Mammuthus columbi
El mamut de Columbia o mamut columbino Mammuthus columbi) es una especie extinta de mamut que habitó en Norteamérica, extendiéndose desde el norte de Estados Unidos hasta Costa Rica en el sur durante la época del Pleistoceno. Fue una de las últimas especies del linaje de los mamuts que se originó a inicios del Plioceno con el hipotético antepasado común entre M. columbi y M. subplanifrons. El mamut columbino evolucionó a partir del mamut de la estepa, el cual ingresó a Norteamérica desde Asia hace unos 1,5 millones de años. El mamut pigmeo del archipiélago del Norte de California evolucionó a su vez que los mamuts columbino. El pariente vivo más cercano del mamut de Columbia y los demás mamuts es el elefante asiático.
| Mamut colombino | ||
|---|---|---|
| Rango temporal: 0,013 Ma - 0,011 Ma Pleistoceno | ||
 Esqueleto de un macho en el Museo George C. Page en Rancho La Brea | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Mammalia | |
| Orden: | Proboscidea | |
| Familia: | Elephantidae | |
| Género: | Mammuthus † | |
| Especie: |
M. columbi † (Falconer, 1857) | |
| Sinonimia | ||
Lista desplegable Elephas jacksoni Mather, 1838
Mammuthus jacksoni Mather, 1838 Parelephas jacksoni Mather, 1838 Elephas columbi Falconer, 1857 Euelephas columbi Falconer, 1857 Parelephas columbi Falconer, 1857 Mammuthus (Parelephas) columbi Falconer, 1857 Elephas primigenius columbii Falconer, 1857 Elephas texianus Owen, 1858 Elephas imperator Leidy, 1858 Mammuthus imperator Leidy, 1858 Euelephas imperator Leidy, 1858 Archidiskodon imperator Leidy, 1858 Mammuthus (Archidiskodon) imperator Leidy, 1858 Elephas jeffersonii Osborn, 1922 Parelephas jeffersonii Osborn, 1922 Mammuthus jeffersonii Osborn, 1922 Elephas roosevelti Hay, 1922 Parelephas roosevelti Hay, 1922 Elephas washingtonii Osborn, 1923 Parelephas washingtonii Osborn, 1923 Elephas maibeni Barbour, 1925 Elephas eellsi Hay, 1926 Elephas floridanus Osborn, 1929 Parelephas floridanus Osborn, 1929 Mammuthus floridanus Osborn, 1929 Archidiskodon sonoriensis Osborn, 1929 Parelephas progressus Osborn, 1942 | ||
Alcanzando 4 metros hasta la cruz y con un peso de 8 - 10 toneladas, el mamut de Columbia fue una de las mayores especies de mamut. Tenía grandes defensas ("colmillos") muy curvadas y cuatro molares, los cuales eran reemplazados seis veces durante la vida del animal. Es muy probable que usara sus defensas y su trompa como los elefantes modernos — para manipular objetos, luchar y alimentarse. Se han descubierto huesos, pelo, excrementos y contenidos estomacales, pero ningún individuo completo. El mamut columbino prefería áreas abiertas, como los paisajes tipo parkland, alimentándose de juncos, pastos y otras plantas. No llegó a vivir en las regiones árticas de Canadá, las cuales eran habitadas por el mamut lanudo. Los rangos de ambas especies pueden haberse superpuesto, y los análisis de biología molecular sugieren que incluso pudieron haberse hibridado. Varias localidades preservan los esqueletos de múltiples individuos de mamuts columbinos, ya sea porque murieron en un único incidente como una repentina inundación, o porque eran lugares que funcionaban como trampas naturales en las cuales los individuos se acumulaban con el tiempo.
Los mamuts columbinos coexistieron con los paleoamericanos, quienes los cazaban por su carne, usaban sus huesos para fabricar herramientas y además hicieron representaciones artísticas suyas. Los restos de mamuts se han encontrado en asociación con artefactos de la cultura Clovis; estos restos presentan la más alta frecuencia de explotación prehistórica de los proboscídeos como fuente de alimento de todo el mundo.[1] El mamut columbino desapareció al final del Pleistoceno hace unos 11 000 años, muy posiblemente como resultado de la caza por los humanos, y su extinción pudo verse acelerada por la pérdida de hábitat debido al cambio climático.[2]
Taxonomía
El mamut columbino fue descrito científicamente en 1857 por el naturalista escocés Hugh Falconer, quien nombró a la especie Elephas columbi en referencia a Cristóbal Colón. El animal fue puesto en conocimiento de Falconer en 1846 gracias a Charles Lyell, quien le envió fragmentos de molar hallados en 1838 durante la excavación del Brunswick Altamaha Canal en Georgia, en el sureste de Estados Unidos. Al mismo tiempo, fósiles similares de Norteamérica fueron atribuidos al mamut lanudo (llamado entonces Elephas primigenius). Falconer encontró que sus especímenes eran distintos, confirmando su conclusión al examinar su estructura interna y al estudiar molares adicionales de México. Aunque William Phipps Blake y Richard Owen creían que E. texianus era un nombre más apropiado para la especie, Falconer rechazó la denominación; además, sugirió que E. imperator[3] y E. jacksoni, los otros dos elefántidos norteamericanos descritos de molares, estaban basados en restos demasiado fragmentarios como para clasificarlos apropiadamente.[4] Material más completo que puede proceder de la misma cantera que el molar fragmentario holotipo de Falconer fue reportado en 2012, y puede ayudar a dar luz sobre este espécimen, debido a que se han generado dudas sobre qué tan adecuado es como holotipo.[5]
A inicios del siglo XX, la taxonomía de los elefántidos extintos se volvió más y más complicada. En 1942, se publicó la monografía póstuma de Henry Fairfield Osborn sobre los Proboscidea, en la cual él usó varios nombres de géneros y subgéneros que habían sido propuestos anteriormente para especies de elefántidos extintas, tales como Archidiskodon, Metarchidiskodon, Parelephas y Mammonteus. Osborn además usó nombres para muchas subespecies o "variedades" regionales o intermedias, y creó recombinaciones tales como Parelephas columbi felicis y Archidiskodon imperator maibeni.[6] La situación taxonómica fue simplificada por varios investigadores a partir de la década de 1970: todas las especies de mamut fueron situadas en el género Mammuthus, y muchas de las diferencias propuestas entre especies se interpretaron en cambio como variación intraespecífica.[7] En 2003, el paleontólogo Larry Agenbroad resumió el estado del arte sobre la taxonomía de los mamuts norteamericanos, y concluyó que varias especies había sido declaradas sinónimos más modernos, y que M. columbi (el mamut columbino) y M. exilis (el mamut pigmeo) eran las únicas especies de mamuts endémicas de América (ya que otras especies vivieron tanto allí como en Eurasia). La idea de que especies como M. imperator (el mamut imperial) y M. jeffersoni (el mamut de Jefferson) eran fases más primitivas o avanzadas de la evolución de la evolución del mamut columbino fue descartada, y se les considera como sinónimos. A pesar de estas conclusiones, Agenbroad advirtió que la taxonomía de los mamuts americanos aún no se ha resuelto completamente.[8]
Evolución
.jpg.webp)
Los primeros miembros conocidos de Proboscidea, el clado que incluye a los elefantes, aparecieron hace cerca de 55 millones de años alrededor del área del mar de Tetis. Los parientes vivos más cercanos de los Proboscidea son los sirenios (dugongos y manatíes) y los damanes (un orden de pequeños mamíferos herbívoros). La familia Elephantidae apareció hace seis millones de años en África, e incluye a los elefantes actuales y los mamuts. Entre los muchos clados hoy extintos, el mastodonte (Mammut), en contra de lo que puede indicar su nombre científico, es solo un pariente distante integrante de otra familia, los Mammutidae los cuales divergieron hace unos 25 millones de años antes de que evolucionaran los mamuts.[9] El elefante asiático (Elephas maximus) es el pariente vivo más cercano a los mamuts. El siguiente cladograma muestra la posición del mamut colombino entre otros proboscídeos, basándose en características del hueso hioides en el cuello:[10]
| ||||||||||||||||||||||||||||||||||

