Sistema inmunitario innato
La inmunidad innata comprende las células y los mecanismos que defienden al ser vivo de la infección por otros organismos de forma no específica. Esto significa que las células de la respuesta innata reconocen, y responden a patógenos de forma genérica y, a diferencia de la respuesta inmunitaria adaptativa, no confiere inmunidad a largo plazo o protectora al huésped.[1] El sistema inmunitario innato proporciona defensa inmediata contra la infección, y se encuentra tanto en animales como en vegetales.

Funciones
Se cree que los elementos de la inmunidad innata constituyen una antigua estrategia evolutiva de defensa, que predomina en las plantas, los hongos, los insectos y los microorganismos multicelulares primitivos (ver Otras formas de inmunidad innata).[2]
Las funciones principales de la respuesta inmunitaria innata en vertebrados incluyen:
- Reclutamiento de células inmunes hacia los sitios de infección y de inflamación, mediante la producción de factores químicos, incluyendo los mediadores químicos especializados, denominados citoquinas.
- Activación de la cascada del sistema del complemento para identificar bacterias, activar las células y promover el aclaramiento de las células muertas o de los complejos de anticuerpos.
- La identificación y remoción de sustancias extrañas presentes en órganos, tejidos, sangre y linfa, a cargo de los leucocitos.
- La activación de la respuesta inmunitario adaptativa mediante un proceso conocido como la presentación de antígenos.
Inflamación
La inflamación es una de las primeras respuestas del sistema inmunitario a la infección o la irritación; la inflamación es estimulada por la liberación de factores químicos y sirve para establecer una barrera física contra la propagación de la infección, y para promover la recuperación de algún tejido dañado siguiendo el aclaramiento de los patógenos.[3]
Los factores químicos producidos durante la inflamación (histamina, bradicinina, serotonina, leucotrienos) sensibilizan los receptores de dolor, causan vasodilatación de los vasos sanguíneos en la escena, y atraen fagocitos, especialmente neutrófilos.[3] Luego, los neutrófilos disparan otras partes del sistema inmunitario mediante la liberación de los factores quimiotácticos para otros leucocitos y linfocitos. La respuesta inflamatoria está caracterizada por el siguiente quinteto de síntomas: enrojecimiento, calor, hinchazón, dolor y posible disfunción de los órganos o tejidos involucrados (functio laesa).
Sistema del Complemento
El sistema del complemento es una cascada bioquímica del sistema inmunitario que ayuda, o “complementa”, la habilidad de los anticuerpos de depurar patógenos o marcarlos para la destrucción por otras células. La cascada está compuesta por muchas proteínas plasmáticas, sintetizadas en el hígado, primariamente por los hepatocitos. Las proteínas trabajan juntas para:
- Disparar el reclutamiento de las células inflamatorias.
- Marcar patógenos para su destrucción a cargo de otras células por opsonización, o recubrimiento, de la superficie del patógeno.
- Trastornar la membrana plasmática de una célula infectada, resultando en la citolisis de la célula infectada, causando la muerte del patógeno.
- Librar al cuerpo de los complejos antígeno-anticuerpo neutralizados.
Los elementos de la cascada de complemento pueden ser encontrados en muchas especies evolutivamente más viejos que en los mamíferos incluyendo las plantas, aves, peces y algunas especies de invertebrados.[4]
Células de la respuesta inmunitaria innata

Todas las células sanguíneas blancas (WBC) son conocidas como leucocitos. Los leucocitos son diferentes de otras células del cuerpo dado que no se asocian con ningún órgano en particular; así, su función está caracterizada por ser independiente, similar a los organismos unicelulares. Los leucocitos son capaces de moverse libremente e interactuar y capturar restos celulares, partículas extrañas, o de invadir microorganismos. Distinto a muchas otras células del cuerpo, la mayoría de los leucocitos inmunitarios innatos no se pueden dividir o reproducirse por sí mismos, pero son los productos de las células madres pluripotenciales presentes en la médula ósea.[1]
Los leucocitos de la respuesta innata incluyen: células asesinas naturales (células NK), mastocitos, eosinófilos, basófilos; y las células fagocíticas (fagocitos) incluyen a los macrófagos, los neutrófilos y las células dendríticas, y funcionan dentro del sistema inmunitario con la identificación y la eliminación de los patógenos que podrían causar infección.[2]
Fagocitos
La palabra fagocito significa literalmente comer células. Estas son células inmunitarias que devoran, patógenos o partículas. Para fagocitar un patógeno o partícula, un fagocito extiende porciones de su membrana celular, extendiendo la membrana alrededor del objeto a fagocitar (disponiendo la partícula dentro de la célula). Una vez dentro de la célula, el patógeno invasor es contenido dentro de un endosoma que se fusiona con un lisosoma.[2] El lisosoma contiene enzimas y ácidos que matan y digieren la partícula u organismo. Los fagocitos generalmente patrullan el cuerpo en busca de patógenos, pero también son capaces de reaccionar a un grupo de señales moleculares altamente especializadas producidas por otras células, llamadas citoquinas.
