Mammuthus
Mammuthus es un género extinto de mamíferos proboscídeos de la familia Elephantidae conocidos comúnmente como mamuts. Existieron desde hace aproximadamente 4,8 millones de años hasta hace apenas 3700 años, en las épocas Plioceno (Neógeno tardío), Pleistoceno y Holoceno (Cuaternario).[1][2] Se han descrito numerosas especies, siendo el mamut lanudo la más conocida de todas. Se han hallado fósiles de mamut en Norteamérica, Eurasia y África. No debe confundirse con el género Mammut, que pertenece a una familia distinta (Mammutidae, conocidos comúnmente como mastodontes).
| Mamut | ||
|---|---|---|
| Rango temporal: 4,8 Ma - 0,0037 Ma Plioceno - Holoceno | ||
 Ilustración histórica de un mamut lanudo (1871). | ||
| Estado de conservación | ||
|
Extinto desde ca. 1700 a. C. | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Mammalia | |
| Orden: | Proboscidea | |
| Familia: | Elephantidae | |
| Género: |
Mammuthus Brookes, 1828 | |
| Especies | ||
| Sinonimia | ||
| ||
Nombre
El nombre ruso de los mamuts, del cual deriva su nombre en otras lenguas, es мамонт (mamont), palabra derivada del antiguo ruso маммот (mammont) y este a su vez proviene, probablemente, de la expresión en lengua mansi mang ont, con el significado "cuerno de la tierra". Los mansis se referían a los mamuts como "topos de tierra" o "excavadores", ya que pensaban que eran criaturas vivientes que vivían en túneles subterráneos y que morían si les tocaba la luz del Sol. Según los mansis, esta era la razón por la que nunca se encontraban mamuts vivos.[3]
En español se conocen con el nombre popular de mamuts, que no debe confundirse con el género Mammut, que pertenece a otra familia (Mammutidae), que se incluye entre los Elephantidae conocidos como mastodontes.
Descripción



Los mamuts eran proboscídeos de gran tamaño, de dimensiones iguales y en algunos casos superiores a los elefantes modernos. Las especies más grandes conocidas son M. sungari (mamut del río Songhua), con una altura en la cruz de 5,3 metros y una longitud de 9,1 metros; y el mamut imperial, que tenía una altura mínima de 5 metros en cruz. Entre las especies más pequeñas se encuentran M. exilis (mamut pigmeo), M. lamarmorai (mamut de Cerdeña"), o la raza enana de M. primigenius (mamut lanudo), que tenían una altura en la cruz de 1 a 2 metros. Los mamuts probablemente tenían un peso de entre 6 y 8 toneladas, pero algunos machos excepcionalmente grandes podrían haber sobrepasado las 12 toneladas.
Los mamuts se caracterizaban principalmente por su cabeza abombada, probóscide musculosa y largos colmillos curvados. Las especies norteñas se encontraban recubiertas de pelo para soportar los fríos glaciares de su ecosistema. El colmillo de mamut más grande del que se tiene registro es de un mamut lanudo, cuya longitud alcanzó los 5 metros. Al igual que el elefante asiático, su espalda era arqueada y sus orejas relativamente pequeñas, pero a diferencia de este, su trompa poseía en su extremo dos lóbulos y sus patas delanteras se dotaban de 5 dedos mientras las traseras de 4. En 2005 se descubrió un colmillo de mamut de 3,3 metros de longitud en Illinois[4]
La mayoría de especies de mamut no eran más grandes que el elefante asiático actual. Se han encontrado fósiles de especies de mamuts enanos en la Isla Santa Bárbara (M. exilis) y en Cerdeña (M. lamarmorae). También existía una raza de mamuts lanudos enanos en la Isla de Wrangel, al norte de Siberia, dentro del Círculo Polar Ártico.
Los mamuts que vivían en las regiones más frías poseían unas orejas que sólo medían la quinta parte de la de los elefantes asiáticos. Esta adaptación al frío permitía reducir la pérdida de temperatura gracias a la menor superficie expuesta al ambiente. Los mamuts que vivían en regiones más cálidas debían de tener las orejas más grandes.[5]
Como en los elefantes actuales, la trompa de los mamuts era móvil y prensil, y estaba muy bien adaptada para realizar movimientos precisos como arrancar plantas del suelo y llevarlas a la boca.[3] También las utilizaban para mojarse con agua, para bañarse o cubrirse con barro para protegerse de los mosquitos y otros insectos.