Debido a que se conocen muchos restos de cada especie de mamut de varias localidades, ha sido posible reconstruir la historia evolutiva del género a través de estudios morfológicos. Las especies de mamut pueden ser identificadas a partir del número de crestas en el esmalte (o placas laminares) en sus molares: las especies primitivas tenían pocas crestas, y el número se incrementó gradualmente a medida que las nuevas especies evolucionaban para comer vegetación más abrasiva. Las coronas de los dientes se volvieron más altas y los cráneos también ganaron altura a fin de acomodarlas. Al mismo tiempo, los cráneos se volvieron más cortos del frente hacia atrás para minimizar el peso de la cabeza.[11][12] Los cráneos cortos y altos del mamut lanudo y el columbino son la culminación de este proceso.[9]
Los primeros miembros conocidos del género Mammuthus son las especies africanas M. subplanifrons del Plioceno, y M. africanavus del Pleistoceno. Se cree que el primero es el ancestro de las formas posteriores. Los mamuts entraron a Europa hace unos 3 millones de años. La forma europea más antigua ha sido nombrada M. rumanus; y se extendió a partir de Europa hasta llegar a China. Solo sus molares son conocidos, los cuales muestran que tenía 8–10 crestas en el esmalte. Una población desarrolló 12–14 crestas, divergiendo y reemplazando a la forma anterior, convirtiéndose en M. meridionalis hace unos 2–1.7 millones de años. A su vez, esta especie fue reemplazada por el mamut de la estepa (M. trogontherii) con 18–20 crestas, el cual evolucionó en Asia oriental hace unos 2-1.5 millones de años.[11] El mamut columbino evolucionó a partir de una población de M. trogontherii (anteriormente se había considerado que había sido M. meridionalis) la cual cruzó el estrecho de Bering e ingresó a Norteamérica hace unos 1.5 millones de años; este retuvo un número similar de crestas molares. Los mamuts derivados de M. trogontherii evolucionaron molares con 26 crestas hace 400,000 años en Siberia y se convertirían en el mamut lanudo (M. primigenius). Los mamuts lanudos entraron a Norteamérica hace unos 100,000 años.[9]

Una población de mamuts columbinos que vivió hace entre 80,000 y 13,000 años en el archipiélago del Norte de California, a 10 kilómetros de distancia del continente, evolucionó a menos de la mitad del tamaño corporal de los mamuts columbinos continentales. Por tanto, se considera que representan una especie distinta, M. exilis, el mamut pigmeo (o como una subespecie, M. columbi exilis). Estos mamuts presumiblemente alcanzaron las islas al nadar hasta allí cuando el nivel del mar era más bajo, e iniciaron un proceso de enanismo insular debido a la escasez de comida en áreas pequeñas. Los huesos de especímenes grandes también se han hallado en las islas, pero se desconoce si estos representan fases del proceso de enanismo, o llegadas posteriores de mamuts columbinos.[10][13][14]
Un estudio publicado en 2011 sobre el genoma mitocondrial completo (heredado a través de la línea materna) mostró que dos mamuts columbinos examinados, incluyendo el "mamut de Huntington" que es morfológicamente típico, se agrupaban dentro de un subclado de mamuts lanudos. Esto sugiere que ambas poblaciones se hibridaron y produjeron descendencia fértil. Una posible explicación es una introgresión de un haplogrupo de los mamuts lanudos a los columbinos, o viceversa. Una situación similar ha sido documentada en las especies modernas de elefantes africanos (Loxodonta), el elefante africano de sabana (L. africana) y el elefante africano de bosque (L. cyclotis). Los autores de este estudio también sugieren que la forma norteamericana anteriormente denominada como M. jeffersonii puede haber sido un híbrido de ambas especies, ya que es aparentemente un intermedio morfológico.[15] Estos hallazgos fueron inesperados para los científicos, y se tendrá que analizar el ADN nuclear y más especímenes para dar claridad a la situación.[16]
Un estudio publicado en 2015 sobre los molares de mamuts de América del Norte confirmó que M. columbi evolucionó a partir de la especie euroasiática M. trogontherii, y no de M. meridionalis como se había sugerido anteriormente, y señaló que M. columbi y M. trogontherii eran morfológicamente muy parecidos. El estudio también sugirió que los animales situados en el rango en el cual se superponían M. columbi y M. primigenius formaban una metapoblación de híbridos con morfología variable.[17] En una entrevista, uno de los autores del estudio fue más allá y afirmó que M. columbi y M. trogontherii podrían en realidad ser la misma especie que habitó en continentes diferentes, y que el primero podría ser el sinónimo más antiguo del primero (es decir, ser el nombre válido), ya que fue nombrado con anterioridad. Él también señaló que dado que los científicos europeos suelen referirse a los especímenes del Viejo Mundo como M. trogontherii, la aceptación del cambio de nombre probablemente no será inminente.[18]
Descripción


El mamut columbino alcanzaba 4 metros de altura en la cruz, y pesaba entre 8 y 10 toneladas. Tenía el mismo tamaño que las especies anteriores de mamuts M. meridionalis y M. trogontherii, y era mayor que el actual elefante africano y que el mamut lanudo, los cuales alcanzan entre 2.7 a 3.4 metros. Los machos por lo general eran más grandes y más robustos. La mejor indicación del sexo es la cintura pélvica, dado que la abertura que funciona como canal de parto es siempre más ancha en las hembras que en los machos. Como otros mamuts, el columbino tenía una cabeza alta con un único domo y una espalda decreciente con una alta joroba en los hombros; esta forma resultaba de los procesos espinosos (protrusiones) de las vértebras de la espalda que reducían su altura de adelante para atrás Los juveniles, por otro lado, tenían espaldas convexas como los elefantes asiáticos.[19] Otros rasgos del esqueleto incluyen un rostro corto y alto, una sínfisis mandibular redondeada (el punto en que se unen ambas mitades de la mandíbula) y la apófisis coronoides mandibular (la protrusión superior del hueso de la mandíbula) extendiéndose sobre las superficies de los molares.[20]