Las células fagocíticas del sistema inmunitario incluyen: macrófagos, neutrófilos, y células dendríticas. Los receptores de membrana de las células efectoras del sistema inmunitario innato que le permiten actuar en el reconocimiento de señales de peligro son del tipo TLR y PRR.
La fagocitosis de las células propias de los huéspedes es común como parte del desarrollo y el mantenimiento del tejido regular. Cuando el huésped muere, podría ser por procesos inducidos internamente a muerte celular programada (también llamada apoptosis), o causado por lesiones celulares debido a una infección bacteriana o viral, siendo las células fagocíticas las responsables de su remoción del sitio afectado.[1] Ayudando a remover las células muertas antes del crecimiento y desarrollo de nuevas células sanas, la fagocitosis es una parte importante del proceso de recuperación después de cualquier daño ocasionado a los tejidos.
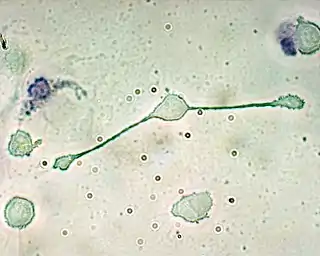
Macrófagos
Los macrófagos, vocablo proveniente del griego, significa "gran célula comedora", son leucocitos fagocíticos grandes, que son capaces de moverse al exterior del sistema vascular al atravesar la membrana celular de los vasos capilares y entrando en áreas intercelulares en persecución de los patógenos invasores. En los tejidos, los macrófagos órgano-específicos están diferenciados a partir de las células fagocíticas presentes en la sangre llamadas monocitos. Los macrófagos son los fagocitos más eficientes, y pueden fagocitar números substanciales de bacterias u otras células o microbios.[2]
La unión de moléculas bacteriales a los receptores sobre la superficie de un macrófago desencadena la ingestión y destrucción de las bacterias a través de la generación de una "brecha respiratoria", causando la liberación de especies reactivas del oxígeno. Los patógenos también estimulan al macrófago a la producción de quimioquinas, que atraen otras células al sitio de la infección.[2]
Neutrófilos

Los neutrófilos, al igual que otros dos tipos celulares: los eosinófilos y los basófilos (ver abajo), son conocidos como granulocitos debido a la presencia de gránulos en su citoplasma, o como células polimorfonucleares (PMNs) debido a su distintivos núcleos lobulados. Los gránulos del neutrófilo contienen una variedad de sustancias tóxicas que matan o inhiben el crecimiento de bacterias y hongos. Similares a los macrófagos, los neutrófilos atacan a los patógenos mediante la activación de un estallido respiratorio. Los productos principales del estallido del neutrófilo son fuertes agentes oxidantes incluyendo el peróxido de hidrógeno, los radicales libres de oxígeno y el hipoclorito. Los neutrófilos son los tipos celulares fagocíticos más abundantes, normalmente representan el 50 a 60 % del total de leucocitos circulantes, y son usualmente las primeras células en llegar al sitio de una infección.[3]
La médula ósea de un adulto normal saludable produce, según la fuente, más de 1 × 1011 (100 000 000 000) neutrófilos por día para unos[3]
o bien unos 1200 000 000 (1200 millones) para otros.[5]
Células dendríticas
Las células dendríticas (DC) son células fagocíticas presentes en tejidos que están en contacto con el medio externo, principalmente la piel (donde dichas células toman el nombre de células de Langerhans), y el revestimiento mucoso interno de la nariz, pulmones, estómago e intestino.[1] Se llaman así por su analogía con las dendritas neuronales, pero las células dendríticas no están conectadas al sistema nervioso. Las células dendríticas son muy importantes en la presentación de antígenos, y sirven como enlace entre los sistemas inmunitario innato y adaptativo.

