Pyrotherium
Pyrotherium (gr. "bestia de fuego") es un género extinto de mamíferos placentarios del orden Pyrotheria, posiblemente pertenecientes al clado extinto de los meridiungulados, propios de Sudamérica.[1]
| Pyrotherium | ||
|---|---|---|
| Rango temporal: 28,4 Ma - 23 Ma Rupeliense, Oligoceno | ||
 Recreación del cráneo de Pyrotherium en el Museo Paleontológico Egidio Feruglio, en Trelew. | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Mammalia | |
| (sin rango): | Meridiungulata † | |
| Orden: | Pyrotheria † | |
| Familia: | Pyrotheriidae † | |
| Género: |
Pyrotherium † Ameghino, 1888 | |
| Especies | ||
| ||
| Sinonimia | ||
Para P. romeroi:
| ||
Hasta el momento, se han descrito dos especies válidas, Pyrotherium romeroi, la cual vivió en lo que es actualmente Argentina y P. macfaddeni de Bolivia, a finales del Oligoceno. P. romeroi en particular es el piroterio más reciente conocido del registro fósil y el mejor conocido por sus restos fósiles, que si bien incompletos son los mejor preservados en el orden entero, indicando que también son los de mayor tamaño, con una longitud corporal estimada de 2.9 a 3.6 metros.[2] También se supone que desarrolló una pequeña trompa, pero no está relacionado con los actuales elefantes (proboscídeos); tan grande es el parecido que al estudiar los restos fósiles, al piroterio se le atribuyó en el pasado un parentesco con los elefantes, si bien el verdadero parentesco de este herbívoro es aún hoy materia de controversia.[2]
Historia del descubrimiento
Los restos de originales Pyrotherium, algunos molares, un premolar y un incisivo, fueron identificados originalmente en la provincia de Neuquén en estratos que datan de la época del Oligoceno tardío, identificados por el naturalista argentino Florentino Ameghino como couche à Pyrotherium (capas de Pyrotherium, en francés) debido a la presencia de fósiles de este animal que fueron los primeros en ser identificados allí;[3][4] ahora se les conoce como parte de la edad-mamífero Deseadense por la zona de la ría Deseado, si bien se ha extendido la duda sobre si realmente el holotipo de Pyrotherium romeroi proviene de Neuquén, siendo posible que en realidad los restos procedieran de Chubut.[5] Ameghino consideraba que esas zonas correspondían a terrenos más antiguos, del Paleoceno[6] e incluso del Cretácico debido a que fueron enviados junto con restos de dinosaurios;[7] los estudios posteriores han mostrado que en realidad proceden del Oligoceno, y de hecho los fósiles de Pyrotherium han llegado a convertirse en el fósil guía de los terrenos del Oligoceno tardío. Debido a que los restos de este animal aparecieron originalmente en los lechos de ceniza volcánica de la Formación Deseado, dieron pie para el nombre del género, que significa "bestia del fuego".[8]
El nombre de la especie P. romeroi se debe al capitán del ejército argentino Antonio Romero quien envió a Ameghino los primeros restos conocidos del animal, aunque en varios textos se usan las grafía erróneas P. romeri o P. romerii.[6] Ameghino nombró a varias especies procedentes de la zona de Deseado tales como P. sorondoi con base en restos parciales, principalmente dientes, pero estudios posteriores indicaron que son parte de una sola especie.[8][9] El primer cráneo relativamente completo no apareció hasta el siglo XX, siendo descubierto por Frederic B. Loomis durante la expedición del Amherst College en 1911-1912, y catalogado como el espécimen ACM 3207.[10]
Restos adicionales del género han aparecido en Quebrada Fiera, provincia de Mendoza (Argentina) y en Salla, en el departamento de La Paz en Bolivia; estos últimos consisten en restos de una mandíbula parcial, fragmentos de huesos del cráneo, dientes y algunos hueso de las extremidades tales como piezas del húmero y astrágalos, los cuales fueron hallados entre las décadas de 1960 y 1980 e inicialmente se consideraron como parte de la especie P. romeroi,[11] y posteriormente fueron clasificados como una especie diferente y de menor tamaño, P. macfaddeni, cuyo nombre de especie es en honor del paleontólogo Bruce J. MacFadden.[8] Restos de molares y huesos del postcráneo hallados en terrenos del Oligoceno tardío de Taubaté, Brasil fueron considerados como un posible hallazgo de Pyrotherium,[12] pero es posible que correspondan a algún género diferente con el que está muy relacionado, pero aún sin describir.[13]