Aparte de su gran tamaño y molares más primitivos, el mamut columbino se diferenciaba del mamut lanudo por su sínfisis mandibular más orientada hacia abajo; los alvéolos (cuencas de los dientes) de los colmillos se dirigían más hacia los lados, alejándose del medio.[15] Su cola era de longitud intermedia entre la de los elefantes modernos y el mamut lanudo. Dado que no se han encontrado tejidos blandos de los mamuts columbinos, se sabe mucho menos sobre su apariencia que en el caso del mamut lanudo. Vivió en hábitats más cálidos que el mamut lanudo, por lo que posiblemente carecía de muchas de las adaptaciones vistas en esa especie. Se ha descubierto pelo que se piensa proviene de un mamut columbinos en la cueva Bechan en Utah, en donde también se ha hallado excremento de mamut. Algunos de estos pelos son ásperos, e idénticos a los que se sabe pertenecían al mamut lanudo; sin embargo, dado que su localización se halla mucho más al sur es improbable que sea pelo de la especie lanuda. La distribución y densidad del pelo en el animal vivo es desconocida, pero probablemente era mucho menos denso que el del mamut lanudo, debido al hábitat más cálido.[9][19]
Dentadura

Los mamuts columbinos tenían colmillos muy largos (en realidad, eran dientes incisivos modificados), los cuales eran más curvados que los de los elefantes modernos. El colmillo más grande conocido de un mamut, de 4.9 metros de longitud, perteneció a un mamut columbino y otros estaban en el rango de 3.5 a 4.1 metros de largo. Los colmillos del mamut columbino generalmente no eran mayores que los de los mamuts lanudos, los cuales alcanzaban 4.2 metros. Los colmillos de las hembras eran mucho más pequeños y delgados. Cerca de un cuarto de la longitud de los colmillos se encontraba dentro de los alvéolos; a partir de ahí crecían de forma espiralada y en direcciones opuestas, curvándose hasta que las puntas terminaban encontrándose, y a veces entrecruzándose. La mayor parte de su peso se habría concentrado cerca del cráneo, con menos torque del que le hubieran dado unos colmillos rectos. Los colmillos además eran usualmente asimétricos, con una considerable variación; algunos colmillos se curvaban hacia abajo, en lugar de hacia los lados, o eran más cortos debido a que se rompían. Los colmillos del mamut columbino eran generalmente menos espiralados que los del mamut lanudo. A la edad de seis meses las crías desarrollaban colmillos de leche de unos pocos centímetros de largo, los cuales eran reemplazados por colmillos permanentes un año después. Un crecimiento anual de los colmillos de 2.5 – 15 cm. continuaba a lo largo de la vida del animal, disminuyendo después de que se alcanzara la adultez.[19][21]
Los mamuts columbinos tenían cuatro dientes molares funcionales al mismo tiempo, dos en el maxilar y dos en la mandíbula. Cerca de 23 cm de la corona se encontraba dentro de la mandíbula, mientras que 2,5 cm sobresalían. La corona iba sobresaliendo hacia adelante y arriba a medida que se desgastaba, de forma comparable a una cinta transportadora. Los dientes tenían crestas separadas de esmalte, el cual estaba cubierto con "prismas" dirigidos hacia la superficie masticatoria. Resistentes al desgaste, estos se sostenían juntos gracias al cemento dental y la dentina. Los molares de un mamut se reemplazaban cinco veces durante la vida del animal. Los primeros molares eran de un tamaño similar a los de los humanos, de 1,3 cm; los terceros eran de 15 cm de largo, y lo sextos eran de 30 cm de largo y pesaban 1,8 kg. Los molares se hacían mayores, conteniendo más crestas con cada reemplazo, y el número de placas variaba entre individuos.[19][22] El crecimiento de una cresta de 18 cm. tomaba cerca de 10.6 años.[23]
Paleobiología

Al igual que en los elefantes modernos, la sensible y musculosa trompa de un mamut era un órgano con muchas funciones. Era usada para manipular objetos e interactuar socialmente. Aunque los mamuts adultos sanos podían defenderse de los depredadores con su tamaño, su trompa y sus colmillos, los juveniles y los adultos debilitados eran vulnerables a los cazadores en grupo como los lobos o los grandes félidos. El ser humano sin embargo, sí era capaz de matar tanto a adultos sanos como enfermos, mediante el uso de lanzas y probablemente también de flechas envenadas[24] como las que usan aún hoy los furtivos en África para matar machos adultos de gran tamaño.[25] Huesos de mamuts columbinos juveniles, acumulados por Homotherium (el félido dientes de cimitarra), han sido hallados en la caverna Friesenhahn en Texas. Los colmillos pueden haberse usado en combates intraespecíficos por territorio y para exhibirse, atrayendo a las hembras e intimidando a los posibles rivales. En lo que se refiere al comportamiento, el descubrimiento en Nebraska de dos mamuts columbinos, muertos los dos con los colmillos enganchados, demuestra que estos animales combatían entre sí. Los mamuts pudieron usar sus colmillos como armas al empujarse, golpearse o estrellarlos, y eran usados en competiciones de empuje al juntarlos, lo que a veces terminaba en su rotura. La curvatura de los colmillos impedía que fueran usados para apuñalar. En Goat Rock Beach en el Parque Estatal Costa de Sonoma, afloramientos de esquisto azul y pedernal (apodados "rocas de mamuts") dan indicios de haber sido usados por los mamuts columbinos o los mastodontes para rascarse. Las rocas tienen áreas pulidas a 3 - 4 metros sobre el suelo, principalmente cerca de sus bordes, y son similares a las rocas africanas que son usadas por los elefantes y otros herbívoros para quitarse el barro y los parásitos. Rocas parecidas se han encontrado en Hueco Tanks, Texas, y en Cornudas Mountain en Nuevo México.[26][27][28]

Las acumulaciones de restos de elefantes modernos han sido denominadas como "cementerios de elefantes", debido a que se creía erróneamente que eran lugares a los que acudían los elefantes viejos que iban a morir. Se han encontrado acumulaciones parecidas de huesos de mamut; se piensa que son el resultado de individuos que murieron en ríos o cerca de ellos a través de miles de años y sus huesos fueron acumulados por el agua (como en el caso del río Acuilla en Florida), o de animales atrapados en el fango. Se piensa que algunas acumulaciones son restos de manadas que murieron al mismo tiempo, quizás debido a una inundación. Ocasionalmente se preservaron mamuts columbinos en depósitos volcánicos como los de Tocuila, en Texcoco, México, en donde un lahar volcánico cubrió al menos a siete individuos hace 12,500 años. Se desconoce cuantos mamuts vivían en un mismo lugar en un momento dado, pero es probable que el número variara según la estación y el ciclo vital. Los elefantes modernos pueden formar grandes rebaños, a veces compuestos por múltiples grupos familiares, y estos rebaños pueden incluir miles de animales migrando juntos. Los mamuts pudieron formar grandes manadas más frecuentemente que los elefantes modernos, dado que los animales viviendo en áreas abiertas tienen mayor oportunidad de hacerlo que aquellos que viven en áreas más forestadas. Aunque no es claro hasta que punto migraban los mamuts columbinos, un análisis isotópico de Blackwater Draw en Nuevo México indicó que pasaban parte del año en las Montañas Rocosas, a 200 km de distancia. El estudio de los anillos de sus colmillos puede ayudar en futuras investigaciones sobre la migración de los mamuts.[19][29][30]

