Islotes pancreáticos
Los islotes pancreáticos o islotes de Langerhans son estructuras formadas por cúmulos densos y compactos de células con función exclusivamente endócrina, del páncreas, que se encargan de producir hormonas como la insulina y el glucagón. El islote ha sido descrito como un mini-órgano complejo, compuesto por un ensamblaje de células endócrinas, y con otras células de apoyo, que controlan la homeostasis de la glucosa en sangre.
En el humano se calcula en ~1 millón ( 1 × 106) el número de islotes en el páncreas.
Cada islote está formado por ~1000 células endócrinas de diferente tipo: células alfa (α), células beta (β), células delta (δ), células gama (γ), células G y células epsilon (ε).
Por el contrario, los acinos del páncreas son las glándulas de secreción externa (exócrinas) del páncreas, encargadas de secretar enzimas dentro del tubo digestivo.
| Islotes pancreáticos | ||
|---|---|---|
 | ||
| Nombre y clasificación | ||
| Sinónimos |
| |
| Latín | [TA]: insulae pancreaticae | |
| TA | A05.9.01.019 | |
| Información anatómica | ||
| Sistema | Endócrino | |
| Arteria |
Arteria pancreaticoduodenal superior Ramas pancreáticas de la arteria esplénica Arteriolas insulares | |
|
| ||
Historia
En 1869 Paul Langerhans publicó en su Tesis el descubrimiento microscópico de acúmulos de células dentro del páncreas:
“En su mayoría, varias células están muy próximas, distribuidas peculiarmente en el parénquima de la glándula (…) los cúmulos "zellhaufen" redondeados tienen un diámetro de 0,1 a 0,24 mm”. Paul Langerhans, Tesis: Beitrage zur mikroskopischen Anatomie der Bauchspeicheldruse (1869).[1] |
En 1893 Edouard Laguesse los llamó «islotes de Langerhans» en reconocimiento a su descubridor.[1]
En 1968 Dean y Matthews por primera vez, atravesaron islotes de ratón intactos con electrodos intracelulares afilados y reportaron la actividad eléctrica de las células pancreáticas.[2]
Estructura

Los vasos sanguíneos en rojo y los nervios en blanco.
Los acúmulos de células endócrinas del páncreas forman pequeños racimos o islotes, dispersos por todo el órgano. Las células ubicadas en el interior de los islotes, se encuentran densamente empaquetadas. Esta disposición hace que el #Espacio intercelular sea escaso entre los diferentes tipos de células.
Las células endócrinas del islote se presentan como un ensamblaje ordenado, en el cual las células secretoras interactúan, para mantener una salida hormonal óptima.[3]
Hay más de un millón (hasta 3 millones 1-3 × 106) de tales islotes en el páncreas humano normal.[4]
Los islotes están compuestos además por células vasculares (células endoteliales, pericitos), eritrocitos, células del estroma (fibroblastos), células inmunitarias (granulocitos, linfocitos, macrófagos, células dendríticas) y células nerviosas (neuronas).[5]
Microaquitectura
Los Islotes pancreáticos tienen un tamaño entre 100-500 micrómetros (μm) y cada uno agrupa ~1000 células endócrinas de diferente tipo.[6]

En los cortes teñidos con H&E, tienen el aspecto de islotes irregulares de color rosa pálido, distribuidos extensamente entre un mar de acinos exocrinos de color más oscuro.
Para identificar los diferentes tipos de células insulares se necesitan métodos de tinción especial como el de Gomori o tinción mediante inmunohistoquímica o inmunofluorescencia.
En el Islote pancreático la proporción de las diferentes células endócrinas varía según la especie.[7]
La localización tridimensional de las células dentro del islote también es diferente.[8]
Gran parte de estas células endócrinas son llamadas células β (beta) relativamente pequeñas y de color azul.
En grupos pequeños alrededor de las células beta, se identifican células de mayor tamaño, de color rosa que son denominadas células α (alfa).
