Plateosaurus
Plateosaurus (gr. "lagarto ancho") es un género extinto de dinosaurio plateosáurido que vivió durante el período Triásico, hace entre 214 a 204 millones de años, en lo que hoy es Europa. Plateosaurus es un sauropodomorfo basal, uno de los así llamados "prosaurópodos". Su nombre probablemente significa "lagarto ancho", traducido de manera errónea como "lagarto plano".[1] Se reconocen como válidas tres especies, la especie tipo desde 2019, Plateosaurus trossingensis[2] del Noriense tardío y el Rhaetiense, Plateosaurus longiceps de edad similar y el levemente más antiguo Plateosaurus gracilis del Noriano inferior. Sin embargo, otras especies han sido asignadas en el pasado, y no hay un consenso extendido sobre la taxonomía de las especies de los plateosáuridos. De igual manera, hay una plétora de sinónimos, nombres duplicados que no son válidos, a nivel de género. La especie tipo anterior, Plateosaurus engelhardti, fue cambiada por lo pobre y dudoso del material tipo.[2]
| Plateosaurus | ||
|---|---|---|
| Rango temporal: 214 Ma - 204 Ma Noriense a Rhaetiense (Triásico Superior) | ||
 Esqueletos montados de P. trossingensis GPIT "Skelett 2", consistente de dos especímenes de la Formación Trossingen, en el museo del Instituto de Geociencias (GPIT) de la Universidad Eberhard-Karls, Tubinga, Alemania. | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Sauropodomorpha | |
| Infraorden: | Prosauropoda | |
| (sin rango): | Plateosauria | |
| Familia: | Plateosauridae | |
| Género: |
Plateosaurus von Meyer, 1837 | |
| Especie tipo | ||
|
Plateosaurus trossingensis Frass, 1913 | ||
| Especies | ||
| ||
| Sinonimia | ||
| ||
Descubierto en 1834 por Johann Friedrich Engelhardt y descrito tres años más tarde por Hermann von Meyer, Plateosaurus fue el quinto género de dinosaurio nombrado que aún se considera válido. Aunque fue descrito antes de que Richard Owen nombrara formalmente a Dinosauria en 1842, no fue uno de los tres géneros usados por Owen para definir al grupo, dado que por entonces, era muy mal conocido y difícil de identificar como un dinosaurio. Se trata uno de los dinosaurios mejor conocidos por la ciencia: cerca de 100 esqueletos han sido encontrados, algunos de los cuales están casi completos. La abundancia de sus fósiles en Suavia, Alemania, ha llevado a que se le dé el apodo de Schwäbischer Lindwurm, en alemán, lindworm suavo.
Plateosaurus era un herbívoro bípedo con un largo cuello móvil, dientes afilados pero rechonchos para triturar plantas, poderosos miembros traseros, brazos cortos pero musculosos y manos prensiles con grandes garras en tres de sus dedos, posiblemente usadas para defenderse y alimentarse. De manera inusual para un dinosaurio, Plateosaurus mostraba una fuerte variación en su desarrollo, en vez de tener un tamaño adulto más o menos uniforme, los individuos completamente desarrollados variaban entre 4,8 a 10 metros de largo y pesaban entre 600 a 4000 kilogramos. Comúnmente, estos animales vivían al menos entre 12 a 20 años, pero su esperanza de vida máxima es desconocida.
A pesar de la gran cantidad y la excelente calidad del material fósil, Plateosaurus fue durante mucho tiempo uno de los dinosaurios menos comprendidos. Algunos investigadores propusieron teorías que más tarde se mostró que contradecían la evidencia geológica y paleontológica, pero estas se volvieron parte del imaginario popular. Desde 1980 la taxonomía, sus relaciones, la tafonomía, como sus restos quedaron enterrados y se fosilizaron, la biomecánica, como funcionaba su estructura ósea, y la paleobiología, circunstancias de la vida, de Plateosaurus han sido reexaminadas en detalle, lo que ha llevado a alterar la interpretación de la biología, postura y comportamiento del animal.
Descripción
.jpg.webp)
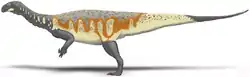
Plateosaurus tenía la forma corporal típica de un dinosaurio bípedo herbívoro, un cráneo pequeño, un cuello largo y flexible compuesto de 10 vértebras cervicales, un cuerpo robusto y una larga cola compuesta de al menos 40 vértebras caudales.[3][4][5] Los brazos de Plateosaurus eran muy cortos, incluso comparados con los de muchos otros "prosaurópodos". Aun así, poseían una constitución robusta, con manos adaptadas para tomar objetos.[4][6] La cintura escapular era estrecha, con frecuencia, se la muestra desalineada en las monturas y dibujos de esqueletos,[6] y sus clavículas terminaban tocándose en la línea media del cuerpo,[4] como en otros sauropodomorfos basales.[7] Los miembros posteriores se mantenían bajo el cuerpo, con las rodillas y tobillos levemente flexionados, y el pie era digitígrado, lo que significa que el animal caminaba apoyándose sobre sus dedos.[4][8][9] Las patas proporcionalmente largas y el metatarso muestran que Plateosaurus podía correr rápidamente con sus patas.[4][6][8][9] La cola de Plateosaurus era la típica de los dinosaurios, musculosa y con gran movilidad.[6]
El cráneo de Plateosaurus es pequeño y estrecho, rectangular al ser visto de lado, y casi tres veces más largo que alto. Tenía un foramen temporal lateral casi rectangular en la parte posterior. Las grandes órbitas oculares, la fenestra anteorbital de aspecto casi triangular y las narinas son casi del mismo tamaño.[3][4][10] Las mandíbulas tenían varios dientes pequeños en forma de hoja: de 5 a 6 en el premaxilar, 24 a 30 en el maxilar, y 21 a 28 en el dentario.[3][4][10] Las gruesas coronas romas de los dientes, que contaban con bordes aserrados le permitían triturar la materia vegetal.[3][4][10] La posición baja de la mandíbula le daba a los músculos masticatorios un efecto de palanca, de modo que Plateosaurus podría realizar una poderosa mordida.[10] Estos rasgos en conjunto sugieren que se alimentaba principal o exclusivamente de plantas.[10] Sus ojos apuntan directamente a los lados, más que al frente, proveyendo una visión periférica completa para vigilar a los posibles depredadores.[3][4][10] Algunos de los esqueletos fósiles han preservado los anillos escleróticos (los anillos de placas óseas que protegen el ojo).[3][4][10]

Las costillas se conectaban a las vértebras de la espalda con dos articulaciones, actuando juntas como una única bisagra, lo cual le ha permitido a los investigadores reconstruir las posiciones de inhalación y exhalación de la caja torácica. La diferencia de volumen entre ambas posiciones definen el volumen de aire intercambiado, la cantidad de aire movido en cada respiración, determinándose que serían aproximadamente 20 litros para un individuo de Plateosaurus con un peso estimado en 690 kilogramos, o 29 ml/kg de peso corporal.[6] Este es un valor típico para las aves, pero no para los mamíferos,[11] e indica que Plateosaurus probablemente tenía un sistema de circulación de aire en los pulmones similar al de las aves;[6] esto implicaría que tendría también una neumatización del esqueleto, con sacos aéreos del pulmón invadiendo los huesos que además permitían aligerar su peso. Los indicios de esta condición no fueron reconocidos sino hasta 2010 y solo se han hallado en los huesos de algunos individuos.[12][13] Combinado con la evidencia de la histología de los huesos[14][15] esto indica que Plateosaurus era endotermo.[15][16]
La especie tipo de Plateosaurus es P. trossingensis.[17] Los adultos de esta especie alcanzaban 4,8 a 10 metros de longitud,[15] y variaban en masa de 600 a 4000 kilogramos.[9] La especie geológicamente más antigua, P. gracilis, anteriormente denominada Sellosaurus gracilis, era algo menor, con una longitud total de 4 a 5 metros.[18]
Descubrimiento e investigación
Los fósiles de Plateosaurus gracilis, la especie más antigua, fueron hallados en la formación geológica de Löwenstein del Noriano inferior.[19] P. engelhardti data de la más reciente Formación Löwensteinde Noriano superiory por ultmo P. longiceps de la formación Trossingen, Noriano superior, y rocas de edades equivalentes.[20] Plateosaurus por lo tanto probablemente vivió aproximadamente entre 214 a 204 millones de años antes del presente.[21]