Utilizando su trompa, los mamuts adultos ingerían unos 180 kg de alimentos diarios. Ingerían casi cualquier tipo de vegetal, aunque su alimento preferido era la hierba.[3]
Comían otras partes de las plantas, como las hojas y la corteza de los árboles, en invierno, cuando la nieve cubría la hierba. La dificultad para masticar hierba, (debido a su riqueza en silicio, que desgasta la dentadura) hizo que los mamuts, al igual que los elefantes desarrollasen dientes molares especializados con una superficie de pequeñas crestas de esmalte. Los dientes anteriores de los mamuts eran los premolares, ocho en su infancia y los molares, 24 en total a lo largo de su vida, aunque en edad infantil había 12 al juntarse los molares y premolares. Los molares llegaban a pesar hasta 2 kilos. A medida que los dientes se desgastaban, eran sustituidos por dientes que crecían más atrás. Al llegar a la edad adulta solo quedaban los molares en número de cuatro[6] El examen de sus molares también ha permitido saber que eran animales de crecimiento lento que no llegaban a la madurez sexual hasta aproximadamente los 20 años de edad.[7] Cuando los 6 conjuntos de molares que un mamut tenía a lo largo de su vida se habían desgastado, el animal moría ya que no podía alimentarse. Este proceso solía producirse entre los 60 y 70 años.[8]
Como los elefantes actuales, los mamuts estaban dotados de colmillos formados por 2 dientes incisivos superiores. Los colmillos de los mamuts eran más grandes que los de sus parientes actuales y su forma también era diferente. Estos colmillos solían medir unos 2 metros y medio, y pesar cerca de 50 kilos. Cuando nacían, las crías de mamut no tenían colmillos. A los 6 meses de edad les crecían unos dientes de leche de sólo 5 centímetros de largo que era sustituidos por colmillos permanentes un año más tarde. Estos colmillos permanentes no dejaban de crecer hasta la muerte del animal.[9]
Las pinturas rupestres permiten saber algunas cosas sobre los mamuts que nunca se habrían podido descubrir a partir de sus esqueletos: las pinturas prehistóricas presentan los mamuts con la espalda curvada, mientras que los fósiles parecen indicar que la espalda de estos proboscídeos era plana. Esta curvatura de la espalda se debía a una joroba de grasa, músculo o pelo que los mamuts tenían en la espalda. La cola de los mamuts era más corta que la de los elefantes.
Basándose en estudios realizados en sus parientes próximos, los elefantes actuales, los mamuts probablemente tenían un periodo de gestación de 22 meses y daban a luz una sola cría en cada embarazo.
Comportamiento
La estructura social de los mamuts era probablemente la misma que la de los elefantes asiáticos, con manadas compuestas de hembras encabezadas por una matriarca, y los machos que vivían solos o en grupos pequeños después de llegar a la madurez sexual. Una vez que abandonaban la manada donde se habían criado, los machos se dedicaban a buscar hembras de otros grupos para aparearse.[10] Tanto el apareamiento como los nacimientos tenían lugar durante una época corta del año debido a la corta estación estival de la estepa del mamut, permitiendo que las crías naciesen en la época del año que había más abundancia de alimentos.
Las trompas del mamut, un ejemplo de dimorfismo sexual, ya que eran más grandes en los machos que en las hembras y estaba adaptadas para las luchas entre machos .Estos libraban unos combates por las hembras en los que intentaban herir al oponente en las partes vulnerables de la espalda y el tórax con sus colmillos. Es probable que la posibilidad de quedar gravemente herido hiciese que no hubiese combates desiguales. Se conoce un caso en que dos mamuts estaban tan igualados que sus colmillos quedaron encastrados unos con otros e, incapaces de separarse, los dos mamuts murieron de hambre.[6]
Sin embargo, al contrario de lo que se creía hasta ahora, no emigraban al sur en los periodos más duros del año. Ninguno de los animales analizados recientemente en Siberia salió de una zona de entre 700 y 800 km de diámetro.[11]
Hábitat
En la época de transición entre el Plioceno y el Pleistoceno, los continentes se encontraban prácticamente en la misma posición que en la actualidad. Las placas tectónicas sobre las que reposan probablemente no se han movido más de 100 km las unas en relación con las otras desde principios del Pleistoceno.
El clima del Pleistoceno se caracterizaba por ciclos glaciales repetidos en los que los glaciares continentales llegaban hasta el paralelo 40 en algunos puntos. Se calcula que durante el máximo glacial, el 30% de la superficie de la Tierra estaba cubierta de hielo. Además, una zona de permafrost se extendía hacia el sur desde el límite de la capa de hielo, unos pocos centenares de km en Norteamérica y más lejos en Eurasia. La temperatura media anual en los límites del hielo era de -6 °C; en el límite del permafrost, de 0 °C.
Al sur de las capas glaciales se acumulaban grandes lagos, porque las vías de drenaje estaban bloqueadas y el aire más frío reducía la evaporación. El centro-norte de Norteamérica estaba completamente cubierto por el lago Agassiz. Más de cien cuencas, ahora secas o prácticamente secas estaban llenas en el oeste americano. El lago Bonneville, por ejemplo, estaba donde se encuentra actualmente el Gran Lago Salado. En Eurasia se desarrollaron grandes lagos como resultado del drenaje de los glaciares. Los ríos tenían un caudal mayor y se entrelazaban. Los lagos africanos estaban más llenos, supuestamente debido a una menor evaporación.
Debido al clima, más frío y seco que en la actualidad, los desiertos eran más secos y extensos. Las precipitaciones eran menores debido a la reducción de la evaporación oceánica y continental, por lo que los bosques eran raros. El ecosistema terrestre predominante era las llanuras de hierba, prados y sabanas de sauces.[12] Las estepas cubrían gran parte de la zona subglacial en Eurasia.