Descripción
Cráneo

El cráneo de Pyrotherium romeroi era alargado y estrecho, compuesto de huesos macizos. Alcanza los 72 centímetros de largo desde sus dientes frontales hasta su cóndilo occipital, y posee un hocico alargado, relativamente estrecho visto desde arriba, con narinas retraídas, una apertura nasal grande situada entre las órbitas de los ojos en medio de los hueso frontales, en paralelo al dorso del cráneo, con paredes óseas gruesas para sujeción muscular; por dentro tienen cavidades llenas de aire. La región occipital, en particular los cóndilos, era particularmente alta, como consecuencia de la flexión de la parte posterior del cráneo con respecto al plano de la base, que formaba un ángulo obtuso con el del hueso palatino; En esta y otras características, Pyrotherium se parecía al de los proboscídeos. Una pequeña cresta emerge del premaxilar y alcanza el hueso nasal, la cual parece estar rota y rodeada por una textura rugosa, que podría ser resultado de la erosión. Se desconoce qué tan grande pudo haber sido, ya que pudo ser apenas una prominencia similar a la que se observa en el proceso narial de los notoungulados y en los roedores, o incluso casi una cresta; esta cresta no es conocida en otros mamíferos, pero tal vez servía como punto de sujeción para los músculos de una posible probóscide o trompa. La cavidad para el cerebro (neurocráneo) está dañada y estaba rodeada de tejido óseo esponjoso; Loomis consideró que indicaba que en vida P. romeroi tenía un cerebro pequeño, de unos 150 milímetros de largo y 50 milímetros de ancho.[10] Análisis posteriores realizados por Bryan Patterson en 1977, tras algunos trabajos de preparación adicional del único cráneo conocido, indicaron algunos errores en las interpretaciones anteriores, y que el cerebro sería algo mayor, de 80 milímetros de ancho, más similar en tamaño al de notoungulados tales como Homalodotherium y Nesodon.[9]
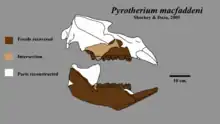
Otro rasgo muy distintivo es la presencia de dos pares de grandes incisivos orientados hacia al frente, en forma de defensas y dispuestos en un ángulo de 45°. Estos presentaban un crecimiento continuo y estaban equipados con una banda de esmalte solo en el frente. Carecía de caninos, y cuenta además con premolares y molares peculiares, con dos crestas altas transversales (bilofodontos), cuyo aspecto general recuerda a los molares de los tapires. Entre los incisivos y los dientes posteriores se extendía un espacio sin dientes, el diastema, que alcanza los 46 milímetros de largo. Los dientes en general, y particularmente los posteriores además ocupaban bastante del área del cráneo, particularmente del paladar. La región auditiva se sitúa mucho más arriba que el paladar en vista lateral y se curva hacia arriba en su parte posterior.[14] En P. macfaddeni el premaxilar tiene un par adicional de alvéolos muy pequeños, lo que sugiere que pudo haber tenido un tercer par de incisivos apenas desarrollados, y sus molares se distinguen por tener un valle bien definidos que separa los lofos anteriores y posteriores.[8]
La fórmula dental de P. romeroi es (2I/0C/3PM/3M, 1i/0c/2pm/3m).[14]

La mandíbula era robusta y tenía una sínfisis bastante desarrollada, larga y estrecha que se extendía hasta el segundo molar, un marcado foramen posterior al tercer molar y una fosa masetérica grande. Solo posee solo dos incisivos, los cuales sobresalen hacia adelante y se orientan al igual que los incisivos superiores en un ángulo de 45°, haciendo contacto con las puntas de estos; se ha pensado que estos podrían ser los segundos incisivos (i2), pero su identificación real es incierta. Al menos en P. macfaddeni poseen una capa de esmalte que solo cubre la parte ventral de los incisivos.[8] Como en el maxilar, posee premolares y molares bilofodontos; la estructura de los molares recuerda la que se encuentra en otros grandes mamíferos arcaicos, como los dinocerados, Barytherium y los deinotéridos.[14]