En 1834, el físico Johann Friedrich Engelhardt descubrió algunas vértebras y huesos de las patas en Heroldsberg cerca de Nuremberg, Alemania.[17] Tres años más tarde el paleontólogo Hermann von Meyer los designó como el espécimen holotipo de un nuevo género, Plateosaurus.[17] Desde entonces, los restos de cerca de 100 individuos de Plateosaurus han sido descubiertos en varios yacimientos europeos.[14]
El material asignado a Plateosaurus ha sido hallado en cerca de 50 localidades en Alemania, mayormente a lo largo de los valles de los ríos Neckar y Pegnitz, Suiza, en Frick y Francia.[23] Tres localidades son de especial importancia, debido a que han producido especímenes en grandes números e inusualmente, de buena calidad: cerca de Halberstadt en Sajonia-Anhalt, Trossingen en Baden-Württemberg, ambas de Alemania y Frick en Suiza.[14] Entre las décadas de 1910 a 1930, las excavaciones en una fosa de arcilla en Sajonia-Anhalt revelaron entre 39 a 50 esqueletos que pertenecían a Plateosaurus, junto con dientes y algunos huesos del terópodo Liliensternus, y dos esqueletos y fragmentos de la tortuga Proganochelys.[14] Parte del material de plateosáurido fue asignado a P. longiceps, una especie descrita por el paleontólogo Otto Jaekel en 1914.[24] La mayor parte del material encontrado allí fue enviando al Museo de Historia Natural de Berlín, en el que una cantidad importante de fósiles fue destruida durante la Segunda Guerra Mundial.[23] La cantera de Halberstadt quedó posteriormente cubierta por una urbanización.[23]

La segunda localidad principal alemana con hallazgos de P. engelhardti, una cantera en Trossingen en la Selva Negra, fue excavada repetidamente en el siglo XX.[23] Entre 1911 a 1932, las excavaciones durante seis salidas de campo lideradas por los paleontólogos alemanes Eberhard Fraas entre 1911 y 1912, Friedrich von Huene entre 1921 a 1923,[4][25] y finalmente Reinhold Seemann en 1932 revelaron un total de 35 esqueletos completos o parciales de Plateosaurus, así como restos fragmentarios de aproximadamente 70 individuos más.[23] El gran número de especímenes de Suavia causó que el paleontólogo alemán Friedrich August von Quenstedt apodara al animal Schwäbischer Lindwurm (lindworm suavo o dragón suavo).[26][27] [28][27] La mayor parte del material de Trossingen quedó también destruido en 1944, cuando el Naturaliensammlung en Stuttgart, predecesor del Museo Estatal de Historia Natural de Stuttgart, conocido con el aceonimo SMNS, fue incendiado tras una incursión de bombardeo de los Aliados. Afortunadamente, un estudio de 2011 del conservador del SMNS Rainer Schoch encontró por lo menos los hallazgos de la excavación de Seemann de 1932, en sus palabras «el material científico más valioso aún está disponible».[29]
Los esqueletos de Plateosaurus de la fosa de arcilla de Tonwerke Keller AG en Frick, Suiza, fueron reconocidos por primera vez en 1976.[14] Aunque los huesos están con frecuencia deformados de manera significativa por los procesos tafonómicos, Frick ha producido esqueletos de P. trossingensis comparables en integridad y posición a los de Trossingen y Halberstadt.[14] Martin Sander, paleontólogo de la Universidad de Bonn, indicó que el área se podría extender por 1,5 kilómetros, convirtiéndolo en el yacimiento de fósiles más grande de Europa. Se estima que se podrían encontrar los restos de un dinosaurio cada 100 metros cuadrados.[30]
En 1997, trabajadores de una plataforma petrolífera del campo petrolero de Snorre, localizada en el extremo norte del Mar del Norte, tropezaron con un fósil al que consideraron como material vegetal mientras taladraban a través de arenisca para la explotación petrolera. El núcleo de la perforación fue extraído de 2256 metros por debajo del suelo marino.[31] En 2003, el espécimen fue enviado a Jørn H. Hurum, paleontólogo en Universidad de Oslo para su estudio. Después de consultar a Martin Sander y Nicole Klein, paleontólogos de la Universidad de Bonn, se analizó la microestructura ósea y concluyeron que la roca preservaba tejido óseo fibroso de un fragmento de un hueso de las extremidades perteneciente a Plateosaurus,[31] convirtiéndolo en el primer dinosaurio hallado en Noruega y el hallado a mayor profundidad.[32] También se ha hallado material de Plateosaurus en la Formación Fleming Fjord del este de Groenlandia.[33]
Material tipo
El material tipo de Plateosaurus engelhardti incluye «aproximadamente 45 fragmentos óseos»,[20][34] de los cuales cerca de la mitad se han perdido.[35] El resto del material fue almacenado en el Instituto de Paleontología de la Universidad de Erlangen-Nuremberg, Alemania.[36] A partir de estos huesos, el paleontólogo alemán Markus Moser en 2003 seleccionó un sacro parcial, una serie de vértebras de la cadera fusionadas, como un lectotipo.[37] La localidad tipo no se conoce a ciencia cierta, pero Moser trató de inferirla a partir de publicaciones previas y el color y el tipo de preservación de los huesos. Concluyó que el material probablemente se recuperó de Buchenbühl, aproximadamente a 2 kilómetros al sur de Heroldsberg.[38] Sin embargo, el lectotipo de P. englehardti no se puede distinguir de P. longiceps o P. trossingensis.
El espécimen tipo de Plateosaurus gracilis, un postcráneo incompleto, es conservado en el Staatliches Museum für Naturkunde Stuttgart, Alemania.[39] La localidad tipo es Heslach, un suburbio de Stuttgart, Alemania.[39]
El espécimen tipo de Plateosaurus longiceps es MB R.1937, que está almacenado en el Museum für Naturkunde de Berlín. Su localidad tipo es Halberstadt, ubicada en Sajonia-Anhalt y el miembro Knollenmergel de la formación Trossingen.[40]
Etimología
La etimología del nombre Plateosaurus no es completamente clara. Moser señaló que la descripción original no contenía esa información, y varios autores han ofrecido diferentes interpretaciones.[1] El geólogo alemán Hanns Bruno Geinitz en 1846 lo llama πλατυς, breit, en español "ancho".[41] En el mismo año, Agassiz ofreció el griego antiguo πλατη, platê, "aleta", "timón", lo tradujo como el latín pala como "azada" o "remo" y σαυρος, sauros, "lagarto".[42][1] Agassiz por lo tanto renombró al género Platysaurus,[43][1] probablemente del griego πλατυς, platys, "ancho, plano, de espaldas anchas", creando un sinónimo más moderno inválido. Los autores posteriores frecuentemente se refieren a esta derivación, en especial al segundo significado que es "plano" de platys, por lo que Plateosaurus es traducido frecuentemente como "lagarto plano".
Con frecuencia, se ha afirmado que platys se supone que se referiría a los restos aplanados, por ejemplo los dientes lateralmente aplanados de Plateosaurus,[44] lo cual es imposible debido a que los dientes y los huesos aplanados como los pubis y algunos elementos craneales eran desconocidos en el momento de su descripción original. En 1855, von Meyer publicó una detallada descripción de Plateosaurus con ilustraciones, pero de nuevo no dio detalles de su etimología. Se refirió repetidamente a su tamaño gigantesco, "Riesensaurus" como lagarto gigante y a sus grandes extremidades, "schwerfüssig", comparando a Plateosaurus con los grandes mamíferos terrestres modernos, pero no describió ningún rasgo importante que encaje con el término «plano» o «en forma de remo».[45][46]
Además de sus descripciones científicas formales, von Meyer también dio una conferencia pública sobre reptiles fósiles en 1851 que incluyó una breve mención de Plateosaurus. Esa charla y una posterior sobre mamíferos fósiles se convirtieron en un libro para la audiencia general publicado en 1852, titulado "Über die Reptilien und Säugethiere der verschiedenen Zeiten der Erde", en castellano "Sobre los reptiles y mamíferos de los diferentes períodos de tiempo de la Tierra".[47] En el texto alemán, en la página 44, von Meyer describió brevemente Plateosaurus y mencionó que tenía "breite, starke Gliedmaassenknochen", que se traduce como "huesos de las extremidades anchos y fuertes". Porque su descripción original de Plateosaurus en 1837 destacó la similitud de los huesos de sus extremidades grandes con los de Megalosaurus e Iguanodon, un significado "lagarto ancho" para el nombre Plateosaurus para referirse en particular a sus huesos robustos de las extremidades parecería plausible.