Las condiciones climáticas permitían una diversidad vegetal que soportaba muchas especies diferentes de animales que normalmente habitarían en zonas distintas. Especies que actualmente viven en el norte, como los lemmini convivían con especies que actualmente viven en el sur, como las mofetas. R. Dale Guthrie argumenta que la larga temporada de crecimiento de las plantas del Pleistoceno era más favorable a la diversidad que a la homogeneidad. Esto se debe porque estos ecosistemas eran mejores para los animales no especializados, que necesitaban comer diferentes tipos de animales para alimentarse correctamente.[13]
La vasta región que se extendía desde el este de Europa, a través de Siberia hasta Alaska y el Yukón recibía el nombre de la estepa del mamut. La abundancia de hierba, forbias y lamiáceas creó un ecosistema de estepa especialmente favorable para los grandes mamíferos como el mamut. La flora del Pleistoceno era especialmente nutritiva debido a que el duro clima obligaba a las plantas a acumular fibras y carbohidratos para poder sobrevivir.[14]
Extinción
La mayoría de los mamuts se extinguieron a finales de la última glaciación. Aún no se ha encontrado una explicación definitiva de su extinción. Una pequeña población sobrevivió en la isla de San Pablo (Alaska) hasta el año 6000 a. C.[2] aproximadamente y los mamuts enanos de la isla de Wrangel (Siberia) no se extinguieron hasta el año 2000 a. C. aproximadamente.[15] Hay diversas teorías para explicar la extinción de los mamuts en concreto y la extinción de la Megafauna del Pleistoceno en general, pero lo más probable es que la extinción no se deba a una sola causa, sino a una combinación de varios factores.
En Norteamérica la extinción de los mamuts, según los últimos descubrimientos científicos, está datada de hace entre 10.500 y 7.500 años. Para llegar a esta conclusión, los científicos se han basado en hallazgos de ADN encontrados en muestras de tierra del permafrost, en la tundra que está en Alaska junto al río Yukón. Las muestras analizadas de tierra contenían restos de orina y heces de mamut.[16]
En cambio en Inglaterra la extinción del mamut se remonta a hace 14 000 años, según el descubrimiento de huesos en Shropshire, siendo en Europa occidental los restos más recientes de mamut lanudo.[17]
Cambio climático

Hace unos trece mil años, la temperatura y la humedad global comenzaron a aumentar, permitiendo la migración de vegetales comestibles hacia el norte. Durante un tiempo, los grandes mamíferos del norte pudieron aprovechar este incremento del alimento disponible, pero finalmente el cambio climático se volvió perjudicial para ellos. Las nuevas condiciones climáticas favorecían a los árboles, que prosperaron a costa de los pequeños arbustos y matorrales de las que se alimentaban los mamuts y los otros grandes mamíferos. Algunos de los animales, como los bisontes y los alces, se adaptaron a esta nueva situación, pero otros, como los mamuts, quedaron diezmados y acabaron extinguiéndose.[18]
Además del cambio en la vegetación y en los ecosistemas, el incremento de la temperatura (de 6 °C entre el 13 000 a. C. y el 8000 a. C.) habría añadido mucha presión a los mamíferos adaptados al frío, causando la extinción. En el caso de los animales como el mamut lanudo, su espeso pelaje, que contribuye a conservar el calor corporal en climas glaciales, podría haber evitado la expulsión del exceso de calor causando la muerte del animal por hipertermia. Los mamíferos grandes, que tienen una menor relación área de superficie-volumen, habrían sufrido más que los mamíferos pequeños.
Además, investigaciones recientes han demostrado que la temperatura media anual del periodo interglacial actual que se ha experimentado en los últimos 10 000 años no es más alta que la de otros periodos interglaciales anteriores, de manera que los mismos mamíferos grandes habían sobrevivido a aumentos de temperatura similares. Por lo tanto, el incremento de la temperatura por sí solo no es una explicación suficiente.[19][20]
Caza por parte del ser humano

En 2015, un estudiante de doctorado de la Universidad de Míchigan y su director de tesis (Michael Cherney y Daniel Fisher) propusieron la hipótesis de que la caza por parte del ser humano podría ser la principal causa de la extinción del mamut lanudo, en lugar del cambio climático.[21] En esta investigación se estudiaron los colmillos de mamuts siberianos juveniles para estimar la edad del destete (momento en el que un animal mamífero deja de mamar), y se observó que esta edad había disminuido de tres años durante los 30 000 años previos a la extinción de la especie. La edad del destete tiende a aumentar debido el cambio climático, según se ha observado en las poblaciones modernas de elefantes, y a disminuir debido a la presión de los depredadores, produciendo que los individuos de la especie maduren más rápido.[21] Se sabe que Homo erectus ya consumía carne de mamut hace 1,8 millones de años.[22]
Los impulsores de esta hipótesis, postulada por primera vez por Paul S. Martin de la Universidad de Arizona en los años sesenta, señalan la aparente coincidencia de la expansión de los humanos por todo el mundo con la extinción de numerosas especies de animales. Un argumento que refuerza esta hipótesis es que se calcula que el 80% de las especies de grandes mamíferos de Norteamérica se extinguieron en los mil años siguientes a la llegada del ser humano al continente.[23] Otro ejemplo es Madagascar, colonizado desde hace unos 1500 años, y donde se extinguieron durante los siglos siguientes a la llegada del ser humano los hipopótamos que habitaban la isla, además de grandes primates como el lémur gigante Megaladapis edwardsi. En cuanto a los mamuts, esta hipótesis podría verse reforzada por los descubrimientos de mamuts con puntas de proyectiles clavadas en los esqueletos,[24] aunque en realidad esto solo probaría que los hombres cazaban mamuts.