Postcráneo
Se han recuperado algunos huesos postcraneales de Pyrotherium romeroi, principalmente de las extremidades. La columna vertebral se conoce muy escasamente; los restos hallados incluyen sobre todo vértebras cervicales, incluyendo el atlas, el axis y las vértebras tercera y cuarta, siendo todas muy cortas. Adicionalmente se conoce una vértebra lumbar, la cual es maciza y con una apófisis reducida, un poco similar a la de Astrapotherium. Un fragmento del omóplato indica que era corto y fuerte; la cavidad glenoidea tenía el doble de su longitud y el acromion era muy alto.[10]
_BHL21733685.jpg.webp)
El húmero es relativamente corto, de 497 a 500 milímetros de longitud, pero sumamente ancho, con amplias inserciones para los músculos; se conocen también el cúbito y el radio, ambos aún más cortos, de unos 225 milímetros, y el cúbito además tenía un gran olécranon.[10] Igualmente se han identificado dos huesos de la muñeca, el unciforme derecho y el magno izquierdo, ambos siendo elementos cortos pero gruesos y de apariencia trapezoidal.[15] También se han hallado un piramidal y un semilunado. Además se incluye un astrágalo y calcáneo, y un fémur.[10]
La pelvis estaba equipada con un hueso ilíaco vasto y masivo, con una acetábulo ubicado hacia abajo y no lateralmente. El fémur carecía del tercer trocánter, con una cabeza recta mucho más alta que el trocánter mayor, y estaba aplanado anteroposteriormente; en esta especie alcanzaba los 630 milímetros de longitud, siendo mayor que el único otro fémur conocido para los piroterios, el de Baguatherium que llegaba a 558 milímetros.[16] La forma de la articulación distal permitía que la tibia se moviera hacia atrás ampliamente, lo que compensaba la falta de flexibilidad en la articulación del pie. La tibia era mucho más corta que el fémur y el peroné estaba muy cerca de la tibia, excepto en la parte central. El astrágalo estaba fuertemente aplanado, era de apariencia muy simple y sin cuello, con una tróclea tibial apenas insinuada y una faceta del navicular ubicada directamente debajo de la tróclea. El tarso de Pyrotherium era característico: el tubérculo del calcáneo estaba comprimido dorsoventralmente, así como la tróclea del astrágalo; además, presenta una reducción extrema del contacto entre el talón y el cuboide.[10] Estas características derivadas, que implican un tipo de locomoción graviportal pero plantígrada, no se encuentran en ningún otro mamífero conocido, con la excepción significativa del Arsinoitherium africano.[8]
Clasificación
Debido a que Pyrotherium posee los característicos dientes posteriores bilofodontos (es decir, con dos crestas), defensas formadas por sus incisivos superiores e inferiores, un cuerpo enorme y robusto junto a la posible presencia de una trompa, se propuso en el pasado que era un pariente cercano de los proboscídeos, o inclusive un miembro de ese grupo (Ameghino 1895, 1897; Lydekker 1896;[17] Loomis 1914).[10] Sin embargo, la mezcla de características del animal es tal que ha llevado a compararlo y relacionarlo en diferentes momentos con otros grupos, como los marsupiales diprotodóntidos (Lydekker 1893;[18] Loomis 1921[19]), los pantodontes amblípodos (Zittel 1893),[20] los perisodáctilos (Ameghino, 1888),[21] los notoungulados (Osborn 1910; Loomis 1914;[10] Scott 1913; Patterson 1977),[9] los xenungulados (Simpson 1945; Cifelli 1983; Lucas 1986, 1993), y los dinocerados a través de su supuesta relación con los xenungulados (Lucas 1986, 1993);[22] en algunos estudios, no se observaron las características más derivadas de Pyrotherium más que en otros mamíferos examinados exceptuando al embritópodo Arsinoitherium del Paleógeno de África.