Especies válidas

La historia taxonómica de Plateosaurus ha sido considerada como «larga y confusa», un «caótico enredo de nombres».[48] A partir de 2019, solo tres especies son universalmente aceptadas como válidas.[2] Las especies tipo P. trossingensis, P. longiceps y P. gracilis, previamente asignadas a su propio género Sellosaurus .[5][18] El paleontólogo británico Peter Galton mostró claramente que todo el material craneano procedente de Trossingen, Halberstadt y Frick pertenecen a una sola especie.[10] Moser llevó a cabo la más extensa y detallada investigación de todo el material de plateosáuridos de Alemania y Suiza, concluyendo que todo el material de Plateosaurus y la mayor parte del material de prosaurópodo de Keuper pertenecen a la misma especie que el material tipo de Plateosaurus engelhardti.[5] Sin embargo, esto es problemático debido al estado no diagnóstico del lectotipo.[49] Moser consideró a Sellosaurus como integrante del mismo género que Plateosaurus, pero no discutió si S. gracilis y P. engelhardti pertenecían a la misma especie.[50] El paleontólogo Adam Yates de la Universidad de Witwatersrand añadió nuevas dudas a la separación de ambos géneros; incluyó el material tipo de Sellosaurus gracilis en Plateosaurus como P. gracilis y reintrodujo el antiguo nombre Efraasia para parte del material que había sido asignado a Sellosaurus.[18] En 1926, von Huene ya había concluido que los dos géneros eran uno solo.[51]
Yates ha advertido que P. gracilis podría ser un metataxón, lo cual significa que no hay evidencia de que el material asignado a este sea monofilético, perteneciente a una sola especie, o que sea parafilético, perteneciente a varias especies.[19] Esto se debe a que el holotipo de P. (Sellosaurus) gracilis no tiene cráneo, y los otros especímenes consisten de cráneos y material que no es comparable con el holotipo, como para poder establecer si pertenecen al mismo taxón. Es por lo tanto posible que el material conocido contenga más especies pertenecientes a Plateosaurus.[52]
Algunos científicos consideran que otras especies también son válidas, por ejemplo, P. erlenbergensis y P.engelhardti.[53] Estas afirmaciones son problemáticas ya que tanto P. erlenbergensis como P. engelhardti tienen especímenes de tipo no diagnóstico.[49]
Especies no válidas

Todas las especies nombradas de Plateosaurus excepto la especie tipo y P. gracilis han terminado siendo sinónimos más modernos de la especie tipo o nombres inválidos.[5][18] Von Huene[54] prácticamente designó una nueva especie y a veces un nuevo género para cada uno de los hallazgos relativamente completos de Trossingen. En total nombró tres especies de Pachysaurus y siete de Plateosaurus y de Halberstadt, una especie de Gresslyosaurus y ocho de Plateosaurus.[14] Más tarde, invalidaría a varias de estas especies, pero siguió convencido de que había más de un género y más de una especie de Plateosaurus presente en ambas localidades. Jaekel también creyó que el material de Halberstadt incluía a varios dinosaurios plateosáuridos, así como a prosaurópodos no plateosáuridos.[24] La investigación sistemática de Galton redujo drásticamente el número de géneros y especies. Galton propuso la sinonimización de todo el material craneano,[10][55][56] y describió las diferencias entre los sintipos de P. engelhardti y el material de Trossingen, que refirió a P. longiceps.[57] Galton reconoció que P. trossingensis, P. fraasianus y P. integer eran idénticos a P. longiceps.[58] Markus Moser, sin embargo, mostró que P. longiceps es en sí mismo un sinónimo más moderno de P. engelhardti.[5] Más aún, una variedad de especies en otros géneros fueron creados para el material perteneciente a P. engelhardti, incluyendo a Dimodosaurus poligniensis, Gresslyosaurus robustus, Gresslyosaurus torgeri, Pachysaurus ajax, Pachysaurus giganteus, Pachysaurus magnus y Pachysaurus wetzelianus.[50] G. ingens se ha considerado independiente de Plateosaurus, pendiente de una revisión del material.
El cráneo de AMNH FARB 6810, hallado en Trossingen en 1925, es el mejor preservado de Plateosaurus a pesar de que fue hallado desarticulado y por lo tanto solo estaba disponible como huesos separados, fue descrito nuevamente en un artículo de 2011.[53] Los autores de esta publicación, los paleontólogos Albert Prieto-Márquez y Mark A. Norell, refirieron este cráneo a P. erlenbergensis, una especie erigida en 1905 por Friedrich von Huene. Si el holotipo de P. erlenbergensis es diagnóstico, es decir, que tiene suficientes rasgos para distinguirlo de otros materiales fósiles, entonces es el nombre correcto para el material asignado a P. longiceps Jaekel, 1913.[53] No obstante, de acuerdo con el último estudio detallado del material holotipo de P. engelhardti por Markus Moser, P. erlenbergensis es un sinónimo más moderno de P. engelhardti.[50]
Aparte de los fósiles claramente pertenecientes a Plateosaurus, hay varios materiales de prosaurópodos de Knollenmergel en colecciones de museos que están etiquetados como Plateosaurus, pero que no pertenecen a la especie tipo y posiblemente ni siquiera a Plateosaurus en absoluto.[59][60] Parte de este material no es diagnóstico; algunos otros de los fósiles han sido reconocidos como diferentes, pero no han sido descritos de forma adecuada.[61]
Especies dudosas o inválidas

Muchas especies de Plateosaurus son consideradas inválidas o dudosas debido a su naturaleza fragmentaria o a lo pobremente conservado de los restos. Especies como P. erlenbergiensis son dificultosas de determinar debido a que al holotipo le faltan el sacro, ilion y la parte proximal del pubis, huesos importantes para determinar su validez.[58]
- P. carinatus (Owen, 1854 (originalmente Massospondylus) = Massospondylus carinatus
- P. cullingworthi (Haughton, 1924) = Plateosauravus cullingworthi
- P. elisae (N.D.) (Sauvage, 1907) = P. ?engelhardti
- P. fraasianus (Huene, 1932) = P. longiceps
- P. ingens (N.D.) (Rütimeyer, 1856) (originalmente Gresslyosaurus) = Gresslyosaurus ingens
- P. integer (Fraas vide Huene, 1915 = P.longiceps
- P. poligniensis (?N.D.) (Pidancet & Chopard, 1862) (originalmente Dimodosaurus) = Dimodosaurus poligniensis
- P. quenstedti (N.D.) (Koken, 1900) (originalmente Zanclodon) = "Z." quenstedti
- P. robustus (N.D.) (Huene, 1907-08) (originalmente Gresslyosaurus) = "Gresslyosaurus" robustus
- P. torgeri (Jaekel, 1911) (originalmente Gresslyosaurus) = "G." torgeri
- P. trossingensis (Frass, 1914) = P. longiceps
- "P." ornatus (N.D.) (Huene, 1907-08) > Sauropodomorpha incertae sedis
- "P." plieningeri (N.D.) (Huene, 1907-08) > Sauropodomorpha incertae sedis
- "P." reinigeri (N.D.) (Huene, 1905) > Sauropodomorpha incertae sedis
- "P." stormbergensis (N.D.) (Broom, 1915) > Sauropodomorpha incertae sedis
Clasificación
El género Plateosaurus se encuadra en un grupo de dinosaurios herbívoros primitivos conocidos como "prosaurópodos".[54] El nombre de este grupo es obsoleto, ya que «Prosauropoda» no es una agrupación monofilética, motivo por el cual se le cita entre comillas, y muchos investigadores prefieren usar el término sauropodomorfo basal.[62][63] Plateosaurus fue el primer "prosaurópodo" en ser descrito.[54] Inicialmente, cuando el género era poco conocido, solo fue incluido en Sauria, siendo por tanto alguna clase de reptil, pero sin estar en ningún otro taxón definido más estrictamente.[17] En 1845, von Meyer creó el grupo Pachypodes, un sinónimo obsoleto de Dinosauria, para incluir a Plateosaurus, Iguanodon, Megalosaurus e Hylaeosaurus.[64] Posteriormente, Othniel Charles Marsh en 1895 propuso la familia Plateosauridae para incluir a Plateosaurus, dentro del infraorden Theropoda.[65][58] Von Huene propondría su traslado a "Prosauropoda",[66] una clasificación que fue aceptada por la mayoría de los paleontólogos.[5][18][67][68] Antes de la llegada de la cladística a la paleontología durante la década de 1980, con su énfasis en grupos monofiléticos, lamados clados, Plateosauridae estaba vagamente definida como un grupo de especies de sauropodomorfos caracterizadas por sus pies y manos anchos, con cráneos relativamente grandes, a diferencia de los pequeños "anquisáuridos" y los "melanorosáuridos" que eran más similares a los saurópodos.[69] La revaluación de los "prosaurópodos" a la luz de los nuevos métodos de análisis llevó a la reducción de la familia Plateosauridae. A lo largo del siglo XX, el clado solamente incluyó a Plateosaurus y a varios de los sinónimos más modernos de este, pero se consideró posteriormente incluir a otro género de sauropodomorfo, Sellosaurus[70] y posiblemente también a Unaysaurus.[71] No obstante, se considera muy probable que Sellosaurus sea en realidad otro sinónimo de Plateosaurus.[18]
Filogenia
Filogenia de los sauropodomorfos basales simplificada a partir del análisis de Yates, 2007.[72] Este es solo uno de los muchos cladogramas propuestos para los sauropodomorfos basales. Algunos investigadores no están de acuerdo con que los plateosáuridos fueran los ancestros directos de los saurópodos.

| Plateosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||
Tafonomía
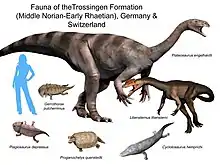
La tafonomía, el proceso de enterramiento y fosilización de los tres principales sitios de Plateosaurus , Trossingen, Halberstadt y Frick, es inusual por varias razones.[14] Los tres sitios son asociaciones monoespecíficas, lo que significa que contienen prácticamente a una sola especie, lo cual requiere de circunstancias muy especiales.[14] Sin embargo, se han hallado en los tres sitios dientes aislados de terópodos , así como restos de la tortuga primitiva Proganochelys.[14] Adicionalmente, un esqueleto parcial de "prosaurópodo" fue hallado en Halberstadt y no pertenece a Plateosaurus, pero está preservado en una posición similar.[24] Todos los sitios han producido esqueletos parciales y completos de Plateosaurus, así como huesos aislados.[14] Los esqueletos parciales tienden a incluir las extremidades posteriores y caderas, mientras que partes de la región anterior del cuerpo o el cuello son raramente hallados y aislados.[14] Los animales eran todos adultos o subadultos, casi completamente desarrollados, no se conocen juveniles o crías.[14] Los esqueletos completos y grandes partes del esqueleto que incluyen los miembros posteriores yacen en posición dorsal hacia arriba, como en las tortugas.[14] Además, en su mayoría están bien articulados, y los miembros posteriores están preservados en una postura de zigzag, frecuentemente con los pies más enterrados en el sedimento que las caderas.[14]
Primeras interpretaciones

En la primera discusión publicada de los hallazgos de Trossingen de Plateosaurus, Fraas en 1913 sugirió que solo el enterrarse en el lodo permitiría la preservación del único esqueleto conocido por entonces.[73] De manera similar, Jaekel interpretó que los hallazgos de Halberstadt corresponden a animales que vadeaban en pantanos demasiado profundos, que quedaron enterrados y terminaron por ahogarse.[24] Concluyó por tanto que los restos parciales fueron transportados al depósito por el agua, y refutó enfáticamente que fuera resultado de una acumulación catastrófica.[24] En contraste, von Huene interpretó el sedimento como depósitos eólicos, en que los animales más débiles, mayormente subadultos, sucumbieron a las duras condiciones de un desierto y terminaron hundiéndose en el lodo de pozas de agua temporales.[25] Von Huene afirmó que la integridad de muchos de los hallazgos indicaba que no fueron transportados, y concluyó que los individuos parciales y huesos aislados eran resultado del pisoteo y la intemperie.[25] Seemann desarrolló un escenario diferente, en el cual las manadas de Plateosaurus se congregaban en grandes pozas de agua, y algunos miembros de la manada eran empujados.[74] Los animales más ligeros conseguían liberarse, mientras que los individuos más pesados se atascaban y morían.[74]
Una escuela de pensamiento diferente se desarrolló casi un siglo después, con la sugerencia del paleontólogo David Weishampel de que los esqueletos de las capas inferiores contenían una manada que murió de manera catastrófica en una corriente de lodo, mientras que aquellos de las capas superiores se acumularon con el tiempo.[23] Weishampel explicó la curiosa asociación monoespecífica al teorizar que Plateosaurus era muy común en su época.[23] Esta teoría fue atribuida erróneamente a Seemann en un popular registro sobre los plateosáuridos en la colección del Instituto y Museo de Geología y Paleontología de la Universidad de Tubinga,[75] y desde entonces se ha vuelto la explicación estándar en muchos sitios de Internet y en libros populares sobre dinosaurios.[5] Rieber propuso un escenario más elaborado, el cual incluía que los animales murieron de sed o hambre, y quedaron concentrados en un solo punto debido a los flujos de lodo.[28]
Trampas de lodo
Una detallada revisión de la tafonomía realizada por el paleontólogo Martin Sander de la Universidad de Bonn, Alemania, encontró que la hipótesis del enterramiento en el lodo sugerida por primera vez por Fraas[73] es correcta.[14] Los animales que tenían o sobrepasaran cierto peso corporal se hundirían en el lodo, el cual se licuaba más a medida que los animales trataban de liberarse. De acuerdo con esto, el escenario de Sander, similar al propuesto para el conocido sitio fósil de Rancho La Brea, es el único que explica todos los datos tafonómicos: el grado de integridad de los cadáveres no fue influenciado por el transporte, lo cual es obvio por la carencia de indicios de transporte antes del enterramiento, sino más bien por el grado de consumo de los animales muertos por los carroñeros. Los juveniles de Plateosaurus y otros tipos de herbívoros eran demasiado ligeros como para quedar atrapados en el lodo o consiguieron liberarse por sí mismos, y por lo tanto no se preservaron. De igual forma, los terópodos carroñeros no quedaron atrapados debido a sus escaso peso corporal combinado con sus pies proporcionalmente grandes. No hay señal alguna de que los animales estuvieran reunidos en manada, o del catastrófico enterramiento de dicha manada, o de la súbita acumulación de animales que habían muerto previamente en algún otro lugar.[14]
Paleobiología
Postura y andadura
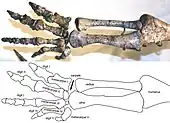
En la literatura científica se ha sugerido prácticamente cada postura imaginable en algún momento para Plateosaurus. Von Huene asumió que era un bípedo digitígrado con miembros posteriores erguidos, basándose en los animales que excavó en Trossingen, con la columna sostenida en un ángulo empinado (al menos durante una locomoción rápida).[4][76] En contraste, Jaekel, el principal investigador del material de Halberstadt, concluyó al principio que los animales caminaban de manera cuadrúpeda, y como los lagartos, tenía una postura extendida a los lados de sus extremidades, con pies plantígrados, y ondulando lateralmente el cuerpo al moverse.[77] Solo un año después, Jaekel empezó a favorecer una desmañada postura parecida a la de un canguro saltando,[3] un cambio de opinión por el cual fue ridiculizado por el zoólogo alemán Gustav Tornier,[78] quien interpretó la forma de las superficies de las articulaciones en las caderas y hombros como típicas de los reptiles. Fraas, el primer excavador del lagerstätte de Trossingen, también favoreció la postura reptiliana.[73][79] Müller-Stoll enumeró una serie de características requeridas para una postura con los miembros erguidos de las que supuestamente carecía Plateosaurus, concluyendo que las reconstrucciones basadas en lagartijas eran las correctas.[80] No obstante, muchas de están adaptaciones en realidad están presentes en Plateosaurus.[6][9] Desde 1980 en adelante, un mejor entendimiento de la biomecánica de los dinosaurios, y los estudios de los paleontólogos Andreas Christian y Holger Preuschoft sobre la resistencia a la flexión de la espalda de Plateosaurus,[8][81] llevó a la amplia aceptación de una postura erguida y digitígrada y de una posición aproximadamente horizontal de la espalda.[82][75][83][84][85][86] Muchos investigadores eran de la opinión de que Plateosaurus podía usar tanto una postura cuadrúpeda (para bajas velocidades) como bípeda (para una locomoción rápida),[8][81][84][87] y Wellnhofer insistió en que la cola se curvaba notablemente hacia abajo, haciendo que la postura bípeda fuera imposible.[86] Sin embargo, Moser mostró que de hecho la cola era recta.[88]

