Plesiorycteropus
El cerdo hormiguero malgache (Plesiorycteropus) es un género de mamífero euterio recientemente extinto de Madagascar. Es el único miembro de la familia Plesiorycteropodidae, probablemente integrado en los Tenrecomorpha, aunque desde su descripción, en 1895, había estado clasificado junto al cerdo hormiguero u oricteropo en el orden Tubulidentata, pero estudios recientes han hallado poca evidencia que los vincule a éstos. Hasta ahora se reconocen únicamente dos especies, P. madagascariensis, la de mayor talla, y P. germainepetterae, las cuales probablemente se superponían en su distribución, ya que se han hallado restos subfósiles de ambas en el mismo yacimiento.
| Cerdo hormiguero malgache | ||
|---|---|---|
| Rango temporal: Holoceno | ||
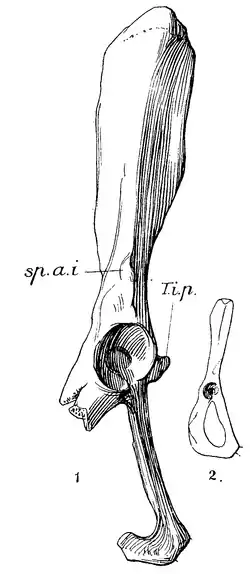 Huesos coxales derechos de la pelvis de Plesiorycteropus madagascariensis (1), comparados con los de una rata topera europea, Arvicola amphibius (2).[Note 1] | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Mammalia | |
| Infraclase: | Eutheria | |
| Orden: | Afrotheria | |
| Suborden: | ¿Tenrecomorpha? | |
| Familia: | Plesiorycteropodidae | |
| Género: |
Plesiorycteropus Filhol, 1895 | |
| Distribución | ||
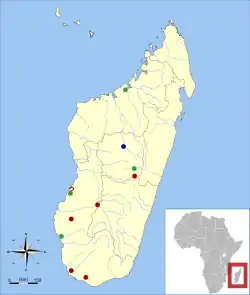 Sitios donde Plesiorycteropus ha sido hallado. Azul: P. madagascariensis y P. germainepetterae; verde: P. madagascariensis; rojo: Plesiorycteropus, especie incierta.[1] | ||
| Especies | ||
| ||
| Sinonimia | ||
El conocimiento de la anatomía de su esqueleto es limitado, ya que sólo se han recuperado a la fecha huesos de las extremidades, pelvis parciales y huesos del cráneo. Plesiorycteropus probablemente era un animal cavador que se alimentaba de insectos como las termitas y hormigas. También muestra adaptaciones para trepar y permanecer sentado. Estimaciones de su masa van de 6 a 18 kilogramos. Cuándo y porqué se extinguió es algo que sigue sin conocerse. Un hueso ha sido datado por radiocarbono en el año 200 antes de Cristo; la destrucción de los bosques por los humanos pudo haber contribuido a su extinción.
Taxonomía
Identificación y especies
El naturalista francés Henri Filhol describió por primera vez a Plesiorycteropus madagascariensis en 1895, a partir de un cráneo parcial hallado en una caverna de Belo. Su descripción fue vaga, incluso para los estándares del siglo XIX, pero lo clasificó como cercano al cerdo hormiguero (Orycteropus).[4] El nombre genérico combina el griego antiguo plesio- "cercano" y Orycteropus, el género del cerdo hormiguero, y el nombre de la especie se refiere a Madagascar. Charles Lamberton, quien tuvo acceso a una muestra mayor para su revisión del género en 1946, notó una variación considerable, pero no identificó especies distintas.[3] En 1994, Ross MacPhee revisó Plesiorycteropus de nuevo y fue capaz de determinar dos especies: P. madagascariensis y una nueva especie más pequeña, que nombró Plesiorycteropus germainepetterae por el científico Germaine Petter. Aparte de su tamaño, las dos especies difieren en varios aspectos morfológicos.[2]
Algunos restos de Plesiorycteropus han sido confundidos en ocasiones con roedores y primates. Así, Forsyth Major describió Myoryctes rapeto en 1908 como una «rata gigante subfósil» sobre la base de dos huesos pélvicos.[5] El nombre del género fue reemplazado por Majoria en 1915, debido a que Myoryctes ya había sido utilizado previamente (homonimia) para un género de gusano nemátodo.[6] Sin embargo, de acuerdo a MacPhee, la pelvis de Majoria era idéntica a la de Plesiorycteropus. Guillaume Grandidier asignó un fémur bien preservado a una nueva especie, pariente gigante del roedor actual votsovotsa (Hypogeomys antimena), a la que denominó Hypogeomys boulei. Posteriormente Lamberton identificó este fémur como perteneciente a Plesiorycteropus, en lo que MacPhee estuvo de acuerdo cuando revisó el género.[7] Los restos que habían sido asignados a Majoria rapeto y a Hypogeomys boulei están en el extremo superior del rango de tamaño del género, indicando que son referibles a la especie P. madagascariensis.[2] Otra pelvis de Plesiorycteropus fue asignada erróneamente a Daubentonia robusta, el extinto aye-aye gigante,[8] y otro material fue considerado como perteneciente a un lémur enano (Cheirogaleus).[9]
