Respiración celular
La respiración celular o respiración interna es un conjunto de reacciones bioquímicas en las cuales determinados compuestos orgánicos son degradados completamente, por oxidación, hasta convertirse en sustancias inorgánicas, proceso que proporciona energía aprovechable para la célula (principalmente en forma de ATP).[1]
Tipos de respiración celular
- Respiración aeróbica: El aceptor final de electrones es el oxígeno molecular, que se reduce a agua. La realizan la inmensa mayoría de organismos, incluidos los humanos. Los organismos que llevan a cabo este tipo de respiración reciben el nombre de organismos aeróbicos.
- Respiración anaeróbica: El aceptor final de electrones es una molécula inorgánica distinta del oxígeno.
Respiración celular

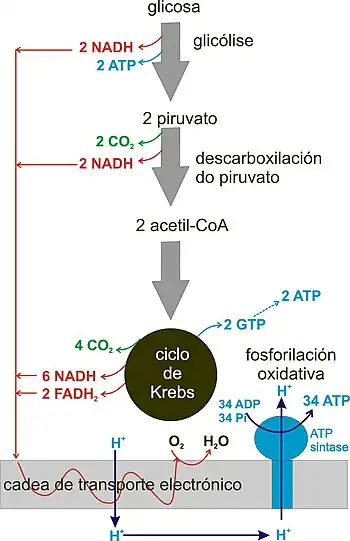
La respiración celular es un proceso metabólico que consiste en oxidar nutrientes y obtener energía de ellos en forma de ATP, para lo cual se requiere oxígeno. Los carbohidratos, lípidos y proteínas pueden ser procesados y consumidos como fuente de energía, los cuales terminan siendo completamente oxidados y reducidos a dióxido de carbono y agua, que son los productos finales de la respiración.[2][3] En algunos casos raros (en bacterias), las sustancias inorgánicas pueden oxidarse en la respiración aeróbica. En eucariotas, la respiración celular aeróbica tiene lugar en las mitocondrias, aunque la fase glucolítica previa es citosólica. En las bacterias tiene lugar en su citosol y membrana.
Los electrones que pierden los sustratos durante su degradación oxidativa respiratoria son recogidos por coenzimas, que los transferirán a la cadena de transporte de electrones y como resultado se producirá ATP durante la fosforilación oxidativa.[4] También se producen pequeñas cantidades de ATP en las primeras etapas (glucólisis) y en el ciclo de Krebs por fosforilación a nivel de sustrato. En general, los electrones que perdieron los sustratos durante su oxidación terminan siendo recogidos por el oxígeno al final de la cadena de transporte de electrones, ya que funciona como el aceptor final de electrones.[4]
Los grupos de dos carbonos en forma de acetil-CoA deben entrar, en la respiración aeróbica en el ciclo de Krebs.[2] Esta acetil-CoA puede tener varios orígenes. Se puede obtener a partir de la glucosa, que sufre un proceso en el citosol llamado glucólisis, que la transforma en piruvato, que luego da lugar a la acetil-CoA.[5] Para "respirar" (oxidar) la glucosa siempre debe existir esta fase glucolítica previa. También se puede obtener a partir de ácidos grasos, mediante el proceso catabólico mitocondrial de beta-oxidación de ácidos grasos, que da lugar a grandes cantidades de acetil-CoA.[6] Finalmente, se puede obtener con menor frecuencia a partir del catabolismo de aminoácidos.[7]
La reacción respiratoria general de una molécula de glucosa (C6H12O6) es la siguiente:
Que ΔG sea negativo indica que se libera energía, que es la que se utiliza para formar ATP a partir de ADP. En general, se estima que la producción de ATP por molécula de glucosa es de 36 a 38 (2 en la glucólisis, 2 en el ciclo de Krebs y el resto en la fosforilación oxidativa) [3]. Pero esta es una cifra máxima teórica, ya que en condiciones de células reales el rendimiento es menor, y se estima en 29 a 30 ATP por glucosa.
El metabolismo aeróbico es 19 veces más eficiente que el anaeróbico fermentativo (que solo produce 2 moles de ATP por mol de glucosa). En la oxidación de la glucosa los procesos respiratorios aeróbicos y las fermentaciones anaeróbicas comparten la fase inicial de la glucólisis. Las fases posglucolíticas son diferentes.
La ecuación de la respiración celular de glucosa que se muestra arriba es una ecuación global, ya que en realidad la respiración es