Lophosoria quadripinnata
Lophosoria quadripinnata (J.F.Gmel.) C.Chr. es una especie de helecho que los análisis moleculares de ADN han ubicado en la familia de las dicksoniáceas, donde se la ubica en su propio género monotípico: Lophosoria.
| Lophosoria quadripinnata | ||
|---|---|---|
 | ||
| Taxonomía | ||
| Reino: | Plantae | |
| Filo: | Tracheophyta | |
| Clase: | Polypodiopsida | |
| Orden: | Cyatheales ("Helechos arborescentes") | |
| Familia: | Dicksoniaceae | |
| Género: |
Lophosoria J.Presl, 1847 | |
| Especie: |
Lophosoria quadripinnata (J.F.Gmel.) C.Chr., 1920 | |
Distribución

Se encuentra en el continente americano, desde Cuba y México hasta el sur de Chile y el extremo occidental de las provincias argentinas de Río Negro y de Neuquén.
En español se le denomina ampe (del mapudungun añpe) o palmilla, pero hay que tener en cuenta que hay varias especies de helechos a las que se llama «palmillas», que poseen frondes más o menos grandes y crecen en climas más bien fríos.
Es una planta de tamaño mediano y, pese a que el rizoma no forma estípite, está claramente emparentada con el resto de los helechos arborescentes debido a caracteres que aparentemente ya estaban en su ancestro común, como los pneumatodos, y el rizoma que cambió de la simetría dorsiventral típica de los demás helechos a una simetría radial típica de los helechos arborescentes. Sus frondes grandes y varias veces pinadas, con el pecíolo alzado adaxialmente, y los pelos en el rizoma y la parte inferior de los pecíolos también recuerdan a los helechos arborescentes.
Para identificar la especie, se usan la posición y las características de los soros que se encuentran en las hojas fértiles. El género ya existía en el Cretácico temprano en el sur de Gondwana, según restos fósiles encontrados en la Antártida. La especie es bien conocida como ornamental.
Descripción
- Introducción teórica en Terminología descriptiva de las plantas

Referencias:
n : generación haploide,
2n : generación diploide,
m! : mitosis,
M! : meiosis,
F! : fecundación
Lophosoria quadripinnata es una planta vascular con las dos generaciones alternadas, esporófito y gametófito, multicelulares e independientes; con esporas como unidad de dispersión y de resistencia. El gametófito es un «talo» (cuerpo sin organización) y el esporófito es un «cormo» (con raíz, vástago y sistema vascular). Debido a estas características, tradicionalmente se las agrupa con las pteridofitas. Poseen esporófitos con megafilos o «frondes» (Euphyllophyta).
El rizoma es masivo, con pelos, no formando estípite (no arborescente), pero es de simetría radial en lugar de horizontal, carácter que aparentemente nació en el ancestro de los helechos arborescentes. Las frondes son de tamaño grande, 2-3 pinadas, con pelos en la parte inferior de los pecíolos, y están alzadas en su parte abaxial, caracteres comunes en los cyatheales (clado de los helechos arborescentes). También están presentes los pneumatodos (líneas de ventilación discretas, o parches, dispuestos en el raquis, pecíolo o rizoma, que tienen la función de facilitar el intercambio gaseoso en los tejidos densos), que son característicos del clado de los helechos arborescentes.

Los soros, sin indusio, se ubican en la cara abaxial, sobre la vena de la última pina. El margen de la hoja puede estar reflejo pero no formando parte de un indusio, como en las demás dicksoniáceas. Los soros poseen numerosos pelos (llamados «parafisos filiformes») entre los esporangios. Las características y posición de los soros son «caracteres diferenciales» que se usan para identificar a la especie.
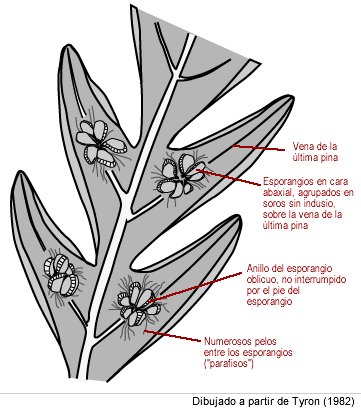 Detalle de cara abaxial de fronde fértil de Lophosoria quadripinnata Obsérvense los esporangios agrupados en soros sin indusio sobre la vena de la última pina, los soros con numerosos parafisos (pelos), los esporangios con anillo oblicuo no interrumpido por el pie del esporangio. Dibujado a partir de Tyron (1982).[1] |
Como en todas las polypodiópsidas, los esporangios son de tipo leptosporangiado (con pie, con cápsula de pared de una célula de espesor, con anillo de dehiscencia en la cápsula). Como en todos los helechos arborescentes, el anillo es oblicuo y completo, no interrumpido por el pie del esporangio.
