referente a anchura lacrimal, el margen lateral del palpebral ventrolateralmente curvando, placa interdentales baja; una posición más alta del canal de Meckel; margen anteroventral más inclinado ventrolateralmente del dentario. Mapusaurus roseae es único en que el proceso de la parte superior cuadrado-yugal del yugal se divide en dos pequeños anteriores, un foramen milohioideo sobre el contacto del dentario con el esplenial. El segundo y tercer metacarpianos fusionados, húmero con el extremo distal amplio y con poca separación en medio de los cóndilos. La brevis fossa del ilion se extiende profundamente en dorsal de la excavación del pedúnculo isquial. También se diferencia de Giganotosaurus en tener las epipófisis cónicas, levemente curvadas en las cervicales que se afilan hacia el extremo distal. Zigapófisis posteriores axiales unidas en la línea media, una lámina prespinal más pequeña y menos elaborada en el centro de las cervicales. Un notable margen dorsal agudo de las espinas dorsales de las vértebras, que son altas, más anchas. EL eje isquiático es curvado y un peroné más delgado[2]
Descubrimiento e investigación
Mapusaurus fue excavado entre 1997 y 2001, por el Proyecto dinosaurio Argentino-Canadiense, de un afloramiento de la Formación Huincul en el Subgrupo Río Limay en Cañadón El Gato. Fue descrito y nombrado por los paleontólogos Rodolfo Coria y Philip Currie en 2006.[2]

El nombre Mapusaurus deriva del mapudungun mapu, que significa "de la tierra", y del griego saurus, "lagarto". La especie hallada, Mapusaurus roseae, fue bautizada así por lleva el nombre de las rocas de color rosa, en las que se encontraron los fósiles y de Rose Letwin , que patrocinó las expediciones que recuperaron estos fósiles .
La obra de remoción de los sedimentos petrificados que cubrían los aproximadamente 300 huesos demandó cinco meses de trabajo. El lugar del hallazgo en el Cañadón del Gato se interpreta como un depósito de agua dulce, "acumulado por una corriente efímera o estacional en una región con clima árido o semiárido".[2] El lecho es especialmente interesante a la luz de la significativa escasez de material fosilizado dentro de la Formación Huincul.

El holotipo designado para el género y especie, Mapusaurus roseae, es un hueso nasal derecho aislado, MCF-PVPH-108.1, alojado en el Museo Carmen Funes, Paleontología de Vertebrados, Plaza Huincul, Neuquén. Doce paratipos han sido nombrados basados en diferentes elementos óseos inconexos. Tomados juntos, todos estos fragmentos componen la mayor parte del esqueleto completo del animal.[2] Los paratipos son, MCF-PVPH-108.5, un lagrimal izquierdo y prefrontal, MCF-PVPH-108.45, húmero derecho, MCFPVPH-108.83, axis, MCF-PVPH-108.90, arco neural cervical, MCF-PVPH-108,115, maxilar derecho, MCFPVPH-108,125, un dentario izquierdo, MCF-PVPH-108,128, un ilion izquierdo, MCF-PVPH-108,165, un isquion izquierdo, MCFPVPH-108,167, un yugal, MCF-PVPH-108,177, un postorbital derecho y palpebral, MCF-PVPH-108,179, esplenial derecho y MCF-PVPH-108.202, un peroné derecho.[2]
El 17 de abril de 2006, se dio una publicidad de prensa más amplia al hallazgo. Además, el tamaño de Mapusaurus fue exagerado, se hizo la falsa afirmación de que era el dinosaurio carnívoro más grande conocido, y se repitieron las especulaciones previas sobre la caza en grupos, aunque el análisis tafonómico de los hallazgos demostró claramente que los diversos esqueletos no fueron enterrados de una sola vez.[9]
Clasificación
Estudios cladísticos realizados por Coria y Currie demuestran definitivamente que Mapusaurus está dentro del grupo de los Carcharodontosauridae, observándose que la estructura del fémur sugiere una relación más cercana con Giganotosaurus de lo que cualquiera de los dos taxones comparte con Carcharodontosaurus. Los investigadores proponen una nueva taxonomía monofilética basada en esta relación: la subfamilia Giganotosaurinae, definida como todos los carcarodontosáuridos más cercanos a Giganotosaurus y Mapusaurus que al Carcharodontosaurus. El género Tyrannotitan se incluye en un principio tentativamente en esta nueva subfamilia, hasta que finalice la publicación de descripciones más detalladas de los especímenes recuperados de este último subtipo, pero hoy se la coloca con el dinosaurio africano.[2]