[8] Si esto se debe a un ancestro común, o al inusual modo de locomoción usado por estos animales (graviportal y plantígrado) continúa siendo una incógnita. Sin embargo, el propio Gaudry (1909) estableció que Pyrotherium era lo bastante diferente de cualquier otro grupo de grandes mamíferos que debía tener su propio orden, sin relación clara con los demás mamíferos.[23] El análisis más reciente realizado, el de Billet en 2010, sugiere que los piroterios como Pyrotherium son un grupo de notoungulados especializados, relacionados con Notostylops,[14][24] aunque no deja de ser una idea controvertida.[2]
Cladograma basado en el análisis filogenético de Cerdeño et al., 2017, destacando la ubicación de las especies de Pyrotherium:[15]
| Notoungulata |
| |||||||||||||||||||||||||||||||||||||||
Paleobiología

Los dientes molariformes bilofodontes de Pyrotherium fueron examinados para determinar su tipo de esmalte, por medio de un microscopio electrónico para examinar sus prismas. Los exámenes mostraron que el esmalte de Pyrotherium sigue un extraño patrón en forma de ojo de cerradura, o patrón de Boyde, en el que los prismas se encuentran densamente agrupados sin matriz interprismática entre ellos. Este tipo de prisma en el esmalte es característico de los piroterios y no es conocido en los otros órdenes de ungulados nativos suramericanos (xenungulados, astrapoterios, litopternos y notoungulados). En P. romeroi además el esmalte posee un tipo distintivo, denominado justamente "esmalte de Pyrotherium" en el cual las bandas de esmalte se disponen de forma vertical con los prismas de forma decusada (es decir, formando patrones en X).[25]
Este análisis permitió además inferir los patrones de masticación de Pyrotherium. Esta sería dominada por la llamada fase 1, en la cual la mandíbula se inclina y se dirige mesialmente, mientras las crestas cortantes de los molares iban comprimiendo el bolo alimenticio. A continuación se desarrollaba una fase 2, en la cual la mandíbula se movía lateralmente; este movimiento parece haber sido menos significativo. Este tipo de masticación y de molares se asemeja a lo observado en algunos otros mamíferos, como los canguros Macropus, el perisodáctilo Lophiodon, el marsupial Diprotodon y el proboscídeo Deinotherium, pero en estos animales su esmalte (y los lofos de los molares) se desgastan rápidamente al llegar a la adultez, quedando una superficie plana para moler, mientras que en Pyrotherium los lofos son mucho más resistentes y pueden apreciarse claramente incluso en individuos de edad avanzada, en los cuales los molares gastados aún tienen crestas afiladas. Una condición similar solo es vista en los embritópodos como Arsinoitherium, el cual también posee esmalte dispuesto en vertical y en Namatherium, el cual se parece mucho en este aspecto a Pyrotherium al tener esmalte y facetas del esmalte muy inclinadas.[25]

Debido a la robusta contextura del animal, muy seguramente era un cuadrúpedo graviportal, es decir un animal con un peso superior a la tonelada cuya estructura física está preparada para soportar esa gran masa, pero no para la velocidad.[26] Con un peso de 900 kilos en P. macfaddeni a más de 3,5 toneladas en P. romeroi con base en estimaciones de sus molares, y de 600-700 kg en P. macfadeni a 1.8 a 2.7 toneladas para P. romeroi con base en ecuaciones derivadas de la relación cabeza-cuerpo,[2] Pyrotherium estuvo entre los mayores mamíferos nativos de Suramérica. Sus huesos son sumamente densos, incluso más que en otros meridiungulados de gran porte como los notoungulados toxodontes y los astrapoterios, lo que implica una especialización extrema hacia la graviportalidad; los análisis de la densidad ósea de su húmero y fémur realizados mediante microtomografía con rayos X indican que su área medular era particularmente compacta, casi comparable a la paquiostosis de mamíferos acuáticos o semiacuáticos, con trabéculos gruesos y cavidades intratrabeculares muy pequeñas, aunque se asemejan externamente a los proboscídeos o los rinocerontes, lo cual lo ayudaría a absorber mejor la energía de impacto en los huesos.[27] Se infiere además que su postura habría sido semiplantígrada, ya que los dedos de las manos soportarían su peso, pero en cambio los pies habrían sido plantígrados, como se infiere de los huesos del tobillo.[15]