Como los elefantes modernos, los mamuts columbinos eran probablemente sociales y vivían en grupos familiares matriarcales (liderados por hembras); la mayor parte de su comportamiento social debió ser similar al de los elefantes modernos. Esto es apoyado por las colecciones fósiles como las del yacimiento de Dent (en inglés: Dent Site) en Colorado y el Monumento Nacional del Mamut, en Waco (Waco Mammoth National Monument), donde se han hallado grupos compuestos enteramente por mamuts coumbinos hembras y ejemplares jóvenes (lo que implica grupos liderados por hembras). En el piso más reciente hay un total de 22 esqueletos, con 15 individuos interpretados como una manada de hembras y juveniles. Se propuso originalmente que murieron durante una inundación repentina, y que la disposición de algunos de los esqueletos sugeriría que las hembras habrían formado un círculo defensivo alrededor de las crías. En 2016, se sugirió en cambio que el grupo murió durante una sequía cerca de un abrevadero que estaba disminuyendo; las marcas de carroñeo en los huesos descartan que se hayan enterrado rápidamente, y la ausencia de crías y de una gran diversidad de otras especies animales halladas juntas en el yacimiento apoyan esta interpretación. Otro grupo, compuesto por un macho y seis hembras, también fue hallado en el mismo yacimiento; aunque ambos grupos murieron hace entre 64,000 y 73,000 años, se ignora si murieron en un mismo evento. En el yacimiento Clovis de Murray Springs, en Arizona, donde se han excavado varios esqueletos de mamuts columbinos, se halló un rastro de huellas fosilizadas similar al dejado por los elefantes modernos que conduce a uno de los esqueletos. El mamut puede haber dejado las pisadas antes de morir, u otro individuo puede haberse aproximado al animal muerto — de forma similar a como los elefantes modernos guardan a sus parientes muertos durante varios días.[19][29][31]
Trampas naturales

Muchos especímenes se acumularon además en trampas naturales, tales como sumideros o dolina y pozos de brea. El Hot Springs Mammoth Site en Dakota del Sur es un sumidero de 26,000 años de antigüedad, de aproximadamente 40 metros de largo que estuvo activo entre 300 a 700 años antes de llenarse con sedimentos. Este yacimiento es un caso opuesto al de Waco; casi todos, de los más de 55 esqueletos descubiertos —nuevos restos se excavan cada año—corresponden a machos, y se acumularon a través del tiempo, y no en un único evento. Se asume que como los elefantes machos modernos, los machos de mamut vivían mayormente en soledad, y eran más aventureros que las hembras (especialmente los machos más jóvenes) y por tanto más susceptibles de terminar en situaciones peligrosas. Los mamuts pueden haber sido atraídos a los pozos por el agua cálida o la vegetación en sus orillas, resbalando y ahogándose, o muriéndose de hambre. Los estudios de isótopos de los anillos de crecimiento han mostrado que muchos de estos mamuts murieron durante la primavera y el verano, lo cual podría correlacionarse con la vegetación cerca del pozo. Un individuo, apodado "Murray" yace de costado, y probablemente murió en esa postura mientras luchaba por liberarse. Las profundas huellas de mamuts tratando de salir del pozo de fango pueden ser vistas en las secciones excavadas en vertical en este yacimiento.[19][29]

Desde el inicio del siglo XX, las excavaciones en Rancho La Brea en Los Ángeles han proporcionado 100 toneladas de fósiles de 600 especies de flora y fauna, incluyendo varios mamuts columbinos. La mayor parte de estos fósiles son restos de animales que quedaron atrapados en charcos de asfalto las cuales se filtraban hasta la superficie de los pozos, hace entre 40,000 a 11,500 años. El polvo y las hojas pudieron haber escondido al asfalto líquido, lo cual atraparía a los animales incautos. Los animales atascados murieron de hambre y cansancio; y sus cuerpos atraían a los depredadores, que a su vez quedaban atrapados. El registro fósil de los pozos de brea se compone mayormente de los restos de depredadores como los grandes cánidos y félidos. Los fósiles de diferentes animales se hallan atrapados juntos cuando son excavados de los pozos.[29] Los pozos de brea no preservan los tejidos blandos, y un estudio de 2014 concluyó que el asfalto puede degradar el ADN de los animales atascados en este, tras un intento de extraer ADN de un mamut columbino.[32]
Dieta

Un mamut columbino adulto pudo haber necesitado más de 180 kilogramos de comida al día, y pudo haber comido durante veinte horas del día. Los mamuts masticaban su comida usando sus poderosos músculos mandibulares para mover la mandíbula hacia adelante y cerca de la abertura de la boca, para luego retraerla mientras la abría; las agudas crestas de esmalte por tanto trituraban al avanzar una sobre otra entre sí, moliendo la comida. Las crestas eran resistentes al desgaste, permitiéndole al animal masticar grandes cantidades de comida que contuviera grava. La trompa pudo haber sido usada para tomar grandes manojos de pastos, escoger brotes y flores o removiendo hojas y ramas de árboles y arbustos, y los colmillos eran usados para cavar en busca de plantas y remover corteza de los árboles. Esto es indicado en los colmillos preservados por las secciones aplanadas y pulidas de la superficie que podían alcanzar el suelo.[19] Estudios de isótopos de mamuts columbino de México y Estados Unidos han mostrado que su dieta variaba según su geografía, consistiendo en una mezcla de plantas C3 (como varias plantas) y plantas C4 (como los pastos), y que por tanto no se restringían solo a pastorear o a ramonear.[33]