Relaciones
Filhol clasificó a Plesiorycteropus como cercano al oricteropo basándose en sus similitudes morfológicas. En su revisión de 1946, Charles Lamberton fue incapaz de proporcionar una asignación definitiva, confundido por las varias similitudes que vio entre cerdos hormigueros, pangolines, armadillos y osos hormigueros. Consideró que era probablemente un miembro primitivo y aislado de Edentata (sinonimia de Xenarthra), un grupo en el que, para Lamberton, además de a los verdaderos xenartros (perezosos, armadillos y osos hormigueros) incluía a cerdos hormigueros y pangolines. Rechazó otras alternativas, como una afinidad próxima a los cerdos hormigueros o la posibilidad de que el material asignado a Plesiorycteropus en realidad no representara a un único animal.[10] Bryan Patterson, quién en la década de 1970 revisó los tubulidentados (el orden al cual pertenecen los cerdos hormigueros y del que son sus únicos representantes vivos), aceptó a Plesiorycteropus como un miembro de este grupo, estimando como convergencias evolutivas muchos parecidos con los pangolines y otros animales.[11] Sin embargo, situó a este animal en su propia subfamilia, Plesiorycteropodinae, en vista de sus diferencias con otros tubulidentados (subfamilia Orycteropodinae), e hipotetizó que habrían llegado a Madagascar en el Eoceno, al mismo tiempo que los lémures.[12] Johannes Thewissen, quien criticó algunos aspectos de la clasificación de Patterson en 1985, también aceptó a Plesiorycteropus como un tubulidentado sin mayor comentario.[13]
Revaluando las contribuciones de Patterson y Thewissen en 1994, Ross MacPhee encontró en sus datos poco sustento para la clasificación de Plesiorycteropus como tubulidentado.[14] MacPhee usó el análisis cladístico para comprobar las relaciones del género con otros euterios, pero halló que diferentes variantes del análisis apoyaban distintas afinidades, pudiendo relacionarse con los oricteropos, damanes, ungulados (mamíferos con pezuñas), e incluso con los lipotiflos (musarañas, topos, erizos y parientes), por lo que no había evidencia convincente de que estuviera especialmente relacionado con cualquier otro grupo de euterios.[15] Por lo tanto, estableció un orden separado para Plesiorycteropus, al que denominó Bibymalagasia,[16] afirmando que podría ser inaceptable dejar a un mamífero reciente sin asignar a algún orden y que hasta el descubrimiento de material adicional, o análisis posteriores, era imposible demostrar afinidades cercanas de Plesiorycteropus con cualquier otro orden.[17] MacPhee consideró asimismo que era posible, aunque improbable, que unos cuantos taxones extintos, como Palaeorycteropus y Leptomanis del Paleógeno de Francia, pudieran ser considerados como bibymalagasios.[18] Varios estudios publicados por Robert Asher y colaboradores en 2003, 2005 y 2007, basándose en la morfología combinada con datos de secuencias de ADN en algunos análisis, produjeron distintas estimaciones de las relaciones de Plesiorycteropus, algunas situándolo dentro de Afrotheria como cercano a los oricteropos o a Afrosoricida, pero otros apoyando una relación con el erizo Erinaceus.[19] Un estudio morfológico de 2004 hecho por Inés Horovitz, centrándose en los ungulados suramericanos extintos (como Notoungulata y Litopterna), situó a Plesiorycteropus entre los tubulidentados y más cercano al extinto cerdo hormiguero Myorycteropus que al actual Orycteropus.[20] Un estudio de 2013 realizado por Michael Buckley examinó secuencias preservadas de colágeno en los huesos de Plesiorycteropus. Buckley halló que el animal estaba cercanamente relacionado con los tenrecs, un grupo de mamíferos insectívoros nativos de Madagascar, y sugirió que debía ser situado en el orden Tenrecoidea junto con los tenrecs y el topo dorado.[21]