Además, los islotes poseen también células delta (δ), células F, PP ( γ)
y epsilon (ε), células G. Las células beta son las más comunes y las epsilon las más raras.[9]
La arquitectura espacial de los tipos de células endócrinas dentro del islote es importante para su función, ya que la conexión intercelular y la señalización parácrina entre células optimizan la secreción de hormonas.[10]
Espacio intercelular
Dentro de los islotes, el intersticio es un espacio crucial para el diálogo entre las células y para la transducción de señales. Se muestra como un «canalículo» o espacio intercelular común, en ubicación opuesta a los capilares, formado por tres o cuatro células adyacentes, y que se encuentra recubierto con microvellosidades.[11] Dentro de este espacio, emerge la cilia primaria desde las células, como una estructura especializada que media esos procesos.[12]
- Islotes murinos
Los islotes de ratón/rata muestran una arquitectura típica de dos zonas homogéneas núcleo y manto, con células β agrupadas en el centro, rodeadas por un manto delgado de células α y δ.
- Islotes humanos
En los islotes pancreáticos humanos las células β están entremezcladas (heterogéneas) con las células α, δ, G, γ y ε.[8][13][14][15]
| Células Alfa (α) glucagón |
Células beta (β) insulina |
Células delta (δ) somatostatina |
Células gama (γ, F, PP) polipéptido pancreático |
Células (G) gastrina |
Células epsilon (ε) grelina | |
|---|---|---|---|---|---|---|
| Murinos | 15% | 75% | 10% | 5% | - | 1% |
| Humano | 33-46% [13] 34.4 ±2.5% [14] | 48-60% 53.9 ±2.5% | 8-12% 10.4 ±0.9% | 2%[15] - | - - |
- - |
Las sustancias que producen cada una de estas células son:
- Células alfa (α): producen la hormona polipeptídica glucagón.
- Células beta (β): producen las hormonas polipeptídicas insulina y amilina (a una razón de 100:1 respectivamente).
- Células delta (δ): producen el péptido somatostatina.
- Células gama ( γ) F: producen un polipéptido pancreático que inhibe las secreciones exocrinas del páncreas y la motilidad del estómago y del intestino.
- Células G: producen la hormona polipéptida gastrina que estimula la producción de HCl por las células parietales del estómago.
- Células epsilon (ε): producen la hormona grelina.[16][17]
Circulación en el islote
Los islotes representan solamente 1-2% del total de la masa pancreática, pero están densamente vascularizados y abastecidos por 5-15% del flujo sanguíneo que ingresa al páncreas.[18]

Los islotes están irrigados por arteriolas, procedentes del
estroma circundante. El tamaño del islote entre 50-100 micrómetros (µm) determina el diámetro de la arteriola aferente. La arteriola al entrar en el islote se divide en capilares. El lecho capilar del islote pancreático muestra una disposición en ovillos y bucles cercanamente enrollados. La vasculatura capilar es densa y se compone de células endoteliales que son delgadas y muy fenestradas.
El escaso tejido fibroso/conjuntivo que forma la matriz extracelular rodea el islote y acompaña las arteriolas y las vénulas.[19]
Linfáticos en el islote
Durante años se había considerado que los islotes carecían de vasos linfáticos funcionales.

Arteriolas en amarillo.
Los vasos linfáticos que rodean algunos de los islotes, penetran en ellos solo en el hilio.
Los linfáticos peri-islotes e intra-islotes podrían participar en la regulación de la inflamación, un hecho común en la diabetes.
Los estudios muestran que los vasos linfáticos (en ratones no diabéticos) están adyacentes a los islotes inflamados, y también se encuentra una red linfática funcional en los islotes que son trasplantados.[20]
Inervación del islote


Los islotes están inervados por nervios colinérgicos simpáticos, adrenérgicos y parasimpáticos.[21]
Los nervios extrapancreáticos se proyectan directamente a los islotes pancreáticos o bien convergen en los ganglios intrapancreáticos.[22]
- Inervación pancreática intrínseca
es una red de neuronas pancreáticas, que se agrupan para formar ganglios pancreáticos y envían proyecciones amielínicas a sus objetivos en el páncreas endocrino y exocrino. Los ganglios están dispersos por el parénquima pancreático y los espacios interlobulillares, pero también están presentes alrededor de los islotes pancreáticos, donde forman complejos neuroinsulares.[23]
Pocos axones colinérgicos parasimpáticos parecen penetrar en el islote humano y las fibras simpáticas inervan preferentemente las células del músculo liso de los vasos sanguíneos ubicados dentro del islote.[24]
Genética celular
La Ontogenia de los diferentes tipos de células de los islotes se ha estudiado ampliamente en ratones, donde los progenitores endocrinos comienzan a expresar ARX o PAX4, empujando su especificación hacia el destino de las células α o β.