El consenso de "bípedo-cuadrúpedo" fue cambiado por un detallado estudio de los miembros delanteros de Plateosaurus por Bonnan y Senter (2007), el cual mostró claramente que Plateosaurus era incapaz de mantener sus manos en pronación.[89] Las manos en pronación en algunas monturas de esqueletos en museos se lograba al intercambiar la posición del radio y el cúbito en el codo. La carencia de pronación en el miembro delantero significa que Plateosaurus era exclusivamente bípedo (y por tanto incapaz de caminar de ninguna otra forma). Los indicadores adicionales de un modo de locomoción puramente bípedo son la gran diferencia de longitud entre las extremidades (las patas son aproximadamente el doble de largas que los brazos), el rango de movimiento muy limitado de los brazos, y el hecho de que el centro de masa descansa directamente sobre las extremidades posteriores.[6][9][90]
Plateosaurus muestra algunas adaptaciones propias de un corredor, incluyendo una postura erguida de los miembros posteriores, una parte inferior de la pata relativamente larga, un alargado metatarso y la postura digitígrada del pie.[9] Sin embargo, en contraste con los mamíferos corredores, el brazo de momento de los músculos extensores es corto, especialmente en el tobillo, en donde falta un distintivo tubérculo que incremente el brazo de momento en el calcáneo.[4] Esto significa que en contraste con los mamíferos corredores, Plateosaurus probablemente no realizaba zancadas con una fase aérea (sin apoyo en el suelo). En su lugar, Plateosaurus debió haber incrementado su velocidad al usar rápidas frecuencias de zancadas, creados por una rápida y poderosa retracción de las extremidades. El uso de la retracción en lugar de la extensión es típica entre los dinosaurios no avianos.[91]
Alimentación y dieta
Hasta la década de 1980, los paleontólogos seguían discutiendo la posibilidad de que los "prosaurópodos" tuvieran una alimentación carnívora.[92][93] Sin embargo, la hipótesis según la cual estos animales eran exclusivamente carnívoros se ha desacreditado, y todos los estudios recientes favorecen una forma de vida herbívora u omnívora para estos animales.[83]

Importantes características craneales (como la articulación de la mandíbula) de muchos «prosaurópodos» son más parecidas a las de los reptiles herbívoros que a los carnívoros, y la forma de la corona dental es parecida a la de las iguanas modernas herbívoras u omnívoras. La máxima anchura de la corona era mayor que la de la raíz de los dientes en muchos «prosaurópodos», incluyendo a Platosaurus; esto da como resultado un borde cortante similar al de los actuales reptiles herbívoros u omnívoros.[83]

Paul Barrett propuso que los prosaurópodos complementaban sus dietas herbívoras con presas pequeñas o carroña.[94] No se han encontrado fósiles de Plateosaurus con gastrolitos (piedras estomacales) en el área del estómago. La antigua y ampliamente citada idea de que todos los dinosaurios grandes (y de manera implícita también Plateosaurus) tragaban gastrolitos para ayudar a su digestión debido a su capacidad relativamente limitada para procesar la comida en la boca ha sido refutada por un estudio realizado por Oliver Wings sobre la abundancia, peso y superficie de los hallazgos fósiles de gastrolitos comparados con los aligátores y avestruces.[95][96] El uso de gastrolitos para la digestión parece haberse desarrollado más bien en la línea evolutiva que va de los terópodos basales a las aves, con un desarrollo paralelo en el ceratopsio Psittacosaurus.[96]
Crecimiento, metabolismo y esperanza de vida

Al igual que todos los dinosaurios no avianos que han sido estudiados, Plateosaurus crecía con un patrón de crecimiento diferente al de los actuales aves y mamíferos. En sus parientes cercanos, los saurópodos que tenían la típica fisiología dinosauriana, el crecimiento era inicialmente rápido, y continuaba algo más lentamente tras la madurez sexual, pero este era determinado; es decir, los animales paraban de crecer después de alcanzar un tamaño máximo.[97] Los mamíferos crecen rápidamente, pero su madurez sexual se alcanza generalmente al final de la etapa de rápido crecimiento. En ambos grupos, el tamaño final es relativamente constante, siendo los humanos atípicamente variables. Los reptiles modernos muestran un patrón de crecimiento parecido al de los saurópodos, inicialmente rápido, luego desacelerando tras la madurez sexual, y casi deteniéndose en una edad avanzada. Sin embargo, su tasa de crecimiento inicial es mucho más baja que la de los mamíferos, aves y dinosaurios. El crecimiento reptiliano es además muy variable, de modo que individuos de la misma edad pueden tener tamaños muy diferentes, y sus tamaños finales también varían significativamente. En las especies actuales, este patrón de crecimiento se vincula con la regulación del comportamiento por la termorregulación y una baja tasa metabólica (es decir, ectotermia), y es conocido como «plasticidad del desarrollo».[15][98]
Plateosaurus seguía una trayectoria de crecimiento parecida a la de los saurópodos, pero con una tasa de crecimiento y un tamaño final variables como en los reptiles actuales probablemente en respuesta a factores ambientales que podían afectar, por ejemplo, a la disponibilidad de comida. Algunos individuos alcanzaron su desarrollo total con solo 4.8 metros de longitud, mientras que otros alcanzaban los 10 metros. No obstante, la microestructura ósea indica un crecimiento rápido, como en los saurópodos y los mamíferos extintos, lo cual sugiere endotermia. Plateosaurus aparentemente representa una etapa inicial en el desarrollo de la endotermia, en la cual la misma estaba separada de la plasticidad en el desarrollo. Esta hipótesis se basa en un detallado estudio de la histología de los huesos largos de Plateosaurus llevado a cabo por Martin Sander y Nicole Klein de la Universidad de Bonn.[15] Un indicio adicional de endotermia es el tipo de pulmón de Plateosaurus que sería parecido al de las aves.[6]
La histología de los huesos largos también ha permitido estimar la edad que alcanzaba un individuo específico. Sander y Klein hallaron que algunos individuos estaban completamente desarrollados a los 12 años de edad, otros aún estaban creciendo lentamente a los 20 años, y un individuo aún estaba en la etapa rápida de crecimiento a los 18 años. El individuo más viejo encontrado tenía 27 años y seguía creciendo; muchos individuos se situaban entre los 12 a 20 años de edad.[15] No obstante, algunos pueden haber vivido mucho más, debido a que los fósiles de Frick y Trossingen son todos de animales que murieron en accidentes, y no por vejez. Debido a la ausencia de individuos de menos de 4.8 metros de longitud, no es posible deducir una serie ontogénica completa para Plateosaurus o determinar la tasa de crecimiento de animales de menos de 10 años de edad.[15]
Patrones de actividad diaria
Las comparaciones entre los anillos escleróticos y el tamaño estimado de la órbita ocular de Plateosaurus y especies de aves y reptiles modernos sugieren que puede haber sido catemeral, es decir activo durante el día y la noche, posiblemente evitando el calor del mediodía.[99]
Paleoecología
En algunas localidades se han hallado grupos de fósiles individuales completos, lo cual indica que las manadas viajaban juntas en el desierto árido del Triásico Superior de Europa, buscando alimentación y nuevos territorios. Vivieron junto a terópodos de tamaño mediano como Liliensternus y Halticosaurus, y el minúsculo Procompsognathus, entre otros contemporáneos se incluyen tortugas tempranas como Proganochelys, anfibios temnospóndilos, aetosaurios, mamíferos primitivos, pterosaurios, esfenodóntidos y peces.[100][101]
Referencias
- Moser, M., 2003, p. 13.