Nombre común
El nombre "cerdo hormiguero malgache" ha sido usado como un nombre común para Plesiorycteropus, pero MacPhee lo consideró inapropiado debido a que el animal puede no estar relacionado con los cerdos hormigueros. En vez de éste, él propuso "bibymalagasio" como nombre común, una palabra elaborada del malgache que significa "animal de Madagascar".[3]
Descripción
Plesiorycteropus es conocido por algunos huesos subfósiles. El material incluye varios cráneos, todos los cuales carecen de huesos faciales, huesos largos completos como el fémur y el húmero (hueso superior del brazo), y otros huesos, pero algunos elementos son aún desconocidos, incluyendo la mayor parte del esqueleto de la mano y el pie.[22] Hay pocas razones para asumir que era distinto en forma al oricteropo.[23] No se han encontrado dientes o mandíbulas referibles a Plesiorycteropus, y en general se supone que el animal era desdentado.[24]
Basándose en la sección transversal de un fémur, MacPhee calculó estimaciones de la masa corporal. La menor estimación, basada en datos comparados de armadillos y pangolines, fue de 6 kilos para el fémur de menor tamaño (referible a P. germainepetterae) y la mayor estimación, basada en datos comparativos de roedores caviomorfos, fue 18 kilos para el mayor fémur disponible (P. madagascariensis); las estimaciones basadas en primates se situaron entre los dos extremos.[25] MacPhee supuso más correctas las estimaciones menores, porque estaban basadas en armadillos, que tienen fémures similares a los de Plesiorycteropus.[26] Por otro lado, el modelo basado en caviomorfos produjo un mejor estimado del tamaño del cerebro de Plesiorycteropus.[27]
Cráneo
Se conocen cuatro cráneos (tres de P. madagascariensis y uno de P. germainepetterae),[28] todos incompletos. Carecen del frente (rostro), y tres están rotos en el mismo lugar (en las cavidades paranasales, en el frente del neurocráneo), lo que sugiere que la parte frontal del cráneo era más delgada y frágil que la posterior, que consiste en huesos gruesos.[29] MacPhee estimó la máxima longitud craneal en P. madagascariensis en 101 milímetros. La longitud del hueso frontal promedia 35.4 milímetros en P. madagascariensis y unos 29.4 milímetros en P. germainepetterae.[30]
El robusto hueso nasal, preservado en un único espécimen, es ancho en el frente, un rasgo inusual entre los placentarios que es también visto en los armadillos, y es también inusualmente aplanado.[31] El laberinto etmoide, en la cavidad nasal, era grande, lo que sugiere que Plesiorycteropus tenía un buen sentido del olfato.[32] Una gran parte del tabique nasal, que separa las cavidades izquierda y derecha, está más osificada que lo usual en otros mamíferos; MacPhee pudo encontrar una condición similar sólo en los perezosos, los cuales tienen una nariz muy corta.[33] El hueso lacrimal es relativamente grande, tiene un único canal lagrimal, que se abre cerca de la sutura entre los huesos frontal y lacrimal, como en los lipotiflos, y posee un pequeño tubérculo cerca de esta abertura, ausente en los oricteropos.[34] La cuenca orbital, que aloja los ojos, es relativamente corta, similar a la de pangolines y armadillos.[35] Un distintivo tubérculo está presente sobre la sutura entre los huesos frontal y parietal en P. germainepetterae, pero no en P. madagascariensis.[36] P. madagascariensis tenía un neurocráneo más expandido y un estrechamiento menos pronunciado entre las órbitas.[2] Posee un foramen redondo mayor, una abertura en el huesos de la órbita. El canal óptico, que aloja los nervios que conducen a los ojos, es estrecho, lo que sugiere que tenía ojos pequeños.[37] Visto el cráneo desde arriba se puede ver poco del hueso escamoso, como en los pangolines y xenartros.[38] Las líneas temporales en el neurocráneo, en las que se insertan algunos músculos, están localizadas más bajas en P. germainepetterae.[2] Como en los oricteropos, los parietales son relativamente grandes. Presentan además un hueso interparietal. A diferencia de los osos hormigueros y pangolines, el occipucio (la parte trasera del cráneo) es plana y vertical. Plesiorycteropus carece de muescas sobre el foramen magnum (la abertura que conecta el cerebro con la médula espinal), las cuales están presentes en los oricteropos.[39] La cresta nucal, una proyección en el occipucio, es recta en P. madagascariensis, pero en P. germainepetterae ésta se interrumpe en el medio, similar a la situación en los armadillos y damanes.[40]