Los factores de transcripción específicos del tipo de célula guían a las células hacia la maduración.
Para las células α, los factores de transcripción incluyen: MAFB, IRX2 y BRN4.
Son factores de transcripción clave para la maduración de las células β: PDX1, MAFA y NKX6.1.
El desarrollo de la secuenciación de ARN de una sola célula (scRNA-Seq en inglés) permite perfilar el contenido de ARNm de células individuales de tejidos heterogéneos. Ofrece una oportunidad para caracterizar mejor las identidades celulares de las diferentes células de los islotes pancreáticos humanos.
Los conjuntos de genes definen con precisión los diferentes tipos de células de los islotes.[25]
Los genes relacionados con las células α: GCG, ARX, FEV y TM4SF4.
Los genes clave relacionados con las identidades de las células β: INS, IAPP, DLK1 y RBP4.
Los genes relacionados con las células γ: PPY, ID2, ETV1.
Genes clave relacionados con las identidades de las células δ: SST, RBP4, HHEX, LEPR.[16]
Fisiología de los islotes
Cada islote es una red densamente empaquetada de células, compuesta por varios tipos celulares con una función endócrina que controla a nivel corporal: la homeostasis de la glucosa, el metabolismo y el comportamiento alimentario.[26]

Los islotes basan su funcionalidad en una variedad de señales quimiotácticas y de adhesión, para establecer y organizar las relaciones intercelulares. El contacto físico y las señales secretadas entre ciertos tipos de células optimizan la secreción de hormonas.[27][10]
El islote pancreático funciona como un solo órgano con señalización coordinada de manera estrecha entre los diferentes tipos de células. Esta red permite la respuesta a los cambios en la glucosa en sangre y a las señales dentro del mismo islote (a través de uniones comunicantes o señalización parácrina) e impulsos nerviosos extrínsecos, de manera rápida y sensible.[21]
Dentro de la estructura del islote, las hormonas forman una complicada red de funciones paracrinas, a las que se suman moléculas de señalización secundarias, que regulan la función y la supervivencia de las células.[26]
Electrofisiología
Vm = voltaje de membrana; mV = milivoltios; s = segundos. Electrofisiología.
Los primeros estudios de electrofisiología en islotes de ratón intactos, atravesados con electrodos, decribieron la actividad eléctrica compuesta, producida por las células beta, que ocupan el sector central.
En condiciones normales, la actividad eléctrica del islote aislado se registra definida, periódica, y estable durante largos períodos de tiempo.
En condiciones basales se registran potenciales en picos sincronizados de hasta 0,5 mV de amplitud, con 200-1000 milisegundos (ms) de duración y una frecuencia estable de 0,5-2 Hertz (Hz).[28]
En células aisladas de islotes de ratones, ratas, conejillos de indias y humanos, los estudios han revelado que son eléctricamente excitables y que contienen una serie de canales iónicos dependientes e independientes del voltaje en su membrana plasmática.[2]
En islotes humanos y de ratón, la actividad eléctrica de las células β dentro de un islote, que determina la secreción de insulina, experimenta oscilaciones rápidas (períodos de decenas de segundos) y lentas (períodos de varios minutos) en respuesta a un estímulo de glucosa.[29]
El sistema nervioso autónomo regula la secreción de hormonas del páncreas endócrino, y por eso, afecta el metabolismo de la glucosa. Los sistemas nerviosos parasimpático y sistema nervioso simpático inervan directamente el islote pancreático.[24]
Como micro-órgano endócrino, el islote necesita un flujo sanguíneo constante, para así poder detectar y responder con rapidez a los cambios de sustancias en el plasma (sangre), in vivo cada célula beta secretora de insulina, parece estar asociada con al menos una célula endotelial.[30][31]
Función celular

El islote ha sido descrito como un mini-órgano complejo, compuesto por una variedad de células endócrinas y otras células de apoyo y sostén, que controlan de forma estrecha la homeostasis de la glucosa en sangre.[9][32]
Las células endocrinas están encerradas por una capa de membrana basal y matriz extracelular (BM-ECM), que las separa del tejido exocrino y son alimentadas por una vasculatura independiente formada por una densa red de capilares.[33]
Células alfa y glucagón
Los gránulos secretores que contienen glucagón de las células alfa (α) con un diámetro de 250-300 nanómetros (nm), que poseen un contenido electrodenso que llena las vesículas. Tienen una gran zona central esférica muy electrodensa rodeada por un reborde angosto menos electronicodenso.
