Myriapoda
Los miriápodos o miriópodos[1] (Myriapoda, del griego μυρια myria, "innumerable" y ποδος podos, "pie") son un subfilo de artrópodos mandibulados, similares a los insectos en algunos aspectos, pero con muchos caracteres que los diferencian de estos. Comprenden cuatro grupos bien definidos: los ciempiés, milpiés, paurópodos, y sínfilos, todos terrestres; se han descrito más de 16 550 especies.[2] Todos tienen en común un cuerpo compuesto por dos regiones: cabeza y tronco, este último es largo y está formado por muchos segmentos y multitud de pares de patas (Eumillipes persephone, un milpies de Australia ostenta el récord, con 653 pares de patas[3]).
| Miriápodos | ||
|---|---|---|
| Rango temporal: Silúrico - Holoceno | ||
 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Arthropoda | |
| Subfilo: | Myriapoda | |
| Clases | ||
En la clase Chilopoda, el par de patas más cercano a la cabeza está modificado y hace las funciones de aguijones inoculadores de veneno. Los cuatro grupos también exhiben diferencias marcadas en aspectos tales como la alimentación, por ejemplo, dándose el caso de especies predadoras (clase Chilopoda) y otras de hábitos detritívoros (clase Diplopoda).
Durante el Carbonífero Superior y el Pérmico existieron miriápodos de más de 1 m de longitud.[4]
Características

La cutícula presenta diferentes grados de esclerotización y calcificación, y es más o menos impermeable, pero carece de la capa cérea típica de los insectos, por lo que quedan más o menos expuestos a la desecación, cosa que evitan refugiándose en lugares húmedos y frescos.
La cabeza lleva las antenas, en la base de las cuales se abren los poros de los órganos de Tömösvary, órganos sensoriales de función desconocida. Los ojos son simples, sin verdaderos omatidios, aunque en algunos grupos (como los escutigeromorfos) se reúnen cientos de ellos y forman un falso ojo compuesto. Las piezas bucales tienen una estructura básica similar a la de los insectos (mandíbulas, primer par de maxilas y segundo par de maxilas o labio). Las mandíbulas están presentes en todos los grupos de miriápodos; el primer y segundo par de maxilas sufren diversas modificaciones en los diferentes grupos:
- Diplópodos y Paurópodos. Con mandíbulas; primer par de maxilas fusionadas originando un gnatoquilario; segundo par de maxilas ausente.
- Quilópodos. Con mandíbulas, primer y segundo par de maxilas fusionados. Primer par de patas modificado en grandes uñas (forcípulas) asociadas a una glándula venenosa, que usan para capturar las presas y por tanto, actúan como piezas bucales adicionales.
- Sínfilos. Con mandíbulas, primer par de maxilas separado y segundo par de maxilas fusionado para formar un labio.
El sistema circulatorio está formado por un corazón tubular dorsal que bombea hacia la cabeza la hemolinfa a través de la aorta. El intercambio de gases se realiza gracias a un sistema de tráqueas y espiráculos análogo al de los hexápodos. Poseen uno o dos pares de tubos de Malpighi derivados del proctodeo (ectodérmicos), con función excretora.
Biología y ecología

Los miriápodos son dioicos (sexos separados) y ovíparos, aunque hay casos de partenogénesis entre los diplópodos, quilópodos y sínfilos. La inseminación es en muchos casos indirecta; los machos depositan los espermatóforos en las inmediaciones de la hembra, que los recoge y almacena en el cuerpo.
La mayoría de los miriápodos son lucífugos (huyen de la luz) e higrófilos (buscan la humedad). Por ello, muchos son nocturnos, y se refugian durante el día bajo piedras, entre la hojarasca, en troncos en descomposición, entre el musgo, etc. Los quilópodos son depredadores agresivos y activos; el resto son principalmente herbívoros y muchos son detritívoros.
Filogenia
Posición de los miriápodos en el filo Arthropoda
Tradicionalmente los miriápodos han sido considerados parientes cercanos de los insectos (hexápodos), y que juntos formaban el subfilo Uniramia, dentro del cual los miriápodos son tratados como una superclase. Su condición fue cuestionada por algunos zoólogos (p. ej. Barnes, 1968[6]), y en particular fue considerado probable que fueran un grupo parafilético. Sin embargo, estudios genéticos apoyan su condición monofilética, y sugieren que pueden no ser parientes tan cercanos de los hexápodos, pudiendo estar más cerca de los quelicerados.[7]
Varios análisis moleculares recientes y también fósiles han demostrado que los parientes cercanos de los hexápodos son los crustáceos con quienes forman el clado Pancrustacea, este último es hermano de los miriápodos, la hipótesis de que los miriápodos y hexápodos comprenden grupos hermanos ha sido descartada en gran medida, por tanto las relaciones filogenéticas entre grupos de artrópodos serían las siguientes, incluyendo una posible filogenia interna de los miriápodos:[7][8][9][10][11][12]
| Arthropoda |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Filogenia interna de los miriápodos
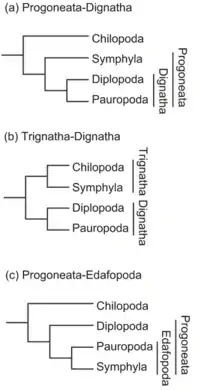
En cuanto a su filogenia interna, los datos morfológicos apoyaban la parafilia o la polifilia de los miriápodos respecto a los hexápodos (Wheeler, 1998[13] Los datos moleculares por su parte sugieren, la monofilia de los miriápodos al no estar estrechamente emparentados con los hexápodos: (Zrzavý et al.,[14]
Las relaciones filogenéticas dentro los miriápodos no están todavía resueltas, se han propuesto las siguientes relaciones para los grupos de miriápodos:[15]
Véase también
 Portal:Artrópodos. Contenido relacionado con Artrópodos.