- «Opinion 2435 (Case 3560) – Plateosaurus Meyer, 1837 (Dinosauria, Sauropodomorpha): new type species designated». The Bulletin of Zoological Nomenclature 76 (1): 144-145. 2019. ISSN 0007-5167. doi:10.21805/bzn.v76.a042.
- Jaekel, O. (1911), Die Wirbeltiere. Eine Übersicht über die fossilen und lebenden Formen [The Vertebrates. An overview of the fossil and extant forms] (en alemán), Berlin: Borntraeger.
- Huene, F. von (1926). «Vollständige Osteologie eines Plateosauriden aus dem schwäbischen Keuper» [Complete osteology of a plateosaurid from the Swabian Keuper]. Geologische und Paläontologische Abhandlungen, Neue Folge (en alemán) 15 (2): 139-179.
- Moser, M. (2003). open access PDF «Plateosaurus engelhardti Meyer, 1837 (Dinosauria, Sauropodomorpha) aus dem Feuerletten (Mittelkeuper; Obertrias) von Bayern» [Plateosaurus engelhardti Meyer, 1837 (Dinosauria, Sauropodomorpha) from the Feuerletten (Mittelkeuper; Obertrias) of Bavaria]. Zitteliana Reihe B, Abhandlungen der Bayerischen Staatssammlung für Paläontologie und Geologie (en german with extended English summary) 24: 1-186. OCLC 54854853.
- Mallison, H. (2010). «The digital Plateosaurus II: an assessment of the range of motion of the limbs and vertebral column and of previous reconstructions using a digital skeletal mount». Acta Palaeontologica Polonica 55 (3): 433-458. doi:10.4202/app.2009.0075.
- Yates, A.M.; Vasconcelos, C.C. (2005). «Furcula-like clavicles in the prosauropod dinosaur Massospondylus». Journal of Vertebrate Paleontology 25 (2): 466-468. ISSN 0272-4634. JSTOR 4524460. doi:10.1671/0272-4634(2005)025[0466:FCITPD]2.0.CO;2.
- Christian, Andreas; Koberg, Dorothee; Preuschoft, Holger (1996). «Shape of the pelvis and posture of the hindlimbs in Plateosaurus». Palaeontologische Zeitschrift 70 (3–4): 591-601. doi:10.1007/BF02988095.
- Mallison, H. (2010). «The digital Plateosaurus I: body mass, mass distribution and posture assessed using CAD and CAE on a digitally mounted complete skeleton». Palaeontologia Electronica. 13.2.8A.
- Galton, Peter M. (1984). «Cranial anatomy of the prosauropod dinosaur Plateosaurus from the Knollenmergel (Middle Keuper, Upper Triassic) of Germany. I. Two complete skulls from Trossingen/Württ. With comments on the diet». Geologica et Palaeontologica 18: 139-171.
- Frappell, P.B.; Hinds, D.S.; Boggs, D.F. (2001). «Scaling of respiratory variables and the breathing pattern in birds: an allometric and phylogenetic approach». Physiological and Biochemical Zoology 74 (1): 75-89. PMID 11226016. doi:10.1086/319300.
- Wedel, M.J. (2007), «What pneumaticity tells us about 'prosauropods', and vice versa», en Barrett, P.M.; Batten, D.J., ed., Evolution and Palaeobiology of Early Sauropodomorph Dinosaurs (Special Papers in Palaeontology 77), Oxford: Blackwell Publishing, pp. 207-222, ISBN 978-1-4051-6933-2.
- Yates, A.M.; Wedel, M.J.; Bonnan, M.F. (2011). «The early evolution of postcranial skeletal pneumaticity in sauropodomorph dinosaurs». Acta Palaeontologica Polonica. advance online (accepted manuscript): 85. doi:10.4202/app.2010.0075. Archivado desde el original el 22 de mayo de 2011.
- Sander, P.M. (1992). «The Norian Plateosaurus bonebeds of central Europe and their taphonomy». Palaeogeography, Palaeoclimatology, Palaeoecology 93 (3–4): 255-299. doi:10.1016/0031-0182(92)90100-J.
- Sander, M.; Klein, N. (2005). «Developmental plasticity in the life history of a prosauropod dinosaur». Science 310 (5755): 1800-1802. Bibcode:2005Sci...310.1800S. PMID 16357257. doi:10.1126/science.1120125.
- Klein, N.; Sander, P.M. (2007), «Bone histology and growth of the prosauropod dinosaur Plateosaurus engelhardti von Meyer, 1837 from the Norian bonebeds of Trossingen (Germany) and Frick (Switzerland)», en Barrett, P.M.; Batten, D.J., ed., Evolution and Palaeobiology of Early Sauropodomorph Dinosaurs (Special Papers in Palaeontology 77), Oxford: Blackwell Publishing, pp. 169-206, ISBN 978-1-4051-6933-2.
- Meyer, H. von (1837). «Mitteilung an Prof. Bronn (Plateosaurus engelhardti» [message to Prof. Bronn (Plateosaurus engelhardti)]. Neues Jahrbuch für Geologie und Paläontologie (en alemán) 1837: 316.
- Yates, A.M. (2003). «Species taxonomy of the sauropodomorph dinosaurs from the Löwenstein Formation (Norian, Late Triassic) of Germany». Palaeontology 46 (2): 317-337. doi:10.1111/j.0031-0239.2003.00301.x.
- Yates, 2003, p. 331.
- Yates, 2003, p. 332.
- Deutsche Stratigraphische Kommission (Ed.). Stratigraphische Tabelle von Deutschland 2002 [mapa]. (2002) ISBN 3-00-010197-7. Consultado el 10 de diciembre de 2011.
- Moser, M., 2003, p. 8.
- Weishampel, D.B. (1984), «Trossingen: E. Fraas, F. von Huene, R. Seemann, and the "Schwäbische Lindwurm" Plateosaurus», Third Symposium on Terrestrial Ecosystems, Short Papers. (Reif, W.-E.; Westphal, F. edición), Tübingen: ATTEMPTO, pp. 249-253.
- Jaekel, O. (1913–1914). «Über die Wirbeltierfunde in der oberen Trias von Halberstadt» [About the vertebrate finds in the Upper Triassic of Halberstadt]. Paläontologische Zeitschrift (en alemán) 1: 155-215. doi:10.1007/BF03160336. Archivado desde el original el 28 de diciembre de 2013. Consultado el 7 de septiembre de 2014.
- Huene, F. von (1928). «Lebensbild des Saurischier-Vorkommens im obersten Keuper von Trossingen in Württemberg» [Pictures of the life of the Saurischia present in the upper-most Keuper of Trossingen in Württemberg]. Palaeobiologica (en alemán) 1: 103-116.
- Quenstedt, F.A. (1858), Der Jura [The Jurassic] (en alemán), Tübingen: H. Laupp'schen Buchhandlung.
- Sander, P. M., 1992, p. 255.
- Rieber, H. (1985). «Der Plateosaurier von Frick» [The plateosaur from Frick]. University Zürich (en alemán) 1985 (6): 3-4.
- Schoch, R.R. (2011). «Tracing Seemann's dinosaur excavation in the Upper Triassic of Trossingen: his field notes and the present status of the material». Palaeodiversity 4: 245-282. «p. 271 ».
- «Dinosaur mass grave discovered in Switzerland». CBC News (en inglés). 10 de agosto de 2007. Consultado el 22 de febrero de 2018.
- Hurum, J.H.; Bergan, M.; Müller, R.; Nystuen, J.P.; Klein, N. (2006). «A Late Triassic dinosaur bone, offshore Norway». Norwegian Journal of Geology 86: 117-123. Archivado desde el original el 29 de septiembre de 2006.
- Owen, James (26 de abril de 2006). «Oil Drillers Strike World's Deepest Dinosaur». National Geographic News News (en inglés). Consultado el 22 de febrero de 2018.
- Jenkins, F.A. Jr.; Shubin, N.H.; Amaral, W.W.; Gatesy, S.M.; Schaff, C.R.; Clemmensen, L.B.; Downs, W.R.; Davidson, A.R.; Bonde, N.; Osbaeck, F. (1994). «Late Triassic continental vertebrates and depositional environments of the Fleming Fjord Formation, Jameson Land, East Greenland». Meddelelser om Grønland, Geoscience 32: 1-25.