En sus descripciones de Plesiorycteropus, Lamberton y Patterson postularon distintas interpretaciones de la localización de la fosa mandibular, donde la mandíbula inferior se articula con el cráneo. MacPhee encontró problemas con estas interpretaciones y sugirió que la verdadera fosa mandibular era parte del área que Lamberton identificó como tal, en el lateral del neurocráneo. La fosa es pequeña y baja, lo que sugiere que el animal no era capaz de una mordida poderosa.[41] En la parte posterior de esta fosa hay un proceso pseudoglenoide,[42] más prominente en P. germainepetterae.[2] En P. germainepetterae se encuentra una pequeña abertura cerca del foramen oval, quizás el foramen vascular, no presente en P. madagascariensis.[2]
El hueso petroso forma una parte relativamente grande del techo de la cavidad timpánica, que aloja el oído medio;[43] partes del petroso están más desarrolladas en P. madagascariensis.[2]
Los moldes endocraneanos (del interior del cráneo) indican que la parte del neocórtex del cerebro era relativamente pequeña.[44]
Esqueleto postcraneal
Se conocen 34 vértebras de Plesiorycteropus. Estos animales tenían al menos siete vértebras sacras y cinco o seis lumbares. El hallazgo de vértebras caudales de la base de la cola, las cuales disminuyen en tamaño lentamente desde el frente hacia su término, sugieren que la cola era larga. No hay evidencia de articulaciones entre las vértebras, características de los xenartros.[45] En las siete vértebras torácicas (del tórax), las articulaciones con los disco intervertebrales tiene forma de fríjol y son mucho más anchas lateralmente que de adelante a atrás.[46] En el arco neural de las vértebras torácicas posteriores y en todas las lumbares hay un canal transarcual longitudinal.[47]
Una escápula (omóplato), tentativamente asignada a Plesiorycteropus, tenía el acromión, una apófisis del hueso, pero probablemente no tan grande como en los cerdos hormigueros o los armadillos. Se han hallado seis húmeros; el hueso es robusto[48] y presenta un foramen entepicondilar en el extremo distal.[49] Hay tres ejemplares del radio, un hueso compacto y macizo en Plesiorycteropus[50] que se parece al radio del pangolín.[51] Los tres cúbitos conocidos muestran que el proceso del olécranon en el extremo proximal está bien desarrollado, pero el extremo distal es estrecho; la morfología de este hueso sugiere que estos animales eran capaces de producir mucha fuerza con sus brazos.[52]
La pelvis es conocida de siete ejemplares, pero muchos están muy incompletos. Estos incluyen un estrecho iliaco y un largo isquion.[53] La tuberosidad isquiática, una parte del hueso estrecha y áspera en muchos placentarios, es amplia y suave en Plesiorycteropus.[54] Con 17 especímenes, el fémur es el hueso largo mejor representado. Se distingue en su largo cuello, de acuerdo a MacPhee similar sólo al de la rata lunar (Echinosorex).[55] Una proyección conocida como tercer trocánter es mayor en P. madagascariensis.[2] La tibia y el peroné se han fusionado en una tibiofíbula, de la cual se conocen ocho especímenes. Este hueso se parece al de los armadillos en su extensa fusión, la compresión del eje de la tibia, la estrechez de la superficie articular en el extremo distal y el amplio espacio entre los huesos.[56] A diferencia de los armadillos, la tibia y el peroné no están inclinados en relación con el otro, corriendo ambos en paralelo.[57] El astrágalo, del cual se conocen cuatro especímenes, es amplio y corto[58] y contiene un proceso posteromedial inusualmente grande.[47] Se conocen siete metápodos (huesos medios de la mano y el pie), variables en tamaño, pero MacPhee fue incapaz de separar los metacarpos (de la mano) de los metatarsos (del pie). Todos son cortos y anchos y son amplios proximalmente y estrechos distalmente.[59] Entre las pocas falanges conocidas, la falange proximal es más corta que la media y las falanges distales son estrechas y en forma de garra.[60]