Portal:Artrópodos. Contenido relacionado con Artrópodos.
Referencias
- Real Academia Española. «miriópodo». Diccionario de la lengua española (23.ª edición).
- Chapman, A. D., 2010. Numbers of Living Species in Australia and the World, 2nd edition. Australian Biodiversity Information Services ISBN (online) 9780642568618
- Marek, Paul E.; Buzatto, Bruno A.; Shear, William A.; Means, Jackson C.; Black, Dennis G.; Harvey, Mark S.; Rodriguez, Juanita (December 2021). "The first true millipede—1306 legs long". Scientific Reports. 11 (1): 23126. Bibcode:2021NatSR..1123126M. doi:10.1038/s41598-021-02447-0. ISSN 2045-2322. PMC 8677783. PMID 34916527. S2CID 245317751
- Martínez-Delclòs, X. 1996. El registro fósil de los insectos. Boln. Asoc. esp. Ent., 20(1-2):9-30 (resumen) Archivado el 23 de enero de 2009 en Wayback Machine.
- Scholtz, Gerhard (2008). «Scarab beetles at the interface of wheel invention in nature and culture?». Contributions to Zoology (National Museum of Natural History Naturalis) 77 (3). ISSN 1875-9866. Archivado desde el original el 21 de octubre de 2016. Consultado el 19 de enero de 2009.
- Barnes, R. D., 1968. Invertebrate Zoology. W. B. Saunders Co., Philadelphia. 743 p.
- Regier; Shultz, J. W.; Zwick, A.; Hussey, A.; Ball, B.; Wetzer, R.; Martin, J. W.; Cunningham, C. W. (2010). «Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences». Nature 463 (7284). pp. 1079-1084. Bibcode:2010Natur.463.1079R. PMID 20147900. doi:10.1038/nature08742.
- Jerome C. Regier, Jeffrey W. Shultz, Andreas Zwick, April Hussey, Bernard Ball, Regina Wetzer, Joel W. Martin & Clifford W. Cunningham (2010). «Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences». Nature 463 (7284): 1079-1083. PMID 20147900. doi:10.1038/nature08742.
- Todd H. Oakley, Joanna M. Wolfe, Annie R. Lindgren and Alexander K. Zaharoff (2013). «Phylotranscriptomics to bring the understudied into the fold: monophyletic ostracoda, fossil placement, and pancrustacean phylogeny». Molecular Biology and Evolution 30 (1): 215-233. PMID 22977117. doi:10.1093/molbev/mss216.
- Giribet, Gonzalo; Edgecombe, Gregory D. (17 de junio de 2019). «The Phylogeny and Evolutionary History of Arthropods». Current Biology (en inglés) 29 (12): R592-R602. ISSN 0960-9822. PMID 31211983. doi:10.1016/j.cub.2019.04.057. Consultado el 2 de junio de 2020.
- Hassanin, Alexandre (2006). «Phylogeny of Arthropod inferred from mitochondrial sequences: Strategies for limiting the misleading effects of multiple changes in pattern and rates of substitution». Molecular Phylogenetics and Evolution 38 (1): 100-116. PMID 16290034. doi:10.1016/j.ympev.2005.09.012. Archivado desde el original el 10 de enero de 2011. Consultado el 16 de abril de 2010.
- Regier, Jerome C.; Shultz, J. W.; Zwick, A.; Hussey, A.; Ball, B.; Wetzer, R.; Martin, J.W.; Cunningham, C. W. (2010). «Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences». Nature 463 (7284): 1079-1084. Bibcode:2010Natur.463.1079R. PMID 20147900. doi:10.1038/nature08742.
- Wheeler, W. C., 1998. Sampling, grounplans, total evidence and the systematics of arthropods. En: R. A. Fortey & R. H. Thomas (eds.): Arthropod Relationships: 87-96. Chapman & Hall, London.
- Zrzavý, J., Mihulka, S., Kepka, P., Bezdék, A. & Tietz, D., 1998. Phylogeny of Metazoa based on morphological and 18S ribosomal DNA evidence. Cladistics, 14(3): 249-285.
- Zwick, Andreas; Regier, Jerome C.; Zwickl, Derrick J.; Gadagkar, Sudhindra R. (2012). «Resolving Discrepancy between Nucleotides and Amino Acids in Deep-Level Arthropod Phylogenomics: Differentiating Serine Codons in 21-Amino-Acid Models». PLoS ONE 7 (11): e47450. Bibcode:2012PLoSO...747450Z. PMC 3502419. PMID 23185239. doi:10.1371/journal.pone.0047450.
Enlaces externos
 Wikimedia Commons alberga una categoría multimedia sobre Myriapoda.
Wikimedia Commons alberga una categoría multimedia sobre Myriapoda. Wikispecies tiene un artículo sobre Myriapoda.
Wikispecies tiene un artículo sobre Myriapoda.- Fotografías de miriápodos variados por Krister Hall Archivado el 4 de marzo de 2016 en Wayback Machine.
- 101 cosas que no sabías sobre los miriápodos
| Control de autoridades |
|
|---|