Paleoecología
Los fósiles de Pyrotherium recuperados tanto en Salla como en Deseado y Quebrada Fiera corresponden a ambientes relativamente secos, con vegetación xerofítica y con períodos de sequías;[28] esto contradiría la hipótesis de que eran animales semiacuáticos, semejantes a hipopótamos, mientras que los restos de astrapoterios (otro grupo de grandes ungulados nativos con defensas) sí se encuentran en zonas asociadas a cuerpos de agua, lo que implicaría que estos vivieron en ambientes húmedos y pudieron pasar algo de tiempo en el agua.[27] Los piroterios habrían usado sus incisivos y trompa para recolectar comida como hojas y ramas de los árboles en manera similar a los rinocerontes negros y los elefantes africanos de bosque.[28]
Pyrotherium cohabitó junto a varios otros mamíferos, varios de ellos de gran tamaño que son típicos de la fauna deseadense de sitios como La Flecha en Argentina. Destaca la presencia de esparasodontes depredadores como Pharsophorus, Notogale y el enorme Proborhyaena, y de otros ungulados principalmente del grupo de los notoungulados, tales como Trachytherus, Leontinia, Rhynchippus, Propachyrucos, Argyrohyrax, Archaeohyrax y Prohegetotherium.[29]
Referencias
- Kenneth, Rose (2006). The Beginning of the Age of Mammals (en inglés). Baltimore, Maryland: The Johns Hopkins University Press. ISBN 0-8018-8472
|isbn=incorrecto (ayuda). - Croft, D. A., Gelfo, J. N., & López, G. M. (2020). Splendid Innovation: The Extinct South American Native Ungulates. Annual Review of Earth and Planetary Sciences, 48.
- Ameghino, F. 1894. Première contribution à la conaissance de la faune mammalogique des couches à Pyrotherium. Boletín del Instituto Geográfico Argentino 15: 603–660.
- Ameghino C. 1914. Le Pyrotheríum, I'étage Pyrothéréen et les couches á Notostylops. Une response á Mr. Loomis. Physis, 1: 446-460.
- Kramarz, A. G., Forasiepi, A. M., & Bond, M. (2011). Vertebrados cenozoicos. In Relatorio del XVIII Congreso Geológico Argentino. Geología y Recursos Naturales de la Provincia del Neuquén (pp. 557-572). Buenos Aires: Asociación Geológica Argentina.
- Ameghino, F. 1889. Contribución al conocimiento de los mamíferos fósiles de la República Argentina, obra escrita bajo los auspicios de la Academia Nacional de Ciencias de la República Argentina para presentarla a la Exposición Universal de París de 1889. Actas Academia de Ciencias de Córdoba 6:1–1027.
- F. Ameghino. 1897. Mammiféres crétacés de l’Argentine (Deuxième contribution à la connaissance de la fauna mammalogique de couches à Pyrotherium) [Cretaceous mammals of Argentina (second contribution to the knowledge of the mammalian fauna of the Pyrotherium Beds)]. Boletín Instituto Geográfico Argentino 18(4–9):406-521
- Shockey, B.J., and Anaya, F. (2004). «Pyrotherium macfaddeni, sp. nov. (late Oligocene, Bolivia) and the pedal morphology of pyrotheres». Journal of Vertebrate Paleontology 24 (2): 481-488. doi:10.1671/2521.
- Patterson, B. 1977. A primitive pyrothere (Mammalia, Notoungulata) from the Early Tertiary of Northwestern Venezuela. Fieldiana, Geology 33: 397–422.
- Loomis, F.B. 1914. The Deseado Formation of Patagonia. Amherst College, Amherst, 232659 p.
- MacFadden BJ, Frailey CD (1984) Pyrotherium, a large enigmatic ungulate (Mammalia, incertae sedis) from the Deseadan (Oligocene) of Salla, Bolivia. Palaeontol 27:867–874
- Couto-Ribeiro, G. and Alvarenga, H. 2009. Primeiro registro de dentes de Pyrotherium para a Formação Tremembé, Bacia de Taubaté, SP. Reunião Anual da Sociedade Brasileira de Paleontologia. Núcleo São Paulo. Abstracts: 21.