Sin embargo, también existen argumentos en contra de esta hipótesis. Los métodos de caza primitivos de los humanos prehistóricos no parecen haber tenido un impacto tan profundo sobre las poblaciones de los mamíferos en otros lugares como África, donde el ser humano llegó mucho antes y no hubo ninguna extinción significativa. Otro de los argumentos en contra de esta hipótesis es que, en la naturaleza, los depredadores no tienden a hacer una caza excesiva de sus presas, ya que las necesitan para alimentarse y reproducirse.[25] Aun así, los humanos podrían ser una excepción gracias a su capacidad de cambiar a otro tipo de presa o incluso a una alimentación a base de vegetales si una determinada especie se extingue.
Gran enfermedad
En 1997, MacFee y Marx propusieron la hipótesis de que la extinción de los grandes mamíferos del Pleistoceno (entre ellos, los mamuts) se debieron en buena medida a una epidemia surgida como efecto indirecto de la llegada del ser humano.[26][27] Los seres humanos, o los animales que viajaban con ellos, introdujeron enfermedades virulentas en poblaciones de mamíferos nativos. Se parte del supuesto de que las grandes especies eran más vulnerables, ya que las especies pequeñas tienen una resistencia mayor gracias a factores como una gestación más corta, poblaciones más numerosas, etc. Estos autores atribuyen al hombre la causa porque otras migraciones anteriores de mamíferos a Norteamérica desde Eurasia no habían causado extinciones.[26]
No obstante, resulta muy difícil sostener que una enfermedad pueda a la vez tener un espectro tan amplio como para acabar con especies de distintas clases (aves, mamíferos, reptiles), y sin embargo, ser tan selectiva como para no afectar a especies emparentadas genéticamente, incidiendo solo en función del tamaño. A esto se añade que una enfermedad debe ser extremadamente virulenta para exterminar a todos los miembros de una misma especie. Así, incluso una enfermedad tan virulenta como el Virus del Nilo Occidental difícilmente podría causar por sí sola la extinción completa de una especie, salvo que existan otros factores que estén afectando a esa especie.[28]
Descubrimiento
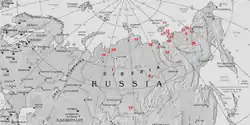
La primera expedición occidental que fue a Siberia para estudiar los enormes huesos que se encontraban allí fue la del anatomista británico Hans Sloane, que en 1728 examinó restos de fósiles de dientes y colmillos y determinó que pertenecían a una especie de elefante. Para explicar la presencia de animales tropicales en latitudes tan altas, sugirió que su extinción se debió a un enorme cambio climático y al Diluvio Universal. Diez años más tarde, Philosophical Magazine publicó la obra del alemán Johann Breyne, incluyendo sus análisis de fósiles e ilustraciones que le había enseñado Daniel Messerschmidt. Breyne corroboró la teoría de Sloane de que los huesos pertenecían a un tipo de proboscídeo.[6]
Georges Cuvier revisó la información disponible sobre el mamut en 1796 y, con la ayuda de las ilustraciones de Messerschmidt, llegó a la conclusión de que el mamut era diferente de los elefantes actuales. En 1799, Johann Friedrich Blumenbach definió el mamut lanudo como Elephas primigenius. Algunos años más tarde, en 1806, el escocés Michael Adams encontró un cadáver de mamut que todavía conservaba parte del pelo y la piel, y con el esqueleto prácticamente intacto. Este mamut, que recibió el nombre de mamut de Adams fue montado en el Museo Zoológico de San Petersburgo.[6]
También se hicieron descubrimientos de mamuts en Norteamérica. En 1743, esclavos negros identificaron unos molares en el este de los Estados Unidos como propias de un elefante. Otro diente molar enorme fue el tema de un informe enviado por David Rittenhouse a la American Philosophical Society en 1786. La expedición de Jefferson y Clark de 1807 recuperó diversos molares que tanto Clark como Caspar Wistar identificaron como pertenecientes a un mamut.[6]
Clonación

Las temperaturas glaciales de Siberia han permitido que algunos restos de mamut hayan quedado suficientemente bien conservados como para extraer su ADN en buen estado. El descubrimiento de una cría de mamut al noroeste de Siberia a mediados del 2007 da esperanzas de poder clonar los mamuts. La cría, muerta hace más de 9000 años, aún tenía intactos los ojos, la trompa y una parte del pelaje. Las únicas partes que le faltaban eran la cola y una oreja.[29]
El proceso de clonación completo propuesto requeriría resto de tejido blando de mamut del cual se pueda extraer una cadena de ADN. Este ADN sería posteriormente fusionado con un óvulo de elefante asiático, el pariente vivo más próximo de los mamuts. Dicho óvulo sería insertado en el útero de una hembra de elefante asiático y, si el proceso tuviese éxito, 22 meses más tarde nacería una cría de mamut.[30] En el 2003 se anunció los planes más ambiciosos hasta ahora, encabezados por científicos japoneses de la Universidad de Kinki y rusos del Centro de Investigación Vektor de Virología y Biotecnología.[31]
Sin embargo, este proceso de clonación aparentemente sencillo, presenta dificultades en cada uno de los diversos pasos. Para comenzar, la cantidad de ácido desoxirribonucleico que se ha obtenido hasta ahora no es suficiente ni en cantidad ni en calidad como para clonar un mamut.[32] No es suficiente con encontrar una cierta cantidad de ADN, hay que encontrar una cadena entera intacta de ADN. Eso es poco probable y algunos genetistas opinan que de hecho es imposible. Además, hay que tener en cuenta que hasta ahora no se ha conseguido nunca clonar un mamífero únicamente por medio del ADN. (La Oveja Dolly, por ejemplo, fue clonada a partir de una célula somática adulta).[33] Tampoco se ha conseguido nunca clonar un elefante, de manera que todavía parece improbable que se pueda clonar un mamut.