Basófilos y eosinófilos
Los basófilos y los eosinófilos son células relacionadas con los neutrófilos (ver arriba). Cuando se activan al encuentro con un patógeno, los basófilos liberan histamina siendo importantes en la defensa contra parásitos, y juegan un papel importante en las reacciones alérgicas (tales como el asma).[2]
Desde su activación, los eosinófilos secretan un rango de proteínas altamente tóxicas y radicales libres que son altamente efectivos en la muerte de bacterias y de parásitos, pero también son responsables del daño tisular que ocurre durante las reacciones alérgicas. La activación y la liberación de toxinas por los eosinófilos es por tanto minuciosamente regulado para prevenir cualquier daño por la destrucción inadecuada de tejido.[3]
Células asesinas naturales
Las células asesinas naturales, o células NK (del inglés "Natural Killer"), son un componente de la respuesta inmunitaria innata. Las células NK atacan las células del huésped que han sido infectadas por microbios, pero no atacan directamente invadiendo los microbios. Por ejemplo, las células NK atacan y destruyen células tumorales, y las células infectadas por virus, mediante un proceso conocido como "auto-desaparición". Este término describe células con bajos niveles de un marcador celular de superficie llamado MHC I (complejo mayor de histocompatibilidad) —una situación que puede elevarse en infecciones virales de las células del huésped—.[4] Tales células se denominan "asesinas naturales" debido a la noción inicial de que no requerían activación para matar a las células que estaban "auto-desaparecidas".
Células T γδ
Como otras células T 'no convencionales' que llevan los invariables receptores de célula T (TCRs), tales como el CD1d-restringido en células asesinas naturales, las células γδ exponen características que las ubican en la frontera entre inmunidad adaptativa e inmunidad innata. Por un lado, las células T γδ podrían ser consideradas como un componente de la inmunidad adaptativa puesto que estas modifican los genes TCR para producir recombinación cruzada y desarrollar un fenotipo de memoria. Sin embargo, las múltiples combinaciones podrían también ser consideradas parte del sistema inmunitario innato donde los receptores restringidos de TCR o NK podrían ser usados como un modelo de reconocimiento del receptor. Por ejemplo, de acuerdo a este paradigma, grandes cantidades de células T Vγ9/Vδ2 responden en horas a moléculas comunes producidas por microbios, y las células T Vδ1 intraepiteliales altamente restringidos responderán a células epiteliales estresadas.
Patógeno-Especificidad
Las partes del sistema inmunitario innato tienen diferente especificidad para diferentes patógenos.
| Patógeno | Principales Ejemplos | [6] Fagocitosis | [6] Complemento | [6] Células asesinas naturales[6] |
|---|---|---|---|---|
| virus intracelular y citoplásmico | sí | no | sí | |
| bacterias intracelulares | sí (específicamente neutrófilos, no para rickettsia) | no | sí (no para rickettsia) | |
| bacterias extracelulares | sí | sí | no | |
| protozoos intracelulares | no | no | no | |
| protozoos extracelulares | sí | sí | no | |
| hongos extracelulares | no | sí | no | |
Evasión a la respuesta inmunitaria innata
Las células del sistema inmunitario innato previenen efectivamente el libre crecimiento de las bacterias dentro del cuerpo; sin embargo, muchos patógenos han desarrollado mecanismos que les permiten evadir el sistema inmunitario innato.[7][8] Las estrategias de evasión que engañan a la inmunidad innata incluyen la replicación intracelular, como ocurre en Salmonella, o una cápsula protectora que previene la lisis por complemento y por los fagocitosis, visto en el Mycobacterium Tuberculosis'.[9]
Las especies Bacteroides son bacterias comensales normalmente, distribuidas en una porción substancial de la flora gastrointestinal de los mamíferos.[10] Algunas especies (B. fragilis, por ejemplo) son patógenos oportunistas, causando infecciones de la cavidad peritoneal. Estas especies evaden el sistema inmunitario mediante la inhibición de la fagocitosis afectando los receptores que los fagocitos usan para fagocitar bacterias o imitando las células del huésped para que el sistema inmunitario no los reconozca como extraños. Staphylococcus aureus inhibe la habilidad del fagocito para responder a las señales por quimioquinas. Otros organismos tales como M. tuberculosis, Streptococcus pyogenes and Bacillus anthracis utilizan mecanismos que matan directamente al fagocito.
