Proteína transmembranal
Una proteína transmembranal es aquella proteína integral de membrana que atraviesa la bicapa lipídica de la membrana celular, una vez (unipaso) o varias (multipaso). Se pueden distinguir tres dominios. En primer lugar una zona que pasa a través de la membrana, de características hidrofóbicas (que repele el agua) para interactuar con los lípidos de la bicapa lipídica (membrana celular). Además, un dominio citosólico y otro extracitosólico en contacto con el interior y exterior de la célula respectivamente, de carácter hidrofílicos ("son afines al agua"). El resultado es una proteína anfipática.
Como proteína integral que es, solo se puede aislar de la bicapa mediante la aplicación de detergentes, como SDS. Las principales funciones son servir como canales transportadores de iones o moléculas, como por ejemplo las acuaporinas que transportan agua a través de la membrana; recepción de señales celulares, anclaje al citoesqueleto o a la matriz extracelular, etc.
Tipos de proteínas transmembranales según su estructura/
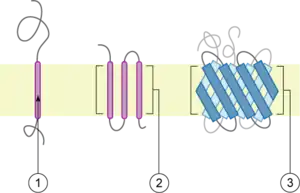
Transmembranal unipasos
Atraviesa la bicapa una sola vez. El extremo N-terminal de la proteína queda a un lado de la membrana, y el extremo C-terminal al otro. De este tipo son los Receptores-PTK (con actividad Proteintirosín kinasa), enzimas del Aparato de Golgi, etc. Un ejemplo muy estudiado es la glicoforina de la membrana del eritrocito. Los dominios de estas proteínas se clasifican como:
- a) Dominio transmembrana
- Presenta una estructura secundaria en α-hélice, con una longitud de unos 25 a 30 aminoácidos. Los residuos laterales han de ser en su mayoría hidrofóbicos, como alanina, leucina, isoleucina, etc. para poder interaccionar con el interior de la bicapa por fuerzas hidrofóbicas y de Van der Waals. Es importante señalar que el interior de la hélice está ocupado, y no sirve como canal. En los extremos de la hélice, pueden estar residuos con carga, que interaccionan con la cabeza polar de los fosfolípidos de la bicapa.
- b) Dominios no transmembrana
- Son generalmente hidrofílicos y globulares, de longitud variable, que se mantienen mediante enlaces de hidrógeno y fuerzas electrostáticas. En el dominio extracitosólico pueden presentarse enlaces disulfuro -S-S- y cadenas de oligosacáridos, cosa que no ocurre en el dominio citosólico.
Transmembranal multipaso
Atraviesan la bicapa lipídica en dos o más ocasiones, normalmente mediante varias α-hélice, aunque hay casos de inserciones a través de β-lámina. Hay muchos ejemplos: receptores asociados a proteínas G triméricas, canales iónicos, porinas en bacterias, y de nuevo en el eritrocito la proteína Banda 3, proteína transmembranal multipaso que atraviesa la membrana con 12 α-hélices, y con sus extremos hacia el citosol.
- a) Multipaso en α-hélice
- Estas proteínas tienen tantas α-hélices como veces atraviesan la membrana. Estas hélices se unen mediante bucles en sus dominios citosólico y extracitosólico. Las α-hélices pueden servir únicamente como mero medio de anclaje a la bicapa, o pueden formar un canal por el que puedan pasar diversas sustancias. En este último caso, las hélices tienen residuos hidrofóbicos que dan por fuera del canal, para interaccionar con la bicapa; mientras que los residuos que dan hacia el canal interno son hidrofílicos. De esta forma, muchas sustancias polares que en ausencia de proteínas no podían cruzar la membrana, ahora sí pueden hacerlo. Para poder formar un canal iónico, se necesita un mínimo de 5 α-hélices.
- Las α-hélice pueden desplazarse, resbalando unas sobre otras, para producir un cambio conformacional. Esto sirve para regular la abertura de canales, transportar compuestos por permeasas y para la transducción de señales
- b) Multipaso en β-lámina
- Mucho menos frecuentes son los casos de proteínas transmembrana multipaso en β-lámina. Ejemplos se dan en cloroplastos, mitocondrias y bacterias. La β-lámina sólo requiere unos 10 aminoácidos para atravesar la bicapa. Existen proteínas con un número de β-lámina desde 8 a 22.
- Normalmente forman canales por los cuales atraviesan diferentes solutos. Es el caso de los poros de porina, una proteína presente en la membrana de algunas bacterias, así como en mitocondrias y cloroplastos, por los que pasan sustancias con un peso molecular inferior a 800 Da por simple difusión. La porina cruza la membrana con 16 β-lámina antiparalelas, que simulan las duelas de un barril. Estas láminas alternan aminoácidos polares y apolares, los polares están orientados a la luz del canal, y los apolares contactan con la parte hidrófoba de la bicapa lipídica. Esto es una diferencia respecto a las α-hélice. Los bucles entre las β-lámina se proyectan hacia el lumen del canal, lo que les confiere especificidad. Además de porina, otros ejemplos son los poros también en forma de barril de maltoporina, para maltosa, y poro de proteína FepA, para iones de hierro, ambos en bacterias.
- No obstante, no todas las proteínas multipaso en β-lámina actúan como canales. Existen enzimas en β-lámina, como la lipasa OMPLA, y algunos receptores de membrana. En estos casos, los residuos polares de las β-lámina obstruyen el canal, imposibilitando esta función. La única función del barril es el anclaje a la membrana.
- A diferencia de las α-hélice, las β-lámina se unen entre sí de una forma más rígida, impidiendo los cambios conformacionales, limitándose así su versatilidad.
Referencias
- Alberts et al (2004). Biología molecular de la célula. Barcelona: Omega. ISBN 54-282-1351-8.
- Lodish et al. (2005). Biología celular y molecular. Buenos Aires: Médica Panamericana. ISBN 950-06-1974-3.
Véase también
| Control de autoridades |
|
|---|
 Datos: Q424204
Datos: Q424204 Multimedia: Transmembrane proteins / Q424204
Multimedia: Transmembrane proteins / Q424204