- Blankenhorn, M. (1898). «Saurierfunde im Fränkischen Keuper» [Saurian finds in the Frankonian Keuper]. Sitzungsberichte der Physikalisch-medicinischen Societät in Erlangen (en alemán). 29 (1897): 67-91. «p. 74 ».
- Moser, M., 2003, p. 12. La investigación de Moser da cuenta que los restos perdidos del material tipo de Plateosaurus identificados originalmente por Blanckenhorn en 1898, fueron vistos por última vez por von Huene en 1907, quien ilustró y describió algunos de ellos. Resumen en inglés, p. 170
- Moser, M., 2003, p. 13. "resumen en inglés en p. 161"
- Moser, M., 2003, p. 13. "véase también pp. 17 y pp. 36–40, resumen en inglés en pp. 160–161 y 163–164"
- Moser, M., 2003, p. 14-15. "resumen en inglés en p. 160"
- Yates, 2003, p. 331-332.
- «PBDB». paleobiodb.org. Consultado el 30 de noviembre de 2020.
- Geinitz, H.B. (1846), Grundriss der Versteinerungskunde [Basics of fossil lore] (en alemán), Dresden and Leipzig: Arnoldische Buchhandlung, p. 813, «p. 89 ».
- Agassiz, L. (1844), Nomenclator Zoologicus, continens nomina systernatica generum animalium tam viventium quam fossilium, secundum ordinem alphabeticum disposita, adjectis auctoribus, libris in quibus reperiuntur, anno editionis, etymologia et familiis, ad quas pertinent, in sirigulis classibus (1842–1846). Fasciculus VI continens Reptilia. (en latin), Soloduri: Jent & Gassmann, p. 34.
- Agassiz, L. (1846), [http://books.google.co.uk/books?id=AuIPAQAAMAAJ&dq=Nomenclatoris%20zoologici%20index%20universalis&pg=PA29 6#v=onepage&q&f=false Nomenclatoris zoologici index universalis, continens nomina systernatica classium, ordinum, familiarurn et generum animalium omnium, tam viventium quam fossilium, secundum ordinem alphabeticum unicum disposita, adjectis homonymiis, plantarum, nec non variis adnotationibus et emendationibus.] (en latin), Soloduri: Jent & Gassmann, p. 296.
- Vollrath, A. (1959), «Schichtenfolge (Stratigraphie)» [Layer sequence (stratigraphy)], Erläuterungen zur geologischen Karte von Stuttgart und Umgebung 1:50 000 [Explanations on the geological map of Stuttgart and surroundings 1:50 000], Freiburg: Geologisches Landesamt in Baden-Württemberg, pp. 27-89, OCLC 61816316, «p. 57 ».
- Meyer, H. von (1855), Zur Fauna der Vorwelt. Zweite Abtheilung. Die Saurier des Muschelkalks, mit Rücksicht auf die Saurier aus dem bunten Sandstein und Keuper. [On the fauna of ancient times. Second Part. The saurians of the Muschelkalk, with reference to the saurians from the colored Sandstone and Keuper.] (en alemán), Frankfurt am Main: H. Keller, pp. 1-167, «pp. 152-155 ».
- Creisler, B. (febrero de 2012). «Plateosaurus: the Etymology and Meaning of a Name». Dinosaur Mailing List, Archive. University of Southern California. Archivado desde el original el 4 de marzo de 2016. Consultado el 7 de septiembre de 2014.
- Meyer, H. von (1852). Über die Reptilien und Säugethiere der verschiedenen Zeiten der Erde. Zwei Reden von Hermann von Meyer. [On the Reptiles and Mammals from the Different Time Periods of the Earth. Two Talks by Hermann von Meyer.] (en alemán). Frankfurt am Main: M., S. Schmerber. pp. 1-150.
- Yates, 2003, p. 317.
- Galton, Peter M. (2012). «Case 3560 Plateosaurus engelhardti Meyer, 1837 (Dinosauria, Sauropodomorpha): proposed replacement of unidentifiable name-bearing type by a neotype». The Bulletin of Zoological Nomenclature 69 (3): 203-212. ISSN 0007-5167. doi:10.21805/bzn.v69i3.a15.
- Moser, M., 2003, p. 152. "resumen en inglés en p. 179"
- Huene, 1926, p. 5.
- Yates, 2003, p. 328.
- Prieto-Márquez, A.; Norell, Mark A. (2011). «Redescription of a nearly complete skull of Plateosaurus (Dinosauria: Sauropodomorpha) from the Late Triassic of Trossingen (Germany)». American Museum Novitates 3727 (3727): 1-58. doi:10.1206/3727.2.
- Huene, F. von (1932). «Die fossile Reptil-Ordnung Saurischia, ihre Entwicklung und Geschichte» [The fossil order of reptiles Saurischia, their development and history]. Monographien zur Geologie und Paläontologie (en alemán) 4: 1-361.
- Galton, Peter M. (1985). «Cranial anatomy of the prosauropod dinosaur Plateosaurus from the Knollenmergel (Middle Keuper, Upper Triassic) of Germany. II. All the cranial material and details of soft-part anatomy». Geologica et Palaeontologica 19: 119-159.
- Galton, Peter M. (1986). «Prosauropod dinosaur Plateosaurus (=Gresslyosaurus) (Saurischia: Sauropodomorpha) from the Upper Triassic of Switzerland». Geologica et Paleontologica 20: 167-183.
- Galton, Peter M. (2000). «The prosauropod dinosaur Plateosaurus Meyer, 1837 (Saurischia, Sauropodomorpha). I: The syntypes of P. engelhardti Meyer, 1837 (Upper Triassic, Germany), with notes on other European prosauropods with "distally straight" femora». Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 216 (2): 233-275.
- Galton, Peter M. (2001). «The prosauropod dinosaur Plateosaurus Meyer, 1837 (Saurischia: Sauropodomorpha; Upper Triassic). II. Notes on the referred species». Revue Paléobiologie, Genève 20 (2): 435-502.
- Weishampel, D.B.; Barrett, P.M.; Coria, R.A.; Loeuff, J.L.; Xing, X.; Xijin, Z.; Sahni, A.; Gomani, E.M.P.; Noto, C. (2004), Weishampel, D.B.; Dodson, P.; Osmólska, H., ed., The Dinosauria (2 edición), Berkeley: University of California Press, p. 524, ISBN 978-0-520-25408-4.
- Moser, M., 2003, p. 152. "resumen en inglés en p.179"
- Jaekel, 1913, p. 193-194.
- Yates, A.M. (2010). «A revision of the problematic sauropodomorph dinosaurs from Manchester, Connecticut and the status of Anchisaurus Marsh». Palaeontology 53 (4): 739-752. doi:10.1111/j.1475-4983.2010.00952.x.
- Rowe, T.B.; Sues, H.-D.; Reisz, R.R. (2011). «Dispersal and diversity in the earliest North American sauropodomorph dinosaurs, with a description of a new taxon». Proceedings of the Royal Society B 278 (1708): 1044-1053. PMC 3049036. PMID 20926438. doi:10.1098/rspb.2010.1867.
- Meyer, H. von (1845). «System der fossilen Saurier» [Taxonomy of fossil saurians]. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrfakten-Kunde (en alemán) 1845: 278-285.
- Marsh, O.C. (1895). «On the affinities and classification of the dinosaurian reptiles». American Journal of Science 50 (300): 483-498. doi:10.2475/ajs.s4-7.42.403.
- Huene, F. von (1926). «On several known and unknown reptiles of the order Saurischia from England and France». Annals and Magazine of Natural History 17 (101): 473-489. doi:10.1080/00222932608633437.
- Young, C.C. (1941). «A complete osteology of Lufengosaurus huenei Young (gen. et sp. nov.) from Lufeng, Yunnan, China». Palaeontologia Sinica, New Series C. 7 (Whole Series No. 121): 1-59.
- Bonaparte, J.F. (1971). «Los tetrápodos del sector superior de la Formación Los Colorados, La Rioja, Argentina (Triásico Superior)» [The tetrapods of the upper part of the Los Colorados Formation, La Rioja, Argentina (Upper Triassic)]. Opera Lilloana 22: 1-183.
- Galton, Peter M. (1976). «Prosauropod dinosaurs (Reptilia: Saurischia) of North America». Postilla 169. (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- Huene, F. von (1905). «Über die Trias-Dinosaurier Europas» [About the Triassic dinosaurs of Europe]. Zeitschrift der Deutschen Geologischen Gesellschaft (en alemán) 57: 345-349.