Los contenidos estomacales de mamuts columbinos son raros, dado que no se han hallado ejemplares momificados, pero se han descubierto restos de plantas entre la pelvis y las costillas del "mamut de Huntington" cuando este fue excavado en Utah. El análisis microscópico mostró que estos restos masticados consistían de juncos, pastos, ramitas y agujas de abetos, roble y arce.[34] Una gran cantidad de excremento de mamut fue hallada en dos cavernas de Utah. Las condiciones secas y la temperatura estable de la Cueva Bechan (bechan es el término navajo para "heces grandes") ha preservado excremento de elefántido de 16,000 a 13,500 años de antigüedad, muy probablemente de mamuts columbinos. El excremento consistía en un 95 por ciento de pasto y de, y varía de 0 a 25 por ciento de plantas leñosas entre el bolo alimenticio, incluyendo plantas de sal, artemisias, abedul rojo y la pícea azul. Esta dieta es parecida a la documentada para el mamut lanudo, aunque el ramoneo parece haber sido más importante para el mamut columbino. La cubierta de excremento mide 41 centímetros de grosor, y tiene un volumen de 227 m³, con grandes bolos de hasta 20 centímetros de diámetro. El excremento de Bechan pudo haber sido producido por un pequeño grupo de mamuts durante un periodo relativamente corto, dado que los elefantes africanos adultos producen un promedio de 11 kilogramos cada dos horas y hasta 90 – 135 kilogramos cada día.[19][35]
Se ha propuesto que las grandes frutas de plantas norteamericanas como el naranjo de los osages, el cafetero de Kentucky y la acacia de tres espinas evolucionaron en conjunto con la ahora extinta megafauna americana, como los mamuts y otros proboscídeos, dado que ninguno de los herbívoros nativos modernos son capaces de ingerir estas frutas y dispersar sus semillas. Los bovinos y caballos introducidos por los europeos acabaron por asumir ese rol ecológico.[36][37]
Ciclo vital
_-_Natural_History_Museum_of_Utah_-_DSC07258.JPG.webp)
Se piensa que la esperanza de vida del mamut columbino sería de alrededor de 80 años. La esperanza de vida de un mamífero se relaciona con sus tamaño; los mamuts columbinos eran más grandes que los elefantes modernos, los cuales viven alrededor de 60 años. La edad de un mamut puede determinarse aproximadamente al contar los anillos de crecimiento de sus colmillos cuando son vistos en corte transversal. Sin embargo, este método no revela los primeros años de un mamut; el crecimiento inicial se representa en las puntas de colmillos, los cuales usualmente son los primeros en desgastarse. En el colmillo restante cada línea grande representa un año, con líneas diarias y semanales en el medio. Las bandas oscuras correspondan al verano, lo que hace posible determinar la estación en la que murió un mamut. El crecimiento del colmillo se ralentizaba cuando se hacía más difícil alimentarse, como ocurría durante una enfermedad o cuando un mamut macho era expulsado del rebaño (los elefantes machos viven en sus rebaños hasta que cumplen diez años).[19]
Los mamuts continuaban creciendo durante la adultez, como ocurre en otros elefántidos. Los machos crecían hasta la edad de 40, y las hembras hasta que cumplían 25. Los mamuts pudieron tener períodos de gestación de 21–22 meses, como los elefantes modernos. Los mamuts columbinos tenían seis juegos de molares en el curso de sus vidas. Al cumplir 6–12 meses erupcionaba el segundo juego, y el primero terminaba de caer a los 18 meses de edad. El tercer conjunto duraba diez años, y este proceso se repetía hasta que el sexto emergía a los 30 años de edad. Cuando este último juego de molares se gastaba el animal perdía la capacidad de masticar, y moriría de hambre.[19]
Casi todas las vértebras del "mamut de Huntington", un espécimen de edad muy avanzada, fueron deformadas por una enfermedad artrítica, y cuatro de sus vértebras lumbares estaban fusionadas; algunos huesos también indican infección bacteriana, como la osteomielitis.[19] La condición de los huesos sugiere que el espécimen murió de vejez y malnutrición.[34]
Distribución y hábitat

Los mamuts columbinos habitaron la mitad sur de Norteamérica, extendiéndose desde el norte de Estados Unidos, atravesando México y llegando tan al sur como Costa Rica. El espécimen costarricense, un molar, fue reportado en 1963 pero se ha perdido desde entonces.[38] El ambiente en estas áreas tenía hábitats más variados que los que frecuentaba el mamut lanudo en el norte. Algunas áreas estaban cubiertas por pastos, plantas herbáceas, árboles y arbustos; su composición variaba de región a región, e incluía pastizales, sabanas y parklands. También había áreas de bosques; aunque los mamuts evitaban los bosques, los claros podían proporcionarles pastos y hierbas.[19]
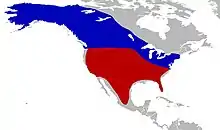
El mamut columbino compartía su hábitat con otros mamíferos extintos del Pleistoceno, como Glyptodon, Smilodon, los perezosos terrestres, Camelops y el mastodonte americano), caballos y bisontes. No habitó en la zona ártica canadiense, la cual era ocupada por los mamuts lanudos. Se han hallado fósiles de ambas especies de mamuts en un mismo lugar pero únicamente en algunas áreas de Norteamérica en la que su distribución se superponía, incluyendo a Hot Springs Site. Se desconoce si ambas especies eran simpátricas y vivían allí simultáneamente, o si los mamuts lanudos llegaban a las áreas del sur cuando las poblaciones de mamuts columbinos se ausentaban.[9] El mamut columbino además coexistió con otros dos proboscídeos extintos, Stegomastodon mirificus y Cuvieronius tropicus en yacimientos de Texas y Nuevo México durante el inicio del Irvingtoniense.[39]
Relación con los humanos

Los humanos penetraron en el continente americano a través de Beringia, y hay pruebas de que interactuaban con los mamuts columbinos. Se han encontrado por ejemplo herramientas hechas con restos de mamuts en varios yacimientos norteamericanos. En Tocuila, México, los huesos de mamut fueron usados hace 13 000 años para producir lascas y núcleos líticos. En el yacimiento de Lange-Ferguson, en Dakota del Sur, se hallaron los restos de dos mamuts de hace 12.800 años junto con hendidores y cantos tallados elaborados con un omóplato de mamut; los cantos se habían usado para carnear a los mamuts. En el mismo yacimiento se encontró una hoja lítica fabricada a partir de un hueso largo de mamut e insertada en las vértebras de un mamut. En Murray Springs, los arqueólogos descubrieron un objeto de 13.100 años de antigüedad hecho de un fémur de mamut; se cree que este objeto sería una llave de eje, un herramienta para fortalecer la madera y el hueso para elaborar astas de lanzas (los inuit usan herramientas similares).[40][41]
Los restos más antiguos que sugieren el consumo de carne de mamut datan de 21.500 a 22.000 años.[42] Los paleoamericanos de la cultura Clovis, la cual surgió unos 7.000 años después, pueden haber sido los primeros humanos en cazar a los mamuts sistemáticamente. Se piensa que este pueblo cazaba mamuts columbinos con lanzas dotadas de puntas clovis, las cuales se arrojaban o empujaban. Aunque las puntas clovis se han encontrado con restos de mamuts columbinos en varios yacimientos, los paleontólogos no se ponen de acuerdo sobre si estos hallazgos representan cacerías, consumo de mamuts ya muertos o son simplemente una coincidencia. Una hembra de mamut en el Naco-Mammoth Kill Site en Arizona, hallada junto con ocho puntas clovis cerca de su cráneo, omóplato, costillas y otros huesos, es considerada como la prueba más convincente de cacería. En los experimentos modernos, las réplicas de lanzas prehistóricas fueron capaces de penetrar las cajas torácicas de elefantes africanos y además reusarse, causando poco daño a las puntas.[43][44]
Otros yacimientos muestran pruebas más circunstanciales de caza de mamuts, como huesos apilados que tienen marcas de carnicería. Algunos de estos yacimientos no están asociados cercanamente con puntas clovis. Lehner Mammoth-Kill Site en el yacimiento de Dent, en donde se hallaron numerosos mamuts jóvenes y adultos con marcas de carnicería y en asociación con puntas clovis, fue interpretado anteriormente como la matanza de rebaños enteros por los cazadores clovis. Sin embargo, los estudios de isótopos han mostrado que estas acumulaciones representan muertes individuales en diferentes estaciones del año, y por lo tanto no son rebaños muertos en un único evento. Muchos otros yacimientos de huesos con marcas de carnicería también pueden representar acumulaciones a través del tiempo, y por lo tanto como prueba de caza a gran escala resultan ambiguos.[43][45]