La posibilidad de clonar un mamut cambió radicalmente en noviembre de 2008, cuando se publicó un estudio realizado por un equipo de investigación japonés, liderado por Sayaka Wakayama, en el Centro de Biología del Desarrollo del Instituto RIKEN en Kobe, ha clonado un ratón que ha permanecido 16 años congelado a -20 °C. Si bien el principal impedimento para llevar a la práctica la clonación era utilizar ADN que estuviese dañado por los cristales de agua congelada, se descubrió que las células nerviosas del cerebro poseían glúcidos que permitían mantener los núcleos de igual forma que la criopreservación, sin daño de ADN.[34] Ahora, el principal inconveniente está representado por la posibilidad de que un núcleo de mamut sea viable en una célula de elefante.
La gestación también presentaría problemas adicionales. No sólo el tiempo de gestación podría tener una duración diferente a la de los elefantes, cosas que impondría una presión añadida al feto del mamut como a la madre elefante, además es probable que el cuerpo de la madre no pudiese cubrir las necesidades de un animal más grande y de metabolismo posiblemente diferente como son los mamuts.[30] También es posible que, una vez nacido, la cría de mamut fuese rechazada por su madre o no tolerase bien la leche de elefante.
Dentro del ámbito científico, hay opiniones divididas entre los que creen que es posible clonar un mamut y los que piensan que es imposible. Dick Mol es uno de los que duda de que sea posible, mientras que uno de sus colegas en la expedición que descubrió el Mamut de Jarkov (Siberia), Larry Agenbroad, apuntaba que hace unos años había científicos que decían que nunca se podría clonar una oveja.[30] Por otro parte, Adrian Lister, uno de los paleontólogos más reputados en el estudio de los mamuts también ha cuestionado la ética de clonar estos animales: «El hábitat natural del mamut ya no existe. Estaríamos creando un animal como atracción en un parque temático. ¿Es ético?»[32]
Esta pregunta también se la plantea en la universidad de McMaster, Mamdouh Shoukri, vicepresidente para las investigaciones y los asuntos internacionales, sobre la base de las investigaciones de Hendrik Poinar, experto en genética molecular evolutiva, del departamento de antropología y patología de esa universidad que lleva desarrollando un proyecto para descifrar el genoma del mamut. Para ello, utilizando una máquina de última tecnología, trabajan en el ADN nuclear vital del mamut, en lugar del mitocondrial, y en las primeras horas de estudio consiguieron secuenciar aproximadamente el uno por ciento del genoma completo del mamut, es decir, los primeros 30 millones de pares de bases. Esto según dicen, hace teóricamente posible recrear un híbrido del extinto animal.[35]
Especies
Mammuthus africanavus
El mamut africano es una de las especies africanas de mamut. Apareció por primera vez hace aproximadamente 4,8 millones de años durante el Plioceno, y se han encontrado fósiles en Chad, Libia, Marruecos y Túnez. Aunque se extinguieron (relativamente) poco después, hace 4 millones de años, es la especie de la cual descendieron el mamut meridional y las especies posteriores de mamut. De hecho, su nombre científico, Mammuthus africanavus, significa mamut antepasado africano. Aunque su origen es africano, ni el 'Mammuthus africanavus ni sus descendientes estaban estrechamente relacionados con el elefante africano. Su linaje se separó de los elefante asiáticos hace aproximadamente 5 millones de años, a principios del Zancliense.
Mammuthus columbi

El mamut colombino es una de las mayores especies de mamut. Los ejemplares de mayor tamaño alcanzaban los 4 metros de altura y pesaban hasta 10 toneladas, pero lo más normal era que midiesen un poco más de 3 metros de altura y que pesaran unas 6 toneladas. La cabeza pesaba entre un 12% y un 25% del peso total del cuerpo.[36] Vivió a finales del pleistoceno y fue una de las últimas especies de la megafauna del Pleistoceno en extinguirse. Se han encontrado fósiles en Rancho La Brea (California), en Texas[37] y en México, donde sus restos son bastante comunes, y se sabe que llegaron hasta Nicaragua.