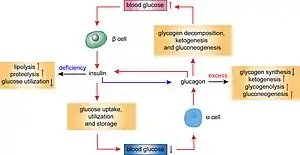
El glucagón aumenta el nivel de glucosa sanguínea (glucemia) al estimular la formación de este carbohidrato a partir del glucógeno almacenado en hepatocitos (glucogenolisis). También ejerce efecto en el metabolismo de proteínas y grasas. La liberación del glucagón es inhibida por la hiperglucemia.
Células beta e insulina
Las células beta (β) individuales son heterogéneas dentro del mismo islote, presentan propiedades funcionales diferentes y en su nivel de liberación de insulina.[34]
En islotes humanos y de ratón, la actividad eléctrica de las células β dentro de un islote, que determina la secreción de insulina, experimenta oscilaciones rápidas (períodos de decenas de segundos) y lentas (períodos de varios minutos) en respuesta a un estímulo de glucosa.[29]
La insulina es sintetizada en el retículo endoplásmico rugoso en forma de un polipéptido llamado pre-pro-insulina que se transforma en pro-insulina, que posee la misma actividad hormonal aunque no de la misma magnitud que la insulina. La proinsulina se modifica en el aparato de Golgi y las vesículas secretoras que salen de este complejo contienen la hormona insulina. La insulina es secretada en reacción a la hiperglucemia y también por algunas hormonas péptidas como glucagón, colecistocinina-pancreocimina y secretina.

Sus acciones principales son estimular la captación de glucosa en varios tipos de células, y disminuir el nivel de glucosa sanguínea, al estimular la conversión de glucosa en glucógeno en los hepatocitos y miocitos, siempre que dicho nivel aumente.
Células delta y somatostatina
Las células delta (δ) poseen gránulos secretores más grandes y menos electrodensos que las células alfa y beta, y sus organelos secretores son menos notables.
La somatostatina es una neurohormona péptida y neurotransmisora que inhibe la liberación de la hormona del crecimiento, de la insulina, el glucagón e incluso de la propia somatostatina pancreática. La regulación mutua de la actividad secretora entre las células alfa, beta y delta sugiere fuertemente alguna disposición especial que facilita la regulación directa de una célula con otra, mecanismo conocido como regulación paracrina.
Células gama y polipéptido pancreático
Las células gama (γ) antes conocidas como F y como PP, se encuentran en la periferia del islote. Son pequeñas y redondeadas y secretan el llamado polipéptido pancreático (PP).
La secreción de polipéptido pancreático es una parte importante del eje páncreas-intestino-cerebro. El PP tiene acciones inhibitorias en gran medida en el intestino, lo que reduce el vaciamiento gástrico y la actividad motora intestinal a través de acciones en el receptor Y4 (PPYR1).[21]
Células G y gastrina
Las células G del islote pancreático, secretan la hormona llamada gastrina que, tras ser liberada, pasa a la sangre y estimula el vaciamiento gástrico y la producción de ácido clorhídrico por las células parietales del estómago.[35][36]
Células epsilon y grelina
Las células epsilon (ε) secretan la hormona llamada grelina que, tras ser liberada, pasa a la sangre, estimula la secreción de hormona del crecimiento (GH) en la hipófisis y favorece la regulación del metabolismo energético regulando el peso corporal y la ingesta por el aumento del apetito.[16][37]
Trasplante de islotes
El trasplante de células de los islotes tiene un potencial considerable como cura para la diabetes tipo 1 enfermedad por destrucción de las células β del páncreas, que sufre 5-10 % de todas las personas con diabetes.