Los petroglifos hallados en la meseta del Colorado pueden representar mamuts columbinos o mastodontes.[40] Un fragmento de hueso de 13 000 años de antigüedad de Vero Beach, Florida es el más temprano ejemplo de arte en América, y muestra a un mamut o mastodonte tallado.[46] Se piensa que los petroglifos de hace 13 000-11 000 años del río San Juan en Utah incluyen representaciones de dos mamuts columbinos; las cabezas en forma de cúpula de los mamuts los distinguen de los mastodontes. También son mostrados con dos "dedos" en la punta de sus trompas, un rasgo observado en las representaciones europeas de los mamuts. Los colmillos son muy cortos, lo cual puede indicar que son hembras. Una talla de un bisonte (posiblemente de la especie extinta Bison antiquus) esta sobrepuesto a uno de los mamuts y puede ser una adición posterior. Otras posibles representaciones de mamuts columbinos han sido descartadas como malinterpretaciones o fraudes.[47]
El mamut columbino es el fósil estatal de Washington.[48] El fósil estatal de Nebraska es "Archie", un espécimen de mamut columbino hallado en ese estado en 1922. "Archie" se encuentra expuesto en el Museo Estatal de la Universidad de Nebraska en Lincoln, y es el mayor espécimen de un mamut montado en Estados Unidos.[49]
Extinción