Comportamiento, ecología, y extinción
| Sitio | ger. | mad. | sp. |
|---|---|---|---|
| Ambolisatra | + | + | |
| Ampasambazimba | + | + | + |
| Ampoza | + | ||
| Anjohibe | + | + | |
| Ankevo | ? | ||
| Anovaha | + | ||
| Antsirabe | + | + | |
| Belo | + | ||
| Bemifany | + | ||
| Masinandraina | + | ||
| Sirave | + | ||
Abreviaturas:
| |||
Los miembros delanteros de Plesiorycteropus muestran especializaciones para excavar arañando, colocando las patas delanteras contra el sustrato, clavando las garras, y luego desplazando las patas delanteras contra el cuerpo.[27] Otras partes del cuerpo también muestran dicha especialización, incluyendo grandes miembros posteriores y una cola ancha.[61] Algunos aspectos de la columna vertebral y la pelvis sugieren que estos animales adoptaban frecuentemente una postura erguida al sentarse.[62] Estos animales también pudieron haber sido capaces de trepar, quizás de manera similar a los gimnuros y a los tenrecs, los cuales tienen ojos pequeños como Plesiorycteropus.[63]
Eran probablemente mirmecófagos, devorando insectos como hormigas y termitas, pero también pudieron haber comido otros alimentos suaves, y dado su pequeño tamaño probablemente no se alimentaban en termiteros, como hacen los cerdos hormigueros.[64]
Plesiorycteropus y otros mamíferos recientemente extintos de la isla pueden haber vivido en y cerca de tierras húmedas. MacPhee tenía material de doce yacimientos del centro, oeste y sur de Madagascar.[65] P. madagascariensis es conocido de sitios a través de todo ese rango geográfico, pero P. germainepetterae ha sido registrado únicamente en el centro, aunque pequeños huesos del sur pudieran pertenecerle también. Por lo tanto las dos especies, al parecer, tenían una amplia superposición geográfica.[2]
Poco se sabe acerca de la extinción de Plesiorycteropus, pero MacPhee asumió que pudo haber ocurrido hace unos 1000 años, cuando se piensa que concluyó la extinción del resto de la fauna conocida solo por subfósiles de Madagascar. Nada similar a estos animales fue reportado por los exploradores europeos de la isla del siglo XVII,[66] y un hueso ha sido datado por radiocarbono alrededor del 2150 antes del presente.[67] Su extinción es algo anómala, ya que otros animales recientemente extintos de Madagascar como los grandes lémures, los hipopótamos malgaches, la fosa gigante y el ave elefante eran generalmente grandes y no insectívoros exclusivos; asimismo, algunas especies con dietas probablemente más especializadas, como la fanaloca (Eupleres goudoti) y el aye-aye (Daubentonia robusta), sobrevivieron.[66] Los primeros colonizadores humanos de Madagascar pueden haber causado la extinción de Plesiorycteropus a través de la destrucción del bosque y otras alteraciones.[23]
Notas
- El espécimen figurado es el M 7085 (Museo de Historia Natural de Londres), que fue descrito por Forsyth Major en 1908 como holotipo de Myoryctes rapeto (ahora sinionimia de Plesiorycteropus madagascariensis).
- homonimia.[3]
- Nomen novum (nombre de reemplazo) para Myoryctes Forsyth Major.[3]
Referencias
- MacPhee, 1994, tabla 1
- MacPhee, 1994, p. 32
- MacPhee, 1994, p. 30
- MacPhee, 1994, p. 11
- Forsyth Major, 1908, p. 97
- Thomas, 1915, p. 58
- MacPhee, 1994, p. 33
- MacPhee, 1994, pp. 33–34
- Godfrey et al., 2001, p. 71
- MacPhee, 1994, pp. 13–14
- Patterson, 1975, p. 215
- Patterson, 1975, p. 216
- MacPhee, 1994, p. 24
- MacPhee, 1994, pp. 23–24
- MacPhee, 1994, pp. 198–199
- MacPhee, 1994, p. 201
- MacPhee, 1994, pp. 202–203
- MacPhee, 1994, p. 201; Tabuce et al., 2008, p. 6, para la edad de los fósiles.