Las bacterias y los hongos también pueden formar biopelículas complejas, proporcionando protección de las células y proteínas del sistema inmunitario; estudios recientes indican que tales biopelículas están presentes en múltiples infecciones exitosas, incluyendo infecciones por Pseudomonas aeruginosa crónica y Burkholderia cenocepacia características de la fibrosis quística.[11]
Otras formas de inmunidad innata
Defensa del huésped en procariotas
Las bacterias (y tal vez otros organismos procariotas), utilizan un mecanismo de defensa único, llamado sistema de modificación a la restricción que los protege de patógenos, tales como los bacteriófagos. En este sistema, las bacterias producen enzimas, denominadas endonucleasas de restricción, que atacan y destruyen regiones específicas del ADN viral de los bacteriófagos invasores. La metilación en el ADN propio del huésped lo marca como propio y lo previene de ser atacado por endonucleasas.[12] Las endonucleasas de restricción y el sistema de modificación por restricción existen exclusivamente en procariotas.
Defensa del huésped en invertebrados
Los invertebrados no poseen linfocitos u otro sistema humoral basado en anticuerpos, y es probable que un multicomponente, el sistema inmunitario adaptativo surgiera en los primeros vertebrados.[13] No obstante, los invertebrados poseen mecanismo que parecen ser precursores de estos aspectos de la inmunidad en vertebrados. El modelo de reconocimiento de receptores es un grupo de proteínas usadas por todos los organismos cercanamente para identificar moléculas asociadas con patógenos microbianos. Los receptores Toll (TLR) son una clase importante del modelo de reconocimiento de receptor, que existe en todos los celomados (animales con una cavidad corporal), incluyendo los humanos.[14] El sistema del complemento, como se discute abajo, es una cascada bioquímica del sistema inmunitario que ayuda a eliminar patógenos de un organismo, y existe en la mayoría de las formas de vida. Algunos invertebrados, incluyendo varios insectos, cangrejos, y gusanos utilizan una forma modificada de respuesta del complemento conocida como el sistema profenoloxidasa (proPO).[13] Los péptidos antimicrobianos son un componente conservado evolutivamente de la respuesta immunológica innata encontrada en todas las clases de vida y representa la principal forma de vida de la inmunidad sistémica en invertebrados. Varias especies de insectos producen péptidos antimicrobianos conocidos como defensinas y cecropinas.
Defensa del huésped en plantas
Los miembros de cada clase de patógeno que infecta a los humanos también infecta a las plantas. Aunque las especies patógenicas exactas varían con las especies infectadas, bacterias, hongos, virus, nemátodos e insectos pueden causar enfermedades de las plantas. Como con los animales, las plantas atacadas por insectos u otros patógenos usan un grupo de complejas respuestas metabólicas que llevan a la formación de compuestos químicos defensivos que combaten la infección o hacen la planta menos atractiva hacia los insectos y otros herbívoros.[15]
Como los invertebrados, las plantas ni generan respuestas a anticuerpos o células T ni poseen células móviles que detecten y ataquen a los patógenos. En cambio, en caso de infección, partes de algunas plantas son tratadas como desechable y reemplazable, en formas que muy pocos animales son capaces de hacer. Excluir o eliminar una parte de una planta ayuda a detener la propagación de una infección.[15]
La mayoría de las respuestas inmunitarias en plantas involucran señales químicas sistémicas enviadas a través de la planta. Las plantas usan un modelo de reconocimiento de receptores para identificar los patógenos y empezar una respuesta basal, la cual produce señales químicas que ayudan a detener la infección. Cuando una parte de una planta se infecta con un patógeno microbiano o viral, en caso de una interacción incompatible desencadenada por ‘’elicitors’’ específicos, la planta produce una respuesta hipersensitiva (HR) localizada, donde las células en el sitio de la infección experimentan una muerte celular programada rápida para prevenir la propagación de una enfermedad a otras partes de la planta. La HR tiene algunas similitudes a la piroptosis animal, tales como un requerimiento de la actividad proteolítica de VPEγ similar a caspasa-1, una proteasa de cisteína que regula el desensamblaje de la célula durante la muerte celular.[16]
Las proteínas de "Resistencia" (R), codificadas por los genes R, están presentes ampliamente en plantas y detectan patógenos. Estas proteínas contienen dominios similares a los receptores NOD y receptores de tipo Toll utilizados en la inmunidad animal innata. La resistencia sistémica adquirida(SAR) es un tipo de respuesta defensive que ofrece resistencia a toda la planta para un amplio espectro de agentes infecciosos. La SAR involucra la producción de mensajeros químicos, tales como el ácido salicílico o el ácido jasmónico. Algunos de estos viajan a través de la planta y señaliza otras células para producir compuestos defensivos para proteger las partes no infectadas, por ejemplo, las hojas. El ácido salicílico en sí mismo, aunque indispensable para la expresión de SAR, no es la señal translocada responsible de la respuesta sistémica. Evidencia reciente indica un papel para los jasmonados en la transmisión de la señal a las porciones distales de la planta.