Los mamuts columbinos y los lanudos desaparecieron conjuntamente entre el final del Pleistoceno y el inicio del Holoceno, junto con la mayor parte de la megafauna de la época.[43] Los restos de mamuts más recientes que han sido datados tienen alrededor de 11,000 años.[50][51][52] Esta extinción formó parte de la extinción del Cuaternario, la cual comenzó hace 40,000 años y tuvo su auge entre 14,000 a 11,500 años antes del presente.[43] Los científicos no saben si estas extinciones sucedieron de manera abrupta o a lo largo de un período relativamente extenso.[50][53] Durante este período, 40 especies de mamíferos desaparecieron de Norteamérica, casi todas con un peso superior a los 40 kilogramos; la extinción de los mamuts no puede, por tanto, explicarse aisladamente.[43]
Aunque los científicos siguen divididos acerca de si fue la caza, los cambios climáticos, o una combinación de ambas, lo que llevó a la extinción a los mamuts columbinos, hoy en día la hipótesis de la caza es la que tiene mayor consistencia, sin desechar el posible efecto sinérgico del cambio climático.
De acuerdo con la hipótesis del cambio climático, el clima más cálido condujo a la reducción del hábitat apropiado para los mamuts, el cual habría cambiado de praderas a bosques, pastizales y semidesiertos, con una vegetación menos diversa (aunque actualmente se cree que esto pudo ser consecuencia, y no causa, de la extinción de la megafauna[2]). La hipótesis del cambio climático tiene además varios puntos débiles que están siendo señalados por numerosos científicos en los últimos años,[2][54] destacando tres hechos que esta hipótesis no puede explicar:
• La megafauna terrestre ha desaparecido a lo largo de los últimos 55.000 años de la mayor parte del planeta, pero lo ha hecho antes en unos continentes que en otros, y antes en los continentes que en sus islas, con diferencias contrastadas de varios miles de años en el caso de los mamuts siberianos, de los perozosos gigantes antillanos, de los elefantes europeos y de los marsupiales gigantes de Tasmania entre los continentes y sus resepctivas islas. Esto no cuadra con un cambio climático, de hecho se contradice con lo esperado para este tipo de evento: a menor espacio para migrar a climas más favorables, y a menor tamaño de las poblaciones insulares, y por tanto menor pool genético para adaptarse a los cambios, mayor es la probabilidad de extinción de las especies.
• El segundo hecho comprobado en las extinciones masivas del Cuaternario, es que las extinciones afectan de forma muy específica a la megafauna, compuesta por animales capaces de recorrer enormes distancias a lo largo de su vida, y más aún en el caso de los dos continentes americanos (que habrían sido los más afectados por el cambio de ciclo glacial de acuerdo a esta hipótesis), porque las cordilleras americanas van de norte a sur y permiten perfectamente la migración (eso explica, por ejemplo, que muchos géneros de plantas que se extinguieron en Europa, con sus cordilleras de oeste a este impidiéndoles el paso, sobrevivieran en Norteamérica y Asia oriental).
• Un cambio climático natural dura miles de años y es gradual, por lo que difícilmente puede explicar por sí solo la capacidad de extinguir al 73% de las especies de megafauna en Norteamérica cuando habrían tenido tanto tiempo para emigrar.
La hipótesis de la sobrecaza, por su parte, atribuye su extinción a la depredación por parte de los humanos.[43] Esta idea fue propuesta inicialmente por Paul S. Martin en 1967,[55] cuando expuso su hipótesis basándose en que había una correlación cada vez más evidente entre la expansión del Homo sapiens por el planeta y los eventos de extinción masiva que afecvtaban de forma casi exclusiva a la megafauna. Los investigadores a lo largo del SXX y principios del XXI sobre este asunto ha variado sobre sus conclusiones. Un estudio de 2002 concluyó que el registro arqueológico no apoyaba la hipótesis de la sobrecaza, dado que de 76 yacimientos clovis examinados, solo 14 (12 con restos de mamut y dos con restos de mastodonte) libraban pruebas sólidas de cacería.[56] La explicación a esta aparente ausencia de restos, llamada la "crítica de la asociación" (Meltzer, 1986) la detalla por ejemplo el ecólogo Santos Fernández en 2016: el periodo de convivencia de mamuts y seres humanos, hasta que los mamuts se extinguieron, constituye una parte muy pequeña del rango geológico de los registros de mamut.[57] Ambas especies habrían convivido alrededor de veinte siglos en América del Norte , pero en una localidad concreta la convivencia fue mucho más corta aún[2] Por lo tanto, es muy complicado encontrar restos arqueológicos que sean precisamente de las pocas décadas o siglos en que los mamuts y los seres humanos coexistieron en esa localidad dada. En cualquier otro caso, sólo se encontraría mamuts sin seres humanos (antes de la coexistencia) o humanos sin mamuts (después de la coexistencia). El patrón esperado es que una pequeña fracción de yacimientos que contienen megafauna tendría restos arqueológicos, un 18% en el caso del estudio de 2002. Así, un nuevo estudio de 2008 halló que el mismo registro de los clovis indicaba en realidad que tenían la frecuencia de explotación prehistórica de proboscídeos como fuente de alimento más alta de todo el mundo, lo que fortalece claramente la hipótesis de la sobrecaza.[1]
Recientes artículos y estudios científicos publicados en esta última década apoyan cada vez con mayor claridad que el hombre debió ser la causa principal de la extinción de la mayor parte de la megafauna desaparecida durante el cuaternario en el planeta.[2][58][54] Por un lado el Homo sapiens era capaz de matar ejemplares adultos de megafauna utilizando lanzas y flechas, en el caso de los paleomaericanos incluso con puntas envenenadas,[24] armas como las que utilizan aún hoy los furtivos para abatir grandes machos adultos de elefante de sabana en Kenia.[25] Por otra parte los mamíferos grandes son generalmente mucho más vulnerables que los pequeños a una caza excesiva debido al tamaño reducido de sus poblaciones y sus bajas tasas de reproducción.[43] Y por último a esto habría que añadir que las especies americanas no coevolucionaron con el hombre, por lo que es bastante posible que presentaran ingenuidad ecológica[2] (o ingenuidad insular) ante el Homo sapiens, consistente en la ausencia de una respuesta defensiva (huida o ataque) ante especies desconocidas. Es la que presentan multitud de especies insulares o la fauna del continente antártico, donde los animales son incapaces de reconocer al hombre como potencial depredador, pero sí tienen una fuerte respuesta defensiva ante sus depredadores naturales.
Referencias
- Surovell, T. A.; Waguespack, N. M. (15 de noviembre de 2008). «How many elephant kills are 14?: Clovis mammoth and mastodon kills in context». Quaternary International 191 (1): 82-97. Bibcode:2008QuInt.191...82S. doi:10.1016/j.quaint.2007.12.001.
- «Human Dispersal and Late Quaternary Megafaunal Extinctions: the Role of the Americas in the Global Puzzle (PDF Download Available)». ResearchGate (en inglés). Consultado el 23 de febrero de 2017.
- Leidy, J. (1858). «Notices of remains of extinct vertebrata, from the Valley of the Niobrara River, collected during the Exploring Expedition of 1857, in Nebraska under the command of Lieut. G.K. Warren, U.S. Top. Eng. By F.V. Hayden, Geologist to the Expedition.». Proceedings of the Academy of Natural Sciences,Philadelphia 10: 20-29.
- Falconer, H.; Murchison, C. (1868). Palaeontological memoirs and notes of H. Falconer, with a biographical sketch of the author 2. London: Robert Hardwicke. pp. 212-239.
- Patterson, D. B.; Mead, A. J.; Bahn, R. A. (2012). «New skeletal remains of Mammuthus columbi from Glynn County, Georgia with notes on their historical and paleoecological significance». Southeastern Naturalist 11 (2): 163-172. doi:10.1656/058.011.0201.
- Osborn, H. F. (1942). Percy, M. R., ed. Proboscidea: A monograph of the discovery, evolution, migration and extinction of the mastodonts and elephants of the world 2. New York: J. Pierpont Morgan Fund. pp. 935-1115.
- Maglio, V. J. (1973). «Origin and evolution of the Elephantidae». Transactions of the American Philosophical Society 63 (3): 1-149. JSTOR 1379357. doi:10.2307/1379357.
- Agenbroad, L. D. (2005). «North American Proboscideans: Mammoths: The state of Knowledge, 2003». Quaternary International. 126-128: 73-25. Bibcode:2005QuInt.126...73A. doi:10.1016/j.quaint.2004.04.016.
- Lister y Bahn, 2007, pp. 12–43.
- Shoshani, J.; Tassy, P. (2005). «Advances in proboscidean taxonomy & classification, anatomy & physiology, and ecology & behavior». Quaternary International. 126–128: 5. Bibcode:2005QuInt.126....5S. doi:10.1016/j.quaint.2004.04.011.
- Lister, A. M.; Sher, A. V.; Van Essen, H.; Wei, G. (2005). «The pattern and process of mammoth evolution in Eurasia». Quaternary International. 126–128: 49. Bibcode:2005QuInt.126...49L. doi:10.1016/j.quaint.2004.04.014.
- Ferretti, M. P. (2003). «Structure and evolution of mammoth molar enamel». Acta Palaeontologica Polonica. 3 48: 383-396.