En el año 2000, una cohorte de siete pacientes permaneció independiente de la insulina durante un año después del trasplante, manteniendo un régimen inmunosupresor.[38]
El éxito del trasplante mantiene la glucemia normal a corto plazo, pero los resultados a largo plazo son desconocidos. Son necesarias inyecciones adicionales de preparados de islotes para obtener una independencia prolongada de la insulina.[39][8]
Las estrategias de reemplazo de células se han llevado a cabo en varios sitios del cuerpo: hígado, músculo, epiplón y tejido subcutáneo.[40]
Las reacciones inmunitarias posteriores al trasplante generan el rechazo del injerto y representan un cuello de botella en el campo del trasplante de islotes.
La fisiología demuestra que la insulina por sí sola no restaura la función completa de los islotes y por qué es tan difícil lograr la normoglucemia sin la existencia de nuevos islotes que funcionen.[41]
Patología
Tumores de los islotes del páncreas
Los tumores neuroendocrinos pancreáticos son los tumores de las células de los islotes, es un tipo de cáncer que comienza en el páncreas.
La mitad de los tumores neuroendocrinos pancreáticos producen hormonas que son liberadas a la sangre y causan síntomas.[42]
Las células de algunos tumores neuroendocrinos pancreáticos continúan secretando hormonas y se llaman tumores funcionales, lo que genera una cantidad excesiva de una determinada hormona en el cuerpo.[43]
Los insulinomas se originan de las células productoras de insulina.
Los glucagonomas surgen de las células productoras de glucagón.
Los gastrinomas surgen de las células productoras de gastrina.
Los somatostatinomas se originan de las células productoras de somatostatina.
Los VIPomas surgen de las células productoras de péptido intestinal vasoactivo (VIP).
Los tumores que secretan ACTH provienen de células que producen la hormona adrenocorticotrópica.
La mayoría (hasta el 70%) de los tumores neuroendocrinos pancreáticos funcionales son insulinomas.[42]
Referencias
- Pascal de Boer; Ben NG Giepmans (2021). «State‐of‐the‐art microscopy to understand islets of Langerhans: what to expect next?». Immunol Cell Biol. 99 (5): 509-520. PMC 8252556. PMID 33667022. doi:10.1111/imcb.12450. Consultado el 25 de febrero de 2023.
- Rorsman P.; Braun M.; Zhang Q. (2012). «Regulation of calcium in pancreatic α- and β-cells in health and disease». Cell Calcium 51 (3-4): 300-308. PMC 3334273. PMID 22177710. doi:10.1016/j.ceca.2011.11.006. Consultado el 6 de abril de 2023.

- Paolo MEDA; Alain PERRELET; Lelio ORCI (1979). «Increase of GAP junctions between pancreatic B-cells during stimulation of insulin secretion» (PDF). J. CELL BIOLOGY (The Rockefeller University Press) 82: 441-448.

- Ionescu-Tirgoviste C.; Gagniuc P.A.; Gubceac E.; Mardare L.; Popescu I.; Dima S.; Militaru M. (2015). «A 3D map of the islet routes throughout the healthy human pancreas.». En Nature research, ed. Scientific Reports 5: 14634. PMC 4586491. PMID 26417671. doi:10.1038/srep14634. Consultado el 26 de marzo de 2023.
- Abdulreda M.H.; Berggren P.O. (2021). «The pancreatic islet: a micro-organ in control». CellR4 Repair Replace Regen Reprogram. (en inglés) 9: e3093. PMC 8017525. doi:10.32113/cellr4_20213_3093. Consultado el 14 de octubre de 2022.

- Alfredo Jácome Roca; Enrique Ardila Ardila; Luz Angela Casas Figueroa (2022). «5: Páncreas endocrino». Fisiología Endócrina (4.a edición). El Manual Moderno. p. 97. Consultado el 12 de septiembre de 2022.