Los mecanismos de silenciamiento del RNA también son importantes en la respuesta sistémica de la planta, como ellas pueden bloquear la replicación del virus.[17] La respuesta del ácido jasmónico, es estimulada en las hojas dañadas por los insectos, e involucra la producción de metil-jasmonato.[15]
Véase también
Referencias
- Alberts, Bruce; Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, and Peter Walters (2002). Molecular Biology of the Cell; Fourth Edition. New York and London: Garland Science. ISBN 0-8153-3218-1.
- Janeway, Charles; Paul Travers, Mark Walport, and Mark Shlomchik (2001). Immunobiology; Fifth Edition. New York and London: Garland Science. ISBN 0-8153-4101-6. .
- Stvrtinová, Viera; Ján Jakubovský and Ivan Hulín (1995). Inflammation and Fever from Pathophysiology: Principles of Disease. Computing Centre, Slovak Academy of Sciences: Academic Electronic Press. Archivado desde el original el 18 de junio de 2007. Consultado el 10 de febrero de 2008.
- Janeway CA, Jr. et al (2005). Immunobiology. (6ta ed. edición). Garland Science. ISBN 0-443-07310-4.
- Delgado J.L. (2010). Cap.23: Fisiología de los granulocitos. «Fisiología Humana». AccesMedicine (4a. edición).
- Unless else specified in boxes, then ref is: Lippincott's Illustrated Reviews: Immunology. Paperback: 384 pages. Publisher: Lippincott Williams & Wilkins; (July 1, 2007). Language: English. ISBN 0781795435. ISBN 978-0781795432. Page 172
- Kennedy, Alan. «Immune Evasion by bacteria». Archivado desde el original el 10 de octubre de 2006. Consultado el 11 de febrero de 2008.
- Finlay B, McFadden G (2006). «Anti-immunology: evasion of the host immune system by bacterial and viral pathogens». Cell 124 (4): 767-82. PMID 16497587.
- Finlay B, Falkow S (1997). «Common themes in microbial pathogenicity revisited». Microbiol Mol Biol Rev 61 (2): 136-69. PMID 9184008.
- Dorland WAN (editor) (2003). Dorland's Illustrated Medical Dictionary (30th edición). W.B. Saunders. ISBN 0-7216-0146-4.
- Kobayashi H (2005). «Airway biofilms: implications for pathogenesis and therapy of respiratory tract infections». Treat Respir Med 4 (4): 241-53. PMID 16086598.
- Restriction Enzymes Access Excellence Classic Collection Background Paper.
- Beck, Gregory and Habicht, Gail S. Immunity and the Invertebrates Scientific American. November 1996:60-66.
- Imler JL, Hoffmann JA. (2001) Toll receptors in innate immunity. Trends Cell Biol. Jul;11(7):304-11. Review. PMID 11413042
- Schneider, David (2005) Plant immune responses Stanford University Department of Microbiology and Immunology.
- Rojo, E. et al. (2004). «VPEgamma exhibits a caspase-like activity that contributes to defense against pathogens.». Curr Biol. 14 (21): 1897-1906. PMID 15530390.
- Baulcombe D (2004). «RNA silencing in plants». Nature 431 (7006): 356-63. PMID 15372043.
| Control de autoridades |
|
|---|
 Datos: Q428253
Datos: Q428253 Multimedia: Innate immune system / Q428253
Multimedia: Innate immune system / Q428253