Las esporas poseen marca trilete. Esta especie posee esporas con una morfología muy particular, con un cinturón ecuatorial («cíngulo») que hizo que durante mucho tiempo se ubicara a la especie en su propia familia, Lophosoriaceae.
La germinación del gametófito corresponde al tipo Cyathea, dando origen a filamentos cortos de dos a seis células en ambas variedades. El desarrollo protálico es de tipo Adiantum. Cuando son criados en laboratorio, los gametófitos de la variedad quadripinnata son siempre cordados, en cambio los gametófitos de la variedad contracta poseen tres morfos, dependiendo de la densidad de esporas a la que se desarrollen (Dyer, 1979):[2] A densidades bajas, son largamente espatulados con un meristema central y un ala ligeramente más desarrollada que la otra; a densidades medias son cordiformes, y a densidades altas son acintados y con una gran cantidad de anteridios.
El gametófito es protándrico (es hermafrodita, primero se desarrollan los anteridios que dan los anterozoides, y luego los arquegonios que dan las oosferas).
Número de cromosomas x = 65.
Sistemática
Los análisis moleculares de ADN han ubicado inequívocamente a la especie en las dicksoniáceas, pero la historia de su circunscripción es larga. En 1970 Pichi Sermolli la circunscribió en su propia familia, Lophosoriaceae Pichi Sermolli,[3] donde fue ubicada durante mucho tiempo. Al principio fue ubicada como hermana de las cyatheáceas, debido a caracteres como los soros abaxiales, o el patrón de germinación del gametófito. En la década de 1990 se ha llegado a hipotetizar una relación cercana con Metaxyaceae debido a caracteres del pecíolo y del tallo, pero análisis posteriores de morfología han sugerido que esas familias no estaban emparentadas (D. S. Conant, datos no publicados, citado en Wolf et al. 1999).[4] Por otro lado, los estudios de la ultraestructura (hechos con el microscopio electrónico de barrido) de las esporas (Gastony y Tyron, 1976)[5] y los estudios recientes del desarrollo del gametofito (Pérez-García et al. 1995)[6] han concluido que Lophosoria difiere en una cantidad significativa de caracteres de los demás helechos arborescentes, por lo que sus afinidades con el resto quedaban como incógnitas. Otros investigadores, como Kramer en Kubitzki (1990)[7] la anidaron dentro de las dicksoniáceas, junto con otros géneros que hoy en día se segregaron en otras familias de helechos arborescentes. A medida que se estudió a la especie molecularmente, fue cada vez más evidente su afinidad con las dicksoniáceas, el primer carácter que demuestra su afinidad es el número de cromosomas (igual al de Dicksonia), y los análisis moleculares de ADN hechos sobre la secuencia rbcL en 1999 también la anidaban dentro de esa familia (Wolf et al. 1999),[4] relación que fue confirmada en un extenso estudio molecular de los helechos arborescentes hecho por Korall et al. (2006,[8] sobre las secuencias atpA, atpB, rps4 y la ya mencionada). Debido a eso la especie fue ubicada en la familia de las dicksoniáceas, en su propio género monotípico, en el sistema de clasificación de monilophytas de Smith 2006.[9]
Circunscripción: la especie está conformada por al menos dos variedades, a veces puestas como especies del género:
- Lophosoria quadripinnata (J.F.Gmel.) C.Chr. var. quadripinnata (amplia distribución en los trópicos americanos, desde regiones húmedas y templadas de Sudamérica, entre los 1000 y 3000 m s. n. m.)
- Lophosoria quadripinnata var. contracta (Hieron.) R.M.Tryon & A.F.Tryon (encontrada en Ecuador, a 2900 msnm; y en la Isla Robinson Crusoe del archipiélago Juan Fernández en Chile).
Las esporas de la variedad contracta son ligeramente más grandes (70 × 75 µm) que las de la variedad quadripinnata (50 × 60 µm). Además, en la variedad quadripinnata los anteridios están formados por cinco células: dos basales, dos anulares y una pequeña opercular elíptica, mientras que en la variedad contracta están formados por tres células: una basal, una anular y una opercular (Stockey 1930).[10] También varían en la morfología del gametófito (ver caracteres).