- Steiner D.J.; Kim A.; Miller K.; Hara M. (2010). «Pancreatic islet plasticity: interspecies comparison of islet architecture and composition». Islets (REVISIÓN) 2 (3): 135-145. doi:10.4161/isl.2.3.11815. Consultado el 1 de marzo de 2023.
- Hoang D-T.; Matsunari H.; Nagaya M.; Nagashima H.; Millis JM.; Witkowski P.; Periwal V.; Hara M.;Jo J. (2014). «A Conserved Rule for Pancreatic Islet Organization.». PLoS ONE (en inglés) (Public Library of Science) 9 (10): e110384. doi:10.1371/journal.pone.0110384. Consultado el 1 de octubre de 2022.
- Noguchi G.M.; Huising M.O (2019). «Integrating the inputs that shape pancreatic islet hormone release». Nature Metabolism (Artículo de revisión) (en inglés) 1. pp. 1189-1201.
- Waters B.J.; Blum B. (2022). «Axon Guidance Molecules in the Islets of Langerhans». Front. Endocrinol. (REVISIÓN). Sec. Diabetes: Molecular Mechanisms (en inglés). Consultado el 2 de octubre de 2022.
- Yamamoto M.; Kataoka K. (1986). «Electron microscopic observation of the primary cilium in the pancreatic islets». Arch Histol Jpn (PDF) (en inglés) (J-STAGE) 49 (4): 449-457. doi:10.1679/aohc.49.449.
- Melena I.; Hughes J.W. (2022). «Islet cilia and glucose homeostasis». Front Cell Dev Biol. (REVISIÓN) (Frontiers (editorial)) 10 (1082193). PMC 9751591. PMID 36531945. doi:10.3389/fcell.2022.1082193. Consultado el 8 de mayo de 2023.
- Cabrera O.; Bergman D.M; Kenyon N.S.; Ricordi C.; Berggren P-O.; Caicedo A. (2006). «The unique cytoarchitecture of human pancreatic islets has implications for islet cell function». Proc Natl Acad Sci USA. Medical Sciences (en inglés) 103 (7): 2334-2339. PMC 1413730. PMID 16461897. doi:10.1073/pnas.0510790103. Consultado el 12 de setiembre de 2022.

- Brissova M.; Fowler M.J.; Nicholson W.E; Chu A.; Hirshberg B.; Harlan D.M.; Powers A.C. (2005). «Assessment of Human Pancreatic Islet Architecture and Composition by Laser Scanning Confocal Microscopy». Journal of Histochemistry & Cytochemistry (en inglés) (SAGE) 53 (9): 1087-1097. Consultado el 12 de setiembre de 2022.
- Wang X.; Zielinski MC.; Misawa R.; Wen P.; Wang T-Y.; Wang C-Z.; Witkowski P.; Hara M. (2013). «Quantitative Analysis of Pancreatic Polypeptide Cell Distribution in the Human Pancreas». PLoS ONE (en inglés) (Public Library of Science) 8 (1): e55501. Consultado el 16 de enero de 2018.
- Wierup N.; Sundler F.; Heller R.S. (2013). «The islet ghrelin cell». J Mol Endocrinol (Artículo de revisión) (en inglés) 52 (1): R35-49. doi:10.1530/JME-13-0122. Consultado el 13 de setiembre de 2022.
- Quesada Candela C. (2016). p7. «Adaptaciones de las poblaciones celulares alfa y beta del islote de Langerhans a la preñez en ratonas C57BL/6». Revista Doctorado (Unidad de Fisiología Celular y Nutrición. Instituto de Bioingeniería. Universidad Miguel Hernández de Elche (UMH)) 2 (1).
- Shah P.; Lueschen N.; Ardestani A.; Oberholzer J.; Olerud J.; Carlsson P-O, et al. (2016). «Angiopoetin-2 Signals Do Not Mediate the Hypervascularization of Islets in Type 2 Diabetes». PLOS ONE (en inglés) (Public Library of Science) 11 (9): e0161834. Consultado el 20 de mayo de 2018.