Posteriormente, en 2013 una revisión hecha por Novas et al., reordenó a varios de los géneros conocidos de la familia Carcharodontosauridae, en esta revisión se cambió a la subfamilia Giganotosaurinae por la Carcharodontosaurinae. Esta nueva subfamilia, solo están incluidos los carcarodontosáuridos que habitaron en Suramérica y África y que vivieron entre el Aptiano y el Cenomaniano. Por lo tanto, Carcharodontosaurus, Tyrannotitan, Mapusaurus y Giganotosaurus son los únicos géneros incluidos en esta subfamilia. Y los tres últimos mencionados, los cuales corresponden solo a taxones patagónicos, fueron reasignados en una nueva tribu llamada Giganotosaurini. Estos tres géneros comparten entre sí varias características, como un proceso postorbital del yugal con una base amplia en sentido anteroposterior (desde el frente hasta la parte de atrás), no tienen pleurocelos en las vértebras caudales, y varias características del fémur como un cuarto trocánter reducido o una cresta cruzada mal definida en la zona distal también los asemejan.[10]
Filogenia
El siguiente cladograma se basa en el estudio hecho por Novas et al., 2013, mostrando la posición de Mapusaurus dentro de la familia Carcharodontosauridae.

| ||||||||||||||||||||||||||||||||||||||||||||||
Nota: El análisis de Novas et al,. incluía al Neovenator como un miembro basal de Carcharodontosauridae, pero aquí se omite a este género debido que actualmente se considera como miembro de la familia Neovenatoridae.[11]
Paleobiología
Los restos fósiles de Mapusaurus fueron descubiertos en un lecho óseo que contenía al menos siete individuos de diversas etapas de crecimiento.[6][5] Coria y Currie especularon que esto podría representar una acumulación de cadáveres a largo plazo, posiblemente por coincidencia, algún tipo de trampa de depredador y puede proporcionar pistas sobre el comportamiento de Mapusaurus.[2] Otros lechos óseos de terópodos conocidos incluyen la Cantera de Dinosaurios Cleveland-Lloyd dominada por Allosaurus de Utah, un lecho óseo Albertosaurus de Alberta y un lecho óseo Daspletosaurus de Montana.
El paleontólogo Rodolfo Coria, del Museo Carmen Funes, contrario a su artículo publicado, repitió en una conferencia de prensa sugerencias anteriores de que esta congregación de huesos fósiles podría indicar que Mapusaurus cazaba en grupos y trabajaba en conjunto para derribar presas grandes, como el inmenso saurópodo Argentinosaurus.[8] Si es así, esta sería la primera evidencia sustancial de comportamiento gregario por parte de grandes terópodos que no sean Tyrannosaurus, aunque se desconoce si podrían haber cazado en manadas organizadas, como lo hacen los lobos, o simplemente haber atacado en una horda. Los autores interpretaron el ambiente deposicional de la Formación Huincul en la localidad de Cañadón del Gato como un depósito de paleocanal de agua dulce, "establecido por una corriente efímera o estacional en una región con clima árido o semiárido".[2] Este lecho óseo es especialmente interesante, a la luz de la escasez general de hueso fosilizado dentro de la Formación Huincul. Un estudio ontogenético de Canale et al en 2014 encontró que Mapusaurus mostró heterocronía, una condición evolutiva en la cual los animales pueden retener una característica ancestral durante una etapa de su vida, pero la pierden a medida que se desarrollan. En Mapusaurus , las fenestras maxilares están presentes en individuos más jóvenes, pero desaparecen gradualmente a medida que maduran.[6]
Paleoecología
Como se mencionó anteriormente, se cree que la Formación Huincul representa un ambiente árido con corrientes efímeras o estacionales. La edad de esta formación se estima en 97 a 93,5 millones de años.[4] El registro de dinosaurio se considera escaso aquí. Mapusaurus compartió su entorno con los saurópodos Argentinosaurus, uno de los saurópodos más grandes, si no el más grande y Cathartesaura . Los terópodos abelisauroides Skorpiovenator e Ilokelesia también vivían en la región.[12]
Referencias
- Holtz, Thomas (2012). Genus List for Holtz (2007) Dinosaurs:The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. Consultado el 28 de septiembre de 2015.