- Asher et al., 2003, p. 148; 2005, p. 919; Asher, 2007, figs. 1–2
- Horovitz, 2004, fig. 1
- Buckley, 2013, p. 1
- MacPhee, 1994, pp. 29–30
- Walker, 1999, p. 1050
- MacPhee, 1994, p. 13; Nowak, 1999, p. 1050
- MacPhee, 1994, table 14
- MacPhee, 1994, p. 147
- MacPhee, 1994, p. 148
- MacPhee, 1994, tabla 6
- MacPhee, 1994, p. 35
- MacPhee, 1994, table 7
- MacPhee, 1994, p. 40
- MacPhee, 1994, p. 42
- MacPhee, 1994, p. 43
- MacPhee, 1994, p. 41
- MacPhee, 1994, p. 44
- MacPhee, 1994, p. 51
- MacPhee, 1994, p. 55
- MacPhee, 1994, p. 78
- MacPhee, 1994, p. 79
- MacPhee, 1994, pp. 32, 79
- MacPhee, 1994, pp. 57, 59, 66
- MacPhee, 1994, p. 66
- MacPhee, 1994, p. 73
- MacPhee, 1994, p. 81
- MacPhee, 1994, p. 83
- MacPhee, 1994, p. 84
- MacPhee, 1994, p. 31
- MacPhee, 1994, p. 102
- MacPhee, 1994, p. 105
- MacPhee, 1994, p. 108
- MacPhee, 1994, p. 111
- MacPhee, 1994, p. 113
- MacPhee, 1994, p. 117
- MacPhee, 1994, p. 124
- MacPhee, 1994, p. 126
- MacPhee, 1994, p. 132
- MacPhee, 1994, p. 135
- MacPhee, 1994, p. 138
- MacPhee, 1994, p. 142
- MacPhee, 1994, pp. 146–147
- MacPhee, 1994, p. 149
- MacPhee, 1994, pp. 153, 156
- MacPhee, 1994, p. 157
- MacPhee, 1994, pp. 158–159
- MacPhee, 1994, pp. 34–35
- MacPhee, 1994, p. 159
- Burney et al., 2004, p. 54
Bibliografía
- Asher, R.J. 2007. A web-database of mammalian morphology and a reanalysis of placental phylogeny. BMC Evolutionary Biology 7:108.
- Asher, R.J., Novacek, M.J. and Geisler, J.H. 2003. Relationships of endemic African mammals and their fossil relatives based on morphological and molecular evidence (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última). (requiere suscripción). Journal of Mammalian Evolution 10(1–2):131–194.
- Asher, R.J., Emry, R.J. and McKenna, M.C. 2005. New material of Centetodon (Mammalia, Lipotyphla) and the importance of (missing) DNA sequences in systematic paleontology (requiere suscripción). Journal of Vertebrate Paleontology 25(4):911–923.
- Buckley, M. 2013. A molecular phylogeny of Plesiorycteropus reassigns the extinct mammalian order 'Bibymalagasia'. PLoS ONE 8(3):e59614.
- Burney, D.A., Burney, L.P., Godfrey, L.R., Jungers, W.L., Goodman, S.M., Wright, H.T. and Jull, A.J.T. 2004. A chronology for late prehistoric Madagascar (requiere suscripción). Journal of Human Evolution 47:25–63.
- Forsyth Major, C.I. 1908. A giant sub-fossil rat from Madagascar, Myoryctes rapeto gen. et sp. nov. (subscription required). Geological Magazine (5)5:97–98.
- Godfrey, L.R., Atkinson, M. and Semprebon, G.M. 2001. Paul Methuen's sleeping treasure: Subfossil lemurs in the zoological collections of the University Museum, Oxford (subscription required). American Journal of Physical Anthropology Suppl. 32:70–71.
- Horovitz, I. 2004. Eutherian mammal systematics and the origins of South American ungulates as based on postcranial osteology (requiere suscripción). Bulletin of Carnegie Museum of Natural History 36:63–79.
- MacPhee, R.D.E. 1994. Morphology, adaptations, and relationships of Plesiorycteropus, and a diagnosis of a new order of eutherian mammals. Bulletin of the American Museum of Natural History 220:1–214.
- Nowak, R.M. 1999. Walker's Mammals of the World. Volume 1. Johns Hopkins University Press, 1936 pp. ISBN 978-0-8018-5789-8
- Patterson, B. 1975. The fossil aardvarks (Mammalia: Tubulidentata). Bulletin of the Museum of Comparative Zoology at Harvard University 147:185–237.
- Tabuce, R., Asher, R.J. and Lehmann, T. 2008. Afrotherian mammals: a review of current data (requiere suscripción). Mammalia 72:2–14.
- Thomas, O. 1915. Notes on the Asiatic bamboo-rats (Rhizomys, etc.). Annals and Magazine of Natural History (8)16:56–61.
Enlaces externos
| Control de autoridades |
|
|---|
 Datos: Q135588
Datos: Q135588 Multimedia: Plesiorycteropus / Q135588
Multimedia: Plesiorycteropus / Q135588 Especies: Plesiorycteropus
Especies: Plesiorycteropus