- Miller K.; Kim A.; Kilimnik G.; Jo J.; Moka U.; Periwal V.; Hara M. (2009). «Islet Formation during the Neonatal Development in Mice.». PLoS ONE (en inglés) (Public Library of Science) 4 (11): e7739. doi:10.1371/journal.pone.0007739. Consultado el 6 de abril de 2023.
- Yin N.; Zhang N.; Lal G.; Xu J.; Yan M.; Ding Y.; Bromberg J.S. (2011). «Lymphangiogenesis Is Required for Pancreatic Islet Inflammation and Diabetes». PLoS ONE (en inglés) (Public Library of Science) 6 (11): e28023. doi:10.1371/journal.pone.0028023. Consultado el 26 de setiembre de 2022.
- Brereton M.F.; Vergari E.; Zhang Q.; Clark A. (2015). «Alpha-, Delta- and PP-cells. Are They the Architectural Cornerstones of Islet Structure and Co-ordination?». J Histochem Cytochem. (en inglés) 63 (8): 575-591. Consultado el 2 de octubre de 2022.
- Li W.; Yu G.; Liu Y.; Sha L. (2019). «Intrapancreatic Ganglia and Neural Regulation of Pancreatic Endocrine Secretion». Front. Neurosci. (REVISIÓN) (en inglés) (Frontiers (editorial)) 13 (21). doi:10.3389/fnins.2019.00021. Consultado el 15 de octubre de 2022.
- Makhmutova M.; Caicedo A. (2021). «Optical Imaging of Pancreatic Innervation». Front. Endocrinol. (REVISIÓN). Diabetes: Molecular Mechanisms (en inglés) (Frontiers (editorial)). Consultado el 20 de octubre de 2022.
- Rodriguez-Diaz R.; Abdulreda M.H.; Formoso A.L.; Gans I.; Ricordi C.; Berggren P-O; Caicedo A. (2011). «Autonomic axons in the human endocrine pancreas show unique innervation patterns». Cell Metab. (en inglés) 14 (1): 45-54. PMC 3135265. PMID 21723503. doi:10.1016/j.cmet.2011.05.008. Consultado el 14 de octubre de 2022.
- van Gurp L.; Fodoulian L.; Oropeza D.; Furuyama K.; Bru Tari E.; Nguyet Vu A.; Kaddis J.S.; Rodríguez I.; Thorel F.; Herrera P.L. (2022). «Generation of human islet cell type-specific identity genesets». Nature Communications (en inglés) 13 (2020). Consultado el 13 de setiembre de 2022.
- Hartig S.M.; Cox A.R. (2020). «Paracrine signaling in islet function and survival». J Mol Med (Berl). (REVISIÓN) (en inglés) (Springer) 98 (4): 451-467. PMC 7899133. PMID 32067063. doi:10.1007/s00109-020-01887-x. Consultado el 9 de abril de 2023.
- Paolo Meda (2013). «Protein-mediated interactions of pancreatic islet cells». Scientifica (Cairo) (REVISIÓN) (Hindawi): 2013:621249. PMC 3820362. PMID 24278783. doi:10.1155/2013/621249. Consultado el 13 de abril de 2023.
- Palti Y.; Ben David G.; Lachov E.; Mika Y.H.; Omri G.; Schatzberger R. (1996). «Islets of Langerhans Generate Wavelike Electric Activity Modulated by Glucose Concentration» (PDF). DIABETES (en inglés) 45: 595-601. Consultado el 6 de abril de 2023.
- Hogan JP.; Peercy BE. (2021). «Flipping the switch on the hub cell: Islet desynchronization through cell silencing.». PLoS ONE (Public Library of Science) 16 (4): e0248974. doi:10.1371/journal.pone.0248974. Consultado el 17 de mayo de 2023.
- Sankar KS.; Green BJ.; Crocker AR.; Verity JE.; Altamentova SM.; Rocheleau JV. (2011). «Culturing Pancreatic Islets in Microfluidic Flow Enhances Morphology of the Associated Endothelial Cells». PLOS ONE (en inglés) (Public Library of Science) 6 (9): e24904. Consultado el 22 de enero de 2018.