- Muhs, D. R.; Simmons, K. R.; Groves, L. T.; McGeehin, J. P.; Randall Schumann, R.; Agenbroad, L. D. (2015). «Late Quaternary sea-level history and the antiquity of mammoths (Mammuthus exilis and Mammuthus columbi), Channel Islands National Park, California, USA». Quaternary Research 83 (3): 502-521. Bibcode:2015QuRes..83..502M. doi:10.1016/j.yqres.2015.03.001.
- Agenbroad, L. D. (2012). «Giants and pygmies: Mammoths of Santa Rosa Island, California (USA)». Quaternary International 255: 2. Bibcode:2012QuInt.255....2A. doi:10.1016/j.quaint.2011.03.044.
- Enk, J.; Devault, A.; Debruyne, R.; King, C. E.; Treangen, T.; O'Rourke, D.; Salzberg, S. L.; Fisher, D.; MacPhee, R.; Poinar, H. (2011). «Complete Columbian mammoth mitogenome suggests interbreeding with woolly mammoths». Genome Biology 12 (5): R51. PMC 3219973. PMID 21627792. doi:10.1186/gb-2011-12-5-r51.
- Milius, S. (2011). «Life: DNA: Mammoths may have mixed: Supposedly separate types may really have been one». Science News 180 (12): 13. doi:10.1002/scin.5591801217.
- Lister, A. M.; Sher, A. V. (13 de noviembre de 2015). «Evolution and dispersal of mammoths across the Northern Hemisphere». Science 350 (6262): 805-809. doi:10.1126/science.aac5660.
- Geggel, L. (12 de noviembre de 2015). «North American Mammoths Actually Evolved in Eurasia». LiveScience.com. Purch. Consultado el 15 de noviembre de 2015.
- Lister y Bahn, 2007, pp. 77–95.
- Kurten, B.; Anderson, E. (1980). Pleistocene Mammals of North America. New York: Columbia University Press. pp. 348-354. ISBN 9780231037334.
- Roberto Díaz Sibaja, Eduardo Jiménez Hidalgo, Ma. Luisa García Zepeda (2018). «Una nueva localidad fosilífera en Oaxaca (México) y el registro más austral de Bison latifrons: Implicaciones paleobiogeográficas, paleoecológicas y paleoambientales». Boletín de la Sociedad Geológica Mexicana 70: 201-222.
- McDaniel, G. E.; Jefferson, G. T. (2006). «Dental variation in the molars of Mammuthus columbi var. M. Imperator (Proboscidea, Elephantidae) from a Mathis gravel quarry, southern Texas». Quaternary International. 142-143: 166-177. Bibcode:2006QuInt.142..166M. doi:10.1016/j.quaint.2005.03.014.
- Dirks, W.; Bromage, T. G.; Agenbroad, L. D. (2012). «The duration and rate of molar plate formation in Palaeoloxodon cypriotes and Mammuthus columbi from dental histology». Quaternary International 255: 79-85. Bibcode:2012QuInt.255...79D. doi:10.1016/j.quaint.2011.11.002.
- «Paleoindians, Proboscideans, and Phytotoxins: Exploring the Feasibility of Poison Hunting During the Last Glacial-Interglacial Transition». ResearchGate (en inglés). Consultado el 23 de febrero de 2017.
- «Going Tuskless | African Wildlife Foundation». African Wildlife Foundation. Consultado el 23 de febrero de 2017.
- Lister y Bahn, 2007, pp. 96–111.
- Graham, R. W.; Lundelius, E. L. (2013). «Friesenhahn Cave: Late Pleistocene paleoecology and predator-prey relationships of mammoths with an extinct scimitar cat». Geological Society of America Field Guides 30: 15-31. doi:10.1130/2013.0030(02).
- Haynes, G. (2012). «Elephants (and extinct relatives) as earth-movers and ecosystem engineers». Geomorphology. 157-158: 99. Bibcode:2012Geomo.157...99H. doi:10.1016/j.geomorph.2011.04.045.
- Lister y Bahn, 2007, pp. 45–75.
- Forest, C. E.; Wolfe, J. A.; Molnar, P.; Emanuel, K. A. (1999). «Paleoaltimetry incorporating atmospheric physics and botanical estimates of paleoclimate». Geological Society of America Bulletin 111 (4): 497. doi:10.1130/0016-7606(1999)111<0497:PIAPAB>2.3.CO;2.
- Wiest, L. A.; Esker, D.; Driese, S. G. (2016). «The Waco Mammoth National Monument may represent a diminished watering-hole scenario based on preliminary evidence of post-mortem scavenging». PALAIOS 31 (12): 592-606. doi:10.2110/palo.2016.053.
- Gold, D. A.; Robinson, J.; Farrell, A. B.; Harris, J. M.; Thalmann, O.; Jacobs, D. K. (2014). «Attempted DNA extraction from a Rancho La Brea Columbian mammoth (Mammuthus columbi): Prospects for ancient DNA from asphalt deposits». Ecology and Evolution 4 (4): 329-336. doi:10.1002/ece3.928.
- Pérez-Crespo, V. C. A. N.; Arroyo-Cabrales, J. N.; Benammi, M.; Johnson, E.; Polaco, O. J.; Santos-Moreno, A.; Morales-Puente, P.; Cienfuegos-Alvarado, E. (2012). «Geographic variation of diet and habitat of the Mexican populations of Columbian Mammoth (Mammuthus columbi)». Quaternary International. 276–277: 8-16. Bibcode:2012QuInt.276....8P. doi:10.1016/j.quaint.2012.03.014.
- Gillette, D. D.; Madsen, D. B. (1993). «The Columbian Mammoth, Mammuthus columbi, from the Wasatch Mountains of Central Utah». Journal of Paleontology. 4 67: 669-680. JSTOR 1305939. doi:10.2307/1305939 (inactivo 2015-04-28).
- Davis, O. K.; Agenbroad, L. D.; Martin, P. S.; Mead, J. I. (1984). «The Pleistocene dung blanket of Bechan Cave, Utah». Carnegie Museum of Natural History Special Publication: 267-282.
- Janzen, D. H.; Martin, P. S. (1982). «Neotropical Anachronisms: The Fruits the Gomphotheres Ate». Science 215 (4528): 19. Bibcode:1982Sci...215...19J. doi:10.1126/science.215.4528.19.
- Catling, P. M. (2001). «Extinction and the importance of history and dependence in conservation». Biodiversity 2 (3): 2-14. doi:10.1080/14888386.2001.9712550.
- Lucas, S. G.; Alvarado, G. E.; Vega, E. (1997). «The Pleistocene mammals of Costa Rica». Journal of Vertebrate Paleontology 17 (2): 413-427. doi:10.1080/02724634.1997.10010985.
- Lucas, S. G.; Morgan, G. S.; Estep, J. W.; Mack, G. H.; Hawley, J. W. (1999). «Co-occurrence of the proboscideans Cuvieronius, Stegomastodon, and Mammuthus in the lower Pleistocene of southern New Mexico». Journal of Vertebrate Paleontology 19 (3): 595-597. doi:10.1080/02724634.1999.10011169.
- Lister y Bahn, 2007, pp. 113–139.
- Saunders, J. J.; Daeschler, E. B. (1994). «Descriptive Analyses and Taphonomical Observations of Culturally-Modified Mammoths Excavated at "The Gravel Pit," near Clovis, New Mexico in 1936». Proceedings of the Academy of Natural Sciences of Philadelphia 145: 1-28. JSTOR 4064981. doi:10.2307/4064981 (inactivo 2015-04-28).
- Vasconcelos, Jose. «Mammuthus columbi». Consultado el 24 de agosto de 2018.
- Lister y Bahn, 2007, pp. 141–163.
- Haury, E. W.; Antevs, E.; Lance, J. F. (1953). «Artifacts with Mammoth Remains, Naco, Arizona». American Antiquity 19: 1-24. JSTOR 276409. doi:10.2307/276409.
- Hoppe, K. A. (2004). «Late Pleistocene mammoth herd structure, migration patterns, and Clovis hunting strategies inferred from isotopic analyses of multiple death assemblages». Paleobiology 30: 129-145. doi:10.1666/0094-8373(2004)030<0129:LPMHSM>2.0.CO;2.
- Purdy, B. A.; Jones, K. S.; Mecholsky, J. J.; Bourne, G.; Hulbert, R. C.; MacFadden, B. J.; Church, K. L.; Warren, M. W.; Jorstad, T. F.; Stanford, D. J.; Wachowiak, M. J.; Speakman, R. J. (2011). «Earliest art in the Americas: Incised image of a proboscidean on a mineralized extinct animal bone from Vero Beach, Florida». Journal of Archaeological Science 38 (11): 2908. doi:10.1016/j.jas.2011.05.022.
- Malotki, E.; Wallace, H. D. (2011). «Columbian mammoth petroglyphs from the San Juan River near Bluff, Utah, United States». Rock Art Research: The Journal of the Australian Rock Art Research Association. 2 28: 143-152.
- «Washington State Fossil». State Symbols USA. Consultado el 16 de octubre de 2013.
- «Nebraska State Fossil url=http://www.statesymbolsusa.org/Nebraska/fossil-mammoth.html». State Symbols USA.
- Fiedel, S. (2009). «Sudden Deaths: The Chronology of Terminal Pleistocene Megafaunal Extinction». American Megafaunal Extinctions at the End of the Pleistocene. Vertebrate Paleobiology and Paleoanthropology. pp. 21-37. ISBN 978-1-4020-8792-9. doi:10.1007/978-1-4020-8793-6_2.
- Hills, L. V.; Harington, C. R. (2003). «New radiocarbon dates for Columbian mammoth and Mexican horse from southern Alberta and the Lateglacial regional fauna». Quaternary Science Reviews 22 (14): 1521-1523. Bibcode:2003QSRv...22.1521H. doi:10.1016/S0277-3791(03)00124-0.
- Waters, M. R.; Stafford, T. W. (2007). «Redefining the Age of Clovis: Implications for the Peopling of the Americas». Science 315 (5815): 1122-1126. Bibcode:2007Sci...315.1122W. doi:10.1126/science.1137166.
- Stuart, A. J. (2015). «Late Quaternary megafaunal extinctions on the continents: A short review». Geological Journal 50 (3): 338-363. doi:10.1002/gj.2633.
- «Human arrival scenarios have a strong influence on interpretations of the late Quaternary extinctions (PDF Download Available)». ResearchGate (en inglés). Consultado el 23 de febrero de 2017.
- Martin, P.S. (1967). «Pleistocene overkill». Natural History 76: 32-38.
- Grayson, D. K.; Meltzer, D. J. (2002). «Clovis Hunting and Large Mammal Extinction: A Critical Review of the Evidence». Journal of World Prehistory 16 (4): 313. doi:10.1023/A:1022912030020.
- «Mammoths: Giants of the Ice Age». ResearchGate (en inglés). Consultado el 24 de febrero de 2017.
- «Late Pleistocene Megafaunal Extinctions». ResearchGate (en inglés). Consultado el 23 de febrero de 2017.
Enlaces externos
 Wikimedia Commons alberga una categoría multimedia sobre Mammuthus columbi.
Wikimedia Commons alberga una categoría multimedia sobre Mammuthus columbi.
 Datos: Q1698713
Datos: Q1698713- Multimedia: Mammuthus columbi / Q1698713
 Especies: Mammuthus columbi
Especies: Mammuthus columbi