- Coria, Rodolfo; Currie, Philip (2006). «A new carcharodontosaurid (Dinosauria, Theropoda) from the Upper Cretaceous of Argentina.». Geodiversitas 28 (1): 71-118. Consultado el 17 de octubre de 2015.
- Campione, Nicolas; Evans, David; Brown, Caleb; Carrano, Matthew (21 de julio de 2014). «Body mass estimation in non-avian bipeds using a theoretical conversion to quadruped stylopodial proportions». Methods in Ecology and Evolution 5 (9): 913-923. doi:10.1111/2041-210X.12226. Consultado el 12 de septiembre de 2015.
- Holtz, Thomas R., Jr. (2012). Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. Genus List.
- Eddy, Drew R.; Clarke, Julia A. (21 de marzo de 2011). «New Information on the Cranial Anatomy of Acrocanthosaurus atokensis and Its Implications for the Phylogeny of Allosauroidea (Dinosauria: Theropoda)». PLOS ONE 6 (3): e17932. Bibcode:2011PLoSO...617932E. ISSN 1932-6203. PMC 3061882. PMID 21445312. doi:10.1371/journal.pone.0017932.
- Canale, Juan Ignacio; Novas, Fernando Emilio; Salgado, Leonardo; Coria, Rodolfo Aníbal (1 de diciembre de 2015). «Cranial ontogenetic variation in Mapusaurus roseae (Dinosauria: Theropoda) and the probable role of heterochrony in carcharodontosaurid evolution». Paläontologische Zeitschrift (en inglés) 89 (4): 983-993. ISSN 0031-0220. doi:10.1007/s12542-014-0251-3.
- Molina-Pérez & Larramendi 2016. Récords y curiosidades de los dinosaurios Terópodos y otros dinosauromorfos, Larousse. Barcelona, Spain p. 262
- Associated Press (2006). Details Revealed About Huge Dinosaurs. ABC News US.
- https://www.pagina12.com.ar/diario/sociedad/3-65727-2006-04-18.html
- Novas, Fernando; Agnolín, Federico; Ezcurra, Martín; Porfiri, Juan; Canale, Juan (2013). «Evolution of the carnivorous dinosaurs during the Cretaceous: The evidence from Patagonia». Cretaceous Research (elSevier) 45: 174-215. doi:10.1016/j.cretres.2013.04.001. Consultado el 13 de septiembre de 2015.
- Benson, Roger; Carrano, Matthew; Brusatte, Stephen (2010). «A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic». Naturwissenschaften 97 (1): 71-78. doi:10.1007/s00114-009-0614-x. Consultado el 13 de septiembre de 2015.
- Sánchez, Maria Lidia; Heredia, Susana; Calvo, Jorge O. (2006). «Paleoambientes sedimentarios del Cretácico Superior de la Formación Plottier (Grupo Neuquén), Departamento Confluencia, Neuquén» [Sedimentary paleoenvironments in the Upper Cretaceous Plottier Formation (Neuquen Group), Confluencia, Neuquén]. Revista de la Asociación Geológica Argentina 61 (1): 3-18 – via ResearchGate.
Enlaces externos
 Wikispecies tiene un artículo sobre Mapusaurus roseae.
Wikispecies tiene un artículo sobre Mapusaurus roseae. Wikimedia Commons alberga una categoría multimedia sobre Mapusaurus roseae.
Wikimedia Commons alberga una categoría multimedia sobre Mapusaurus roseae.
| Control de autoridades |
|
|---|
 Datos: Q131176
Datos: Q131176- Multimedia: Mapusaurus / Q131176
- Especies: Mapusaurus