- Rorsman P.; Ashcroft F.M. (2018). «Pancreatic β-Cell Electrical Activity and Insulin Secretion: Of Mice and Men». Physiol Rev. (REVISIÓN) (en inglés) 98 (1): 117-214. PMC 5866358. PMID 29212789. doi:10.1152/physrev.00008.2017. Consultado el 22 de abril de 2023.
- Huising M.O. (2020). «Paracrine regulation of insulin secretion». Diabetologia (REVISIÓN) (en inglés) (Springer) 63 (10): 2057-2063. PMC 7968070. PMID 32894316. doi:10.1007/s00125-020-05213-5. Consultado el 2 de octubre de 2022.
-
Cataldo Pignatelli; Francesco Campo; Alessia Neroni; Lorenzo Piemonti; Antonio Citro (2022). «Bioengineering the Vascularized Endocrine Pancreas: A Fine-Tuned Interplay Between Vascularization, Extracellular-Matrix-Based Scaffold Architecture, and Insulin-Producing Cells». Transpl Int. (REVISIÓN) 35: 10555. PMC 9452644. PMID 36090775. doi:10.3389/ti.2022.10555. Consultado el 16 de abril de 2023.
- Kravets V.; Dwulet J.M.; Schleicher W.E.; Hodson D.J.; Davis A.M.; Pyle L.; Piscopio R.A.; Sticco-Ivins M.; Benninger R. (2022). «Functional architecture of pancreatic islets identifies a population of first responder cells that drive the first-phase calcium response.». PLoS Biology (REVISIÓN) (en inglés) (Public Library of Science) 20 (9): e3001761. doi:10.1371/journal.pbio.3001761. Consultado el 26 de abril de 2023.
- «Tumores neuroendocrinos pancreáticos». Mayo Clinic. 2021.
- Marambio A.; Watkins G.; Jans J. (2012). «Gastrina: hormona de múltiples funciones». Rev Hosp Clín Univ Chile 23: 139-147.
- Sakata N.; Yoshimatsu G.; Kodama S. (2019). «Development and Characteristics of Pancreatic Epsilon Cells». Int J Mol Sci. (en inglés) 20 (8): 1867. PMC 6514973. PMID 31014006. doi:10.3390/ijms20081867. Consultado el 2 de octubre de 2022.
- Camillo Ricordi; Luca Inverardi; Norma S Kenyon; John Goss; Federico Bertuzzi; Rodolfo Alejandro (2005). «Requirements for success in clinical islet transplantation». Transplantation (en inglés) 79 (10): 1298-1300. doi:10.1097/01.tp.0000157275.64874.84.
- Martin Hertl (2020). «Trasplante de células de los islotes pancreáticos (trasplante de células de los islotes de Langerhans)». Manual MSD, Versión para profesionales. Consultado el 28 de setiembre de 2022.
- Cayabyab F.; Nih L.R.; Yoshihara E. (2021). «Advances in Pancreatic Islet Transplantation Sites for the Treatment of Diabetes». Frontiers in Endocrinology (REVISIÓN). Sec. Diabetes: Molecular Mechanisms (en inglés) (Frontiers (editorial)). Consultado el 2 de octubre de 2022..
- Claresa Levetan (2021). «Frederick Banting's observations leading to the potential for islet neogenesis without transplantation». Journal of Diabetes (REVISIÓN) 14 (2): 104-110. PMC 9060105. PMID 34967992. Consultado el 8 de abril de 2023.
- «¿Qué es tumor neuroendocrino?». American Cancer Society ACS. 2019.
- «Tumores neuroendocrinos pancreáticos». Mayo Clinic.
Enlaces externos
- What happens -Pancreas transplant. Página del Servicio Nacional de Salud (Reino Unido) NHS. En inglés.
- Islet cell transplants for type 1 diabetes
- Página del Instituto Nacional de Diabetes y Enfermedades Digestivas y Renales (NIDDK) del NIH. En inglés.
| Control de autoridades |
|
|---|
 Datos: Q728309
Datos: Q728309 Multimedia: Islets of Langerhans / Q728309
Multimedia: Islets of Langerhans / Q728309