- Ribeiro, G. D. C. Osteologia de Taubatherium paulacoutoi Soria & Alvarenga, 1989 (Notoungulata, Leontiniidae) e de um novo Pyrotheria: dois mamíferos fósseis da Formação Tremembé, Brasil (SALMA Deseadense-Oligoceno Superior). (Doctoral dissertation, Universidade de São Paulo).
- Billet, G. 2010. New observations on the skull of Pyrotherium (Pyrotheria, Mammalia) and new phylogenetic hypotheses on South American ungulates. Journal of Mammalian Evolution 17: 21–59.
- Cerdeño, E., & Vera, B. (2017). New Anatomical Data on Pyrotherium (Pyrotheriidae) from the Late Oligocene of Mendoza, Argentina. Ameghiniana, 54(3), 290-306.
- Salas, R., Sánchez, J. and Chacaltana, C. 2006. A new pre-Deseadan pyrothere (Mammalia) from Northern Peru and the wear facets of molariform teeth of Pyrotheria. Journal of Vertebrate Paleontology 26: 760–769.
- Lydekker, R. 1896. A Geographical History of Mammals. Cambridge University Press, Cambridge, 400 pp.
- Lydekker, R. 1894. Contribuciones al conocimiento de los vertebrados fósiles de Argentina. 1. Observaciones adicionales sobre los ungulados argentinos. Anales del Museo de La Plata 2: 1–91
- Loomis, F. B. 1921. Origin of South American faunas. Bulletin of the Geological Society of America. 32:187–196.
- Zittel, K. A. 1893. Handbuch der Palaeontologie. IV. Band: Vertebrata (Mammalia). Druck und verlan von R. Oldenbourg, Munchen und Leipzig.
- Ameghino, F. 1888. Rápidas diagnosis de algunos mamíferos fósiles nuevos de la República Argentina. P.E. Coni, Ed., Buenos Aires, 17 p.
- Spencer, L. (1986). Pyrothere systematics and a Caribbean route for land-mammal dispersal during the Paleocene. Revista Geológica de América Central.
- Gaudry, A. 1909. Fossiles de Patagonie: le Pyrotherium. Annales de Paléontologie 4:1–28.
- Billet, G. 2011. Phylogeny of the Notoungulata (Mammalia) based on cranial and dental characters. Journal of Systematic Palaeontology 9: 481–497.
- Koenigswald, W 647 . von, Martin, T. and Billet, G. 2015. Enamel microstructure and mastication in Pyrotherium romeroi (Pyrotheria, Mammalia). Paläontologische Zeitschrift 89: 611–634.
- Johnson, S. C. (1984). Astrapotheres from the Miocene of Colombia, South America. University of California, Berkeley.
- Houssaye, A., Fernández, V., and Billet, G. 2016. Hyperspecialization in some South American endemic ungulates revealed by long bone microstructure. Journal of Mammalian Evolution 23: 221–235.
- Croft, D. A. (2016). Horned armadillos and rafting monkeys: the fascinating fossil mammals of South America. Indiana University Press.
- Marani, H. A. (2005). Los Rhynchippinae de Edad Mamífero Deseadense de la Localidad Cabeza Blanca. Doctoral dissertation, Universidad Nacional de la Patagonia San Juan Bosco, Chubut-Argentina.
Bibliografía adicional
- F. Ameghino. 1894. Sur les oiseaux fossiles de Patagonie; et la faune mammalogique des couches à Pyrotherium. Boletín del Instituto Geographico Argentino 15:501-660
- F. Ameghino. 1901. Notices préliminaires sur des ongulés nouveaux des terrains crétacés de Patagonie [Preliminary notes on new ungulates from the Cretaceous terrains of Patagonia]. Boletín de la Academia Nacional de Ciencias de Córdoba 16:349-429
| Control de autoridades |
|
|---|
 Datos: Q2606712
Datos: Q2606712 Multimedia: Pyrotherium / Q2606712
Multimedia: Pyrotherium / Q2606712 Especies: Pyrotherium
Especies: Pyrotherium