Hélice alfa
Las hélices alfa son estructuras secundarias de las proteínas. Esta hélice mantiene su forma por la formación de puentes de hidrógeno entre los átomos de oxígeno del grupo carbonilo de un aminoácido y el átomo de hidrógeno del grupo amino de otro aminoácido situado a cuatro aminoácidos de distancia en la cadena. Los grupos R se extienden hacia afuera de la hélice. Es una estructura anfipática porque posee una parte hidrofílica y una parte hidrófoba, lo que produce el enrollamiento de esta estructura, de manera que la parte hidrófoba no interactúe con el agua.[1]
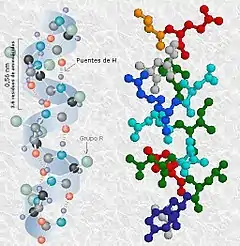
En las proteínas, la hélice α es el principal motivo de estructura secundaria. Fue postulada primero por Linus Pauling, Robert Corey, y Herman Branson en 1953 basándose en las estructuras cristalográficas entonces conocidas de aminoácidos y péptidos y en la predicción de Pauling de la forma planar de los enlaces peptídicos.
Descripción
Los aminoácidos en una hélice α están dispuestos en una estructura helicoidal dextrógira, con unos 3,6 aminoácidos por vuelta. Cada aminoácido supone un giro de unos 100° en la hélice, y los Cα de dos aminoácidos contiguos están separados por 1,5Å. La hélice está estrechamente empaquetada; de forma que no hay casi espacio libre dentro de la hélice. Todas las cadenas laterales de los aminoácidos están dispuestas hacia el exterior de la hélice.
El grupo N-H del aminoácido (n) puede establecer un enlace de hidrógeno con el grupo C=O del aminoácido (n+4). De esta forma, cada aminoácido (n) de la hélice forma dos puentes de hidrógeno con su enlace peptídico y el enlace peptídico del aminoácido en (n+4) y en (n-4). En total son 7 enlaces de hidrógeno por vuelta. Esto estabiliza enormemente la hélice. Está dentro de los niveles de organización de la proteína.
Estabilidad
Los cuatro primeros aminoácidos de la hélice, tal conocida como alfa, y los cuatro últimos solo podrán formar un enlace de hidrógeno en vez de dos, por lo tanto la hélice α suele ser más estable en la zona central que en los extremos. Para compensar esta pérdida, los aminoácidos de los extremos suelen ser polares y forman puentes de H con sus cadenas laterales y la cadena lateral de otros aminoácidos de la hélice. Cuando dos hélices alfa se aproximan entre sí tienden a interaccionar con ángulos de -30 y 60o.
En la hélice los momentos dipolares de todos los aminoácidos están perfectamente alineados, con lo que se forma un dipolo total con una carga parcial positiva en el extremo N-terminal y una carga parcial negativa en el extremo C-terminal.
En una hélice α, las cadenas laterales de los aminoácidos en posición (n) y en posición (n+4) quedan alineados. De forma que si en esas posiciones ponemos dos aminoácidos con carga de igual signo o muy voluminosos se desestabiliza la hélice.
Algunos aminoácidos, llamados disruptores de hélices, pueden desestabilizar la estructura helicoidal. Uno de ellos es la prolina, que al ser un iminoácido (aunque algunos autores cuestionan que la prolina no es en rigor un iminoácido), el N de su enlace peptídico no tiene unido un H para formar un enlace de hidrógeno con el aminoácido en (n+4). Además, el metileno unido al N del enlace peptídico también provoca impedimentos estéricos que hacen que la hélice tienda a romperse en el punto donde esté la prolina, aunque no lo hará si esta es suficientemente larga y estable. La glicina al proporcionar una gran flexibilidad, puesto que su cadena lateral es solo un H, suele estar en los acodamientos al final de la hélice.
Al primer aminoácido de una hélice en el extremo N-terminal se le llama N-cap y al último aminoácido de la hélice, en el extremo C-terminal se le llama C-cap. En posición N-cap, suelen aparecer aminoácidos polares no cargados, como la asparagina, o cargados negativamente, como el ácido glutámico, de forma que se compense la pérdida de un enlace peptídico en los extremos de la hélice que ya hemos comentado y en el caso del glutámico, la carga negativa de su cadena lateral interacciona con la carga parcial positiva del extremo N-terminal de la hélice.
En el C-cap son frecuentes la glicina y la prolina, que como ya hemos comentado rompen la estructura de la hélice, y también aminoácidos cargados positivamente, como la lisina, cuya carga positiva interacciona con la carga parcial negativa del extremo C-terminal de la hélice.
Los polipéptidos cortos habitualmente no son capaces de adoptar la estructura de hélice alfa, ya que el coste entrópico asociado con el plegamiento de la cadena polipeptídica es demasiado alto.
Importancia
Las hélices α además de ser el tipo de estructura secundaria más frecuente en las proteínas, son de gran importancia en los motivos estructurales de unión al ADN, como los motivos hélice-giro-hélice y los dedos de zinc. Esto se debe que el diámetro de 12Å de la hélice α coincide con la anchura de la hendidura mayor del ADN en forma B o B-DNA.
Otros tipos de hélice
Existen otros tipos de estructuras helicoidales similares a la hélice α en las proteínas, pero mucho menos comunes:
- Hélice : similar a la hélice α pero con 3 aminoácidos por vuelta (más extendida y más estrecha).

- Hélice π: está más comprimida y es más ancha que la hélice α (4,4 residuos por vuelta).
La hélice alfa en el arte

Julian Voss-Andreae es un escultor alemán con grados académicos en física experimental y escultura. Desde el año 2001 Voss-Andreae crea "esculturas de proteínas"[2] inspiradas en la estructura proteica, siendo la hélice α uno de sus objetos preferidos. Este artista ha fabricado esculturas de hélice α a partir de diversos materiales, como bambú y otros árboles. En el año 2004 realizó un monumento en memoria de Linus Pauling, descubridor de la hélice alfa, diseñado a partir de una gran viga de acero reordenada según la forma de la estructura de la hélice alfa. La escultura de color rojo brillante y 3 metros de altura se ubica frente a la casa de infancia de Pauling en Portland, Oregón.[3]
Véase también
Note
- -Biología. Curtis H., Barnes S., Schnek A. y Massarini A. (2008) 7ª Edición. Editorial Médica Panamericana. 34-45 pp.
- Voss-Andreae, J (2005). «Protein Sculptures: Life's Building Blocks Inspire Art». Leonardo 38: 41-45. doi:10.1162/leon.2005.38.1.41.
- Moran L, Horton RA, Scrimgeour G, Perry M (2011). Principles of Biochemistry. Boston, MA: Pearson. p. 127. ISBN 0-321-70733-8.
Referencias
- Jesús M. Sanz Estructura de macromoléculas. https://web.archive.org/web/20110724170043/http://adan-embl.ibmc.umh.es/jmsanz/estructuras.htm
- David Eisenberg, "The discovery of the α-helix and β-sheet, the principal structural features of proteins". Proceedings of the National Academy of Sciences USA. (2003). 100:11207-11210. http://www.pnas.org/cgi/content/full/100/20/11207
| Control de autoridades |
|
|---|
 Datos: Q283399
Datos: Q283399 Multimedia: Alpha helix / Q283399
Multimedia: Alpha helix / Q283399