También ha sido descrita Lophosoria quadripinnata var. quesadae A.Rojas en Costa Rica y Panamá (Rojas-Alvarado 1996, fue descrita como la especie Lophosoria quesadae).[11]
Evolución

Dettmann (1986)[12] ha remarcado que las esporas del género extinto Cyatheacidites se corresponden muy bien con las Lophosoria vivientes. La distribución de Cyatheacidites ha sido utilizada para inferir que Lophosoria estaba distribuida en la parte sur de Gondwana durante el Cretácico temprano y que, durante el Terciario, migró a Australia y Sudamérica (Dettmann 1986).[12]
Más recientemente, Cantrill (1998)[13] describió unas hojas fosilizadas en la capa del Cretácico temprano (más específicamente en el Aptiense), en la Antártida, que contiene esporas de Cyatheacidites. La preservación de las hojas y las esporas era tan buena que Cantrill pudo describir sus hallazgos en la categoría de especie, y la describió con el nombre de Lophosoria cupulata D.J.Cantrill (publicada como Lophosoria cupulatus), con el cambio de género, ya que él consideraba que Lophosoria quadripinnata (J.F.Gmel.) C.Chr. era la especie viviente más emparentada a la especie hallada.
Importancia económica
Se utiliza como planta ornamental, tanto viva en parques y jardines como cortada para servir como follaje acompañante de los ramos de flores.[14] Las plantas usadas de la segunda forma suelen extraerse de su ambiente natural, creando ciertos problemas locales de conservación.
En algunas ferias locales del sur de Chile se venden sus brotes, llamados perritos, para consumirlos en ensaladas.[14][15]
Referencias
- Tryon, R.M. & A.F. Tryon. 1982. Ferns and allied plants with special reference to tropical America. Springer-Verlag, Nueva York.
- Dyer, A.F. 1979. "The culture of fern gametophytes for experimental investigation." p. 253-305. In A.F. Dyer (ed.) The experimental biology of ferns. Academic Press, Londres.
- Pichi Sermolli R. E. G. 1970. «Fragmenta Pteridologiae.» -II. Webbia. 24: 699-722.
- Wolf et al. 1999. «Phylogenetic relationships of the enigmatic fern families Hymenophyllopsidaceae and Lophosoriaceae: evidences from rbcL nucleotide sequences.» Plant Syst. Evol. 219: 263-270
- Gastony G. J. y Tyron R. 1976. «Spore morphology in the Cyatheaceae". II. The genera Lophosoria, Metaxya, Sphaeropteris, Alsophila, and Nephelea.» Amer J. Bot. 63: 738-758.
- Pérez-García B., Fraile M. E., Mendoza A. 1995. "Desarrollo del gametófito del Lophosoria quadripinnata (Filicales: Lophosoriaceae)". Revista Biol. Trop. 43: 55-60.
- Kubitzki, K. 1990. «Pteridophytes and Gymnosperms.» en: K. V. Kramer. P. S. Green (Eds.) The families and genera of vascular plants. Vol 1. Springer-Verlag, Berlín, Alemania.
- Korall et al. 2006. "Tree ferns: Monophiletic groups and their relationships as revealed by four protein-coding plastid loci". Molecular phylogenetics and Evolution 39: 830-845.
- A. R. Smith, K. M. Pryer, E. Schuettpelz, P. Korall, H. Schneider, P. G. Wolf. 2006. [https://web.archive.org/web/20080226232147/http://www.pryerlab.net/publication/fichier749.pdf «A classification for extant ferns.» Taxonomy 55(3), 705-731.
- Stokey, A.G. 1930. "Prothallia of the Cyatheaceae." Bot. Gaz. (Crawfordsville) 90: 1-45.
- Rojas Alvarado A. F. 1996. "Aportes a la flora Pteridophyta costarricense". II. Taxones nuevos. Brenesia 45-46: 33-50
- Dettmann M. E. 1986. "Significance of the Cretaceous-Tertiary spore genus Cyatheacidites in tracing the origin and migration of Lophosoria (Filicopsida). Special Papers in Paleontology 35: 63-94.
- Cantrill D. J. 1998. "Early Cretaceous fern foliage from President Head, South Island, Antarctica. Alcheringa 22: 241-258
- Smith-Ramírez, Cecilia (1994). «Usos artesanales del bosque nativo. La extracción silenciosa». Revista Ambiente y Desarrollo X (2): 71-76. Archivado desde el original el 21 de octubre de 2007.
- Hoffmann Jacoby, Adriana (1999). El bosque chilote. Historia natural del archipiélago de Chiloé. Conservación y desarrollo sustentable de sus bosques y biodiversidad. Santiago: Defensores del Bosque Chileno. ISBN 956-7721-22-X.
| Control de autoridades |
|
|---|
 Datos: Q6678562
Datos: Q6678562 Multimedia: Lophosoria quadripinnata / Q6678562
Multimedia: Lophosoria quadripinnata / Q6678562 Especies: Lophosoria quadripinnata
Especies: Lophosoria quadripinnata