Como las condiciones climáticas que reinaban en Norteamérica eran menos severas que las de Siberia, el mamut colombino estaba menos adaptado al frío que el mamut lanudo. Tenía una capa de pelo mucho menos espesa, pero su característica principal eran sus colmillos, que eran los más grandes de todas las especies de mamut y llegaban a medir hasta 5 metros de largo.[38]
Mammuthus exilis
El mamut pigmeo es una especie enana de mamut que vivió en la isla de Santa Rosa, en el Archipiélago del Norte de California. Se cree que una población de mamuts colombinos atravesaron el Canal de Santa Bárbara para emigrar del sur de California a Santa Rosa hace decenas de miles de años.[39] En Santa Rosa comenzó un proceso de enanismo que redujo gradualmente la medida de estos proboscídeos.
Los mamuts pigmeos se extinguieron hace más de 12 000 años a causa de una combinación de caza por parte de los pobladores precolombinos, cambios climáticos y una pérdida del hábitat, a medida que el nivel del mar hizo que Santa Rosa se dividiese en varias islas.
Mammuthus lamarmorai
El mamut de Cerdeña es la única especie de mamut endémica de las islas del Mediterráneo. Como la otra especie insular, el mamut pigmeo, el mamut de Cerdeña padeció un proceso de enanismo que le dio un tamaño muy inferior al de otros mamuts, aproximadamente un metro y medio de altura.
Mammuthus meridionalis

El mamut meridional fue la primera especie en abandonar África para llegar a Eurasia y mucho más tarde a Norteamérica a través del estrecho de Bering, aprovechando que el nivel del mar era mucho más bajo que en la actualidad. Esta especie de mamut vivía en las regiones de bosque abierto y de clima más benigno y se alimentaba de hierba y hojas.[40] Es el antepasado directo del mamut colombino y el mamut de las estepas.
El mamut meridional era más grande que los elefantes actuales, con una altura de 4 metros y un peso de 10 toneladas. También era diferente de los mamuts posteriores, ya que vivía en una zona templada donde no necesitaba la espesa capa de pelo típica del mamut lanudo o del mamut de la estepa.[41] Cuando el nivel de aumentó lo suficiente, las poblaciones de M. meridionalis de Norteamérica y Europa evolucionaron en M.imperator y M. trogongtherii, respectivamente.
Mammuthus primigenius
El mamut lanudo se conoce a partir de los huesos y cadáveres congelados encontrados al norte de Eurasia y Norteamérica, con ejemplares especialmente bien conservados en Siberia (Rusia). Los ejemplares más antiguos conocidos datan de sedimentos de aproximadamente 250 000 años de antigüedad, de la Glaciación de Riss; se trata de formas de transición, y las formas más avanzadas aparecieron posteriormente.[6] Se desarrollaron a partir del mamut de las estepas[42]
Los mamuts lanudos se caracterizaban por su pelo, especialmente espeso, con pelos que medían hasta 90 centímetros de longitud. Otra adaptación al clima glacial de su hábitat era una capa de grasa que en ocasiones medía hasta 9 centímetros de grosor.[42] Desaparecieron de la mayoría de su hábitat a finales del Pleistoceno, pero una subespecie enana sobrevivió en la isla de Wrangel hasta aproximadamente el año 1700 a. C.[43]
Mammuthus subplanifrons
El mamut sudafricano apareció hace unos 5 millones de años y es la especie más antigua de mamut. Su hábitat se extendía desde Sudáfrica hasta Kenia. Los primeros restos se descubrieron en 1928 en Sudáfrica y consistieron en unos cuantos colmillos y molares y fragmentos de maxilares. El mamut sudafricano ya presentaba una ligera curvatura en los colmillos, típica de los mamuts.[44]
Mammuthus trogontherii
El mamut de las estepas se extendió en gran parte del norte de Eurasia y Norteamérica a finales del pleistoceno (entre 600 000 y 250 000 años atrás). Probablemente evolucionó a partir de poblaciones siberianas del mamut meridional. Por lo tanto, representa un primer estado en la evolución de los mamuts de la estepa y de la tundra y es un antepasado del mamut lanudo.
Medía hasta 4,7 metros de altura, lo que le convertía en uno de los proboscídeos más grandes que hayan existido nunca, junto con Deinotherium. Tenía unos molares con más crestas de esmalte que las de M. meridionalis; eso se debía a su dieta basada en las gramíneas.[45] Sus colmillos en forma de espiral podían medir hasta 5,2 metros en los machos viejos. Los mamuts de las estepas fueron sustituidos por sus descendientes, los mamuts lanudos, hace unos 250 000 años. Los colmillos de las hembras, en cambio, eran más delgados y poco curvados. El ejemplar más completo conocido es de una hembra descubierta en 1996 en Kikinda, Serbia.
Los mamuts y la criptozoología
A lo largo de los años ha habido numerosos rumores de que el mamut realmente no está extinto y pequeñas manadas aisladas sobrevivían en la tundra del hemisferio norte, vasto y poco poblado. A finales del siglo XIX, según Bengt Sjögren, los rumores persistían sobre la supervivencia de mamuts en los lugares más remotos de Alaska.[46] En octubre de 1899, un tal Henry Tukeman contó detalladamente cómo había matado un mamut en Alaska y luego había dado el ejemplar al "Smithsonian Institution" de Washington D. C. Pero el museo negó el asunto, que resultó ser una novatada. Sjögren cree que el mito comenzó cuando el biólogo americano C.H. Townsend, durante un viaje en Alaska, vio a esquimales cambiar unas defensas gigantescas, él les preguntó si todavía vivían mamuts en Alaska y les mostró un dibujo del animal.