- Leal, L.A.; Azevedo, S.A.K.; Kellner, A.W.A.; Da Rosa, A.A.S. (2004). «A new early dinosaur (Sauropodomorpha) from the Caturrita Formation (Late Triassic), Paraná Basin, Brazil». Zootaxa 690: 1-24.
- Yates, A.M. (2007), «The first complete skull of the Triassic dinosaur Melanorosaurus Haughton (Sauropodomorpha: Anchisauria)», en Barrett, P.M.; Batten, D.J., ed., Evolution and Palaeobiology of Early Sauropodomorph Dinosaurs (Special Papers in Palaeontology 77), Oxford: Blackwell Publishing, pp. 9-55, ISBN 978-1-4051-6933-2.
- Fraas, Eberhard (1913). «Die neuesten Dinosaurierfunde in der schwäbischen Trias» [The latest dinosaur finds in the Swabian Triassic]. Naturwissenschaften (en alemán) 1 (45): 1097-1100. Bibcode:1913NW......1.1097F. doi:10.1007/BF01493265.
- Seemann, R. (1933). «Das Saurierlager in den Keupermergeln bei Trossingen» [The sauria lagerstätte in the Keuper marls near Trossingen]. Jahreshefte des Vereins für Vaterländische Naturkunde, Württemberg (en alemán) 89: 129-160.
- Weishampel, D.B.; Westphal, F., Die Plateosaurier von Trossingen. [The plateosaurs from Trossingen.] (en alemán), Tübingen: ATTEMPTO, pp. 1-27.
- Huene, F. von (1907–1908). «Die Dinosaurier der europäischen Triasformation mit Berücksichtigung der aussereuropäischen Vorkommnisse» [The dinosaurs of the European Triassic Formation, with consideration of non-European occurrences]. Geologische und Paläontologische Abhandlungen, Supplement-Band (en alemán) 1: 1-419. Link to full text only available in US
- Jaekel, O. (1910). «Die Fussstellung und Lebensweise der grossen Dinosaurier» [The foot posture and mode of life of the large dinosaurs]. Zeitschrift der Deutschen Geologischen Gesellschaft, Monatsberichte (en alemán) 62 (4): 270-277.
- Tornier, G. (1912). «[no title]». Zeitschrift der Deutschen Geologischen Gesellschaft, Monatsberichte (en alemán) 64 (1): 2-24.
- Fraas, Eberhard (1912). «Die schwäbischen Dinosaurier» [The Swabian dinosaurs]. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg (en alemán) 68: lxvi-lxvii.
- Müller-Stoll, H. (1935). «Studie zur biologischen Anatomie der Gattung Plateosaurus» [Study on the biological anatomy of the genus Plateosaurus]. Verhandlungen des Naturhistorisch-Medizinischen Vereins zu Heidelberg (en alemán) 18 (1): 16-30.
- Christian, Andreas; Preuschoft, Holger (1996). «Deducing the body posture of extinct large vertebrates from the shape of the vertebral column». Palaeontology 39: 801-812.
- Moser, M., 2003, p. 138-142. "resumen en inglés en p. 176
- Galton, Peter M.; Upchurch, Paul (2004), «Prosauropoda», en Weishampel, D.B.; Dodson, P.; Osmólska, H., ed., The Dinosauria (2 edición), Berkeley: University of California Press, pp. 232–258, ISBN 978-0-520-25408-4.
- Galton, Peter M. (1990), «Basal Sauropodomorpha-Prosauropoda», en Weishampel, D.B.; Dodson, P.; Osmólska, H., ed., The Dinosauria (1 edición), Berkeley: University of California Press, pp. 320-344, ISBN 0-520-06727-4.
- Paul, 1997, p. 145.
- Wellnhofer, P. (1994). «Prosauropod dinosaurs from the Feuerletten (Middle Norian) of Ellingen near Weissenburg in Bavaria. In "Second Georges Cuvier Symposium, Montbeliard (France)", 1992». Revue de Paléobiologie. special 7: 263-271.
- Paul, G.S. (1997), «Dinosaur models: the good, the bad, and using them to estimate the mass of dinosaurs», en Wolberg, D.L.; Stump, E.; Rosenberg, G., ed., Dinofest International: Proceedings of a Symposium held at Arizona State University, Philadelphia: Academy of Natural Sciences, pp. 129-154, ISBN 0-935868-94-1.
- Moser, M., 2003, p. 142-144. "resumen en inglés en pp. 176–177"
- Bonnan, Matthew; Senter, Phil (2007), «Were the basal sauropodomorph dinosaurs Plateosaurus and Massospondylus habitual quadrupeds?», en Barrett, P.M.; Batten, D.J., ed., Evolution and Palaeobiology of Early Sauropodomorph Dinosaurs (Special Papers in Palaeontology 77), Oxford: Blackwell Publishing, pp. 139-155, ISBN 978-1-4051-6933-2.
- Mallison, H. (2011), «Plateosaurus in 3D: How CAD Models and Kinetic–Dynamic Modeling Bring an Extinct Animal to Life», en Klein, N.; Remes, K.; Gee, C.; Sander M., ed., Biology of the Sauropod Dinosaurs: Understanding the life of giants, Life of the Past (series ed. Farlow, J.), Bloomington: Indiana University Press, pp. 219-236, ISBN 978-0-253-35508-9.
- Gatesy, S.M. (1990). «Caudofemoral musculature and the evolution of theropod locomotion». Paleobiology 16 (2): 170-186.
- Cooper, M.R. (1980). «The prosauropod dinosaur Massospondylus carinatus Owen from Zimbabwe: its biology, mode of life and phylogenetic significance». Occasional Papers of the National Museums and Monuments of Rhodesia, Series B, Natural Sciences 6 (10): 689-840.
- Attridge, J.; A.W. Crompton and Farish A. Jenkins, Jr. (1985). «The southern Liassic prosauropod Massospondylus discovered in North America». Journal of Vertebrate Paleontology 5 (2): 128-132.
- Barrett, Paul M. (2000), «Prosauropod dinosaurs and iguanas: Speculations on the diets of extinct reptiles», en Sues, H.-D., ed., Evolution of Herbivory in Terrestrial Vertebrates: Perspectives from the Fossil Record, Cambridge: Cambridge University Press, pp. 42–78, ISBN 0-521-59449-9.
- Wings, O. (2007). «A review of gastrolith function with implications for fossil vertebrates and a revised classification». Acta Palaeontologica Polonica 52 (1): 1-16.
- Wings, O.; Sander, P.M. (2007). «No gastric mill in sauropod dinosaurs: new evidence from analysis of gastrolith mass and function in ostriches». Proceedings of the Royal Society B 274 (1610): 635-640. PMC 2197205. PMID 17254987. doi:10.1098/rspb.2006.3763.
- Sander, P.M. (1999). «Life history of Tendaguru sauropods as inferred from long-bone histology». Mitteilungen des Museums für Naturkunde Berlin, Geowissenschaftliche Reihe 2: 103-112. doi:10.1002/mmng.1999.4860020107.
- Se debe señalar que no es lo mismo que la plasticidad del desarrollo neuronal.
- Schmitz, L.; Motani, R. (2011). «Nocturnality in dinosaurs inferred from scleral ring and orbit morphology». Science 332 (6030): 705-708. Bibcode:2011Sci...332..705S. PMID 21493820. doi:10.1126/science.1200043.
- Lucas, S. G., 1998. Global Triassic tetrapod biostratigraphy and biochronology. Palaeogeography, Palaeoclimatology, Palaeoecology 143: 347-384.
- Cox, Barry, R.J.G. Savage, and others. (1999). Encyclopedia of Dinosaurs & prehistoric creatures. 14 July 2007:pg 124.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikispecies tiene un artículo sobre Plateosaurus.
Wikispecies tiene un artículo sobre Plateosaurus. Wikimedia Commons alberga una categoría multimedia sobre Plateosaurus.
Wikimedia Commons alberga una categoría multimedia sobre Plateosaurus.
| Control de autoridades |
|
|---|
 Datos: Q131046
Datos: Q131046- Multimedia: Plateosaurus / Q131046
- Especies: Plateosaurus