En el siglo XIX, se transmitieron varios informes sobre "grandes bestias peludas" a las autoridades rusas por un miembro de una tribu siberiana, pero nunca se encontraron pruebas científicas sobre eso. En 1946, un hombre de negocios francés que trabajaba en Vladivostok, Sr. Gallon, aseguró que en 1920 había encontrado a un trampero ruso que decía haber visto a "elefantes" gigantes y peludos, viviendo en el corazón de la taiga. Gallon añadía que este trampero no había oído hablar antes de mamuts y que hablaba de mamuts como de animales que vivían en el bosque, en una época en la que se los imaginaba viviendo en la tundra y en la nieve.[46]
Referencias
- «Woolly Mammoth (Mammuthus primigenius)». The Academy of Natural Sciences of Drexel University. Archivado desde el original el 4 de marzo de 2012. Consultado el 7 de marzo de 2012.
- Guthrie RD (junio de 2004). «Radiocarbon evidence of mid-Holocene mammoths stranded on an Alaskan Bering Sea island». Nature 429 (6993): 746-9. PMID 15201907. doi:10.1038/nature02612.
- Haines, Tim (2001). «Mammoth Journey». Walking with Beasts. Londres: BBC Books. Data de acceso: 27 de marzo 2008.
- Recently discovered 11-foot long Woolly Mammoth tusk on display at the Illinois State Museum Comunicado de prensa del Illinois Department of Natural Resources, 14 de agosto del 2006
- Les oreilles; L'encyclopédie du mammouth
- «Woolly Mammoth (Mammuthus primigenius)». Archivado desde el original el 22 de septiembre de 2008. Consultado el 4 de septiembre de 2008.
- Keddie, Grant. The Mammoth Story. Archivado desde el original el 27 de septiembre de 2007. Consultado el 5 de septiembre de 2008.
- Molaires; L'encyclopédie du mammouth
- Défenses; L'encyclopédie du mammouth
- Haines, Tim (2001). «Mammoth Journey». Walking with Beasts. Londres: BBC Books. Data d'accés: 27-3-2008.
- La ciencia descifra la vida diaria de los mamuts
- Pleistocene Park Underway: Home for Reborn Mammoths?
- R. Dale Guthrie, 1984
- R. Dale Guthrie, 1995
- Vartanyan, S.L.; Kh. A. Arslanov; T. V. Tertychnaya; S. B. Chernov (1995). «Radiocarbon Dating Evidence for Mammoths on Wrangel Island, Arctic Ocean, until 2000 BC». Radiocarbon (Department of Geosciences, The University of Arizona) 37 (1): pp 1-6. Archivado desde el original el 12 de diciembre de 2007. Consultado el 10 de enero de 2008.
- «Extinción de los mamuts». www.solociencia.com. Consultado el 16 de febrero de 2017.
- «Arqueología y paleontología». www.solociencia.com. Consultado el 16 de febrero de 2017.
- Mammoth Extinction Caused by Trees, Study Suggests, a National Geographic News.
- Andersen, S. T (1973). The differential pollen productivity of trees and its significance for the interpretation of a pollen diagram from a forested region. A Quaternary plant ecology: the 14thsymposium of the British Ecological society, University of Cambridge, 28–30 March 1972 (ed. Birks, H. J. B. and West, R. G). Oxford: Blackwell Scientific Pubs. ISBN 0-632-09120-7.
- Ashworth, C.A. (1980). «Environmental implications of a beetle assemblage from the Gervais formation (Early Wisconsinian?), Minnesota». Quat. Res. (en inglés) 13: 200-12.
- «Shift in weaning age supports hunting-induced extinction of Siberian woolly mammoths». Science Daily (en inglés). 15 de octubre de 2015. Consultado el 12 de junio de 2016.
- Levy, Sharon (2006). «Clashing with Titans». BioScience 56 (4): pp 292-298. Consultado el 28 de març del 2008.
- Anthony D. Barnosky, Paul L. Koch, Robert S. Feranec, Scott L. Wing, Alan B. Shabel. «Assessing the Causes of Late Pleistocene Extinctions on the Continents». Science 306 (5693): 70-75.
- Diamond, J. (1984). Historic extinctions: a Rosetta stone for understanding prehistoric extinctions. A Quaternary extinctions: A prehistoric revolution (ed. P.S. Martin and R.G. Klein). Tucson, AZ: Univ. Arizona Press. pp. 824-62. ISBN 0-8165-1100-4.
- May, R. M. (2001). Stability and complexity in model ecosystems. Princeton: Princeton Univ. Press. ISBN 0-691-08861-6.
- MacFee, R.D.E. & Marx, P.A. (1997). Humans, hyperdisease and first-contact extinctions. A Natural Change and Human Impact in Madagascar (eds S. Goodman & B.D. Patterson). Washington D.C.: Smithsonian Press. pp. 169-217. ISBN 1-56098-683-2.
- MacFee, R.D.E. & Marx, P.A. (1998). «Lightning Strikes Twice: Blitzkrieg, Hyperdisease, and Global Explanations of the Late Quaternary Catastrophic Extinctions». American Museum of Natural History. Archivado desde el original el 25 de febrero de 2021. Consultado el 16 de diciembre de 2008.
- Lyons, K, Smith, F.A., Wagner, P.J., White, E.P., and Brown, J.,H. (2004). «Was a 'hyperdisease' responsible for the late Pleistocene megafaunal extinction?». Ecology 7: 859-68. Archivado desde el original el 4 de septiembre de 2006.
- Roger Highfield (11 de julio de 2007). «Found, a perfect baby mammoth». The Daily Telegraph (en inglés). Archivado desde el original el 13 de julio de 2007.
- «Will Mammoths Walk Again?». Discovery Channel (en inglés). 9 de marzo de 2001. Archivado desde el original el 14 de febrero de 2007.
- «Scientists 'to clone mammoth'». BBC News (en inglés). 18 de julio de 2003.
- Woolly Mammoth Resurrection, "Jurassic Park" Planned, National Geographic News]
- McLaren A (2000). «Cloning: pathways to a pluripotent future». Science 288 (5472): 1775-80. PMID 10877698.
- Sayaka Wakayama; Hiroshi Ohta; Takafusa Hikichi; Eiji Mizutani; Takamasa Iwaki; Osami Kanagawa; Teruhiko Wakayama (11 de noviembre de 2008). «Production of healthy cloned mice from bodies frozen at −20°C for 16 years». PNAS - Proceedings of the National Academy of Sciences (en inglés) 105 (45). doi:10.1073/pnas.0806166105.
- «Descifrado el genoma del mamut lanudo». Solo Ciencia.
- «Mammoth Facts» (en inglés). Archivado desde el original el 29 de junio de 2012. Consultado el 5 de enero de 2022.
- «Site Background and History». Official Waco Mammoth Site - Waco, Texas (en inglés). Archivado desde el original el 17 de septiembre de 2008. Consultado el 9 de noviembre de 2017.
- Mammuthus Columbi; L'encyclopédie du mammouth
- Channel Islands (USA) pygmy mammoths (Mammuthus exilis) compared and contrasted with M. columbi, their continental ancestral stock, L. D. Agenbroad
- Palmer, Douglas (1999). The Marshall Illustrated Encyclopedia of Dinosaurs & Prehistoric animals. London: Marshall Editions Developments Ltd. 3-8290-6747-X.
- Fossil Info
- «Yukon Beringia Interpretive Centre - Woolly Mammoth». www.beringia.com. Archivado desde el original el 13 de marzo de 2008. Consultado el 12 de marzo de 2008.
- Ronald M. Nowak: Walker's Mammals of the World. Johns Hopkins University Press, 1999 ISBN 0-8018-5789-9
- «ElefantAsia». Archivado desde el original el 29 de octubre de 2007. Consultado el 5 de septiembre de 2008.
- Mammuthus trogongtherii; L'encyclopédie du mammouth
- Sjögren, Bengt. Farliga djur och djur som inte finns, Prisma, 1962.
Bibliografía
- Lister, A.; Bahn, P. (1994). Mammoths. Nova York: Macmillan.
- Versini, A. (2004). Au temps des mammouths. París: Muséum national d'histoire naturelle. ISBN 2-85653-567-4.
- Joger, U.; Kamcke, C. (2005). Mammut: Elefanten der Eiszeit. ISBN 3-938693-04-5.
- Martin, P. (2005). Twilight of the mammoths: Ice Age extinctions and the rewilding of America. Berkeley: University of California Press.
- Capelli, C. et al. (2006). A nuclear DNA phylogeny of the woolly mammoth (Mammuthus primigenius). Molecular Phylogenetics and Evolution 40 (2) 620–627.
- Levy, S. (2006). Clashing with Titans. BioScience 56 (4) 292-298.
- Lister, A.; Bahn, P. (2007). Mammoths: Giants of the Ice Age. University of California Press. ISBN 0-520-25319-1.
Véase también
Enlaces externos
- "Mammoth find. New evidence links Siberian, North American mammals", Pequeño mamut originario de Norteamérica hallado en Siberia, artículo del Canada National Post, 5 de septiembre de 2008.
- The Mammoth Story de Grant Keddie. Artículo publicado en el sitio web del Royal British Columbia Museum
- 10.000 años muy bien llevados. Hallado en Siberia un bebé de mamut que se ha conservado casi íntegro gracias a los hielos nota de ElPais.com.
- "Sequencing the nuclear genome of the extinct woolly mammoth", Secuencian el 80% del genoma del Mamut Lanudo, Nota de la Revista Nature, 20 de noviembre de 2008.
- "Mammoth Genome Project" Archivado el 7 de mayo de 2010 en Wayback Machine., Pennsylvania State University.
- Imágenes artísticas de varias especies de mamut.
- ¿Es de verdad posible clonar un mamut? (Vídeo explicativo del posible proceso tras el hallazgo de sangre líquida en un espécimen encontrado en Siberia) ABC.es, 6 de junio de 2013.
| Control de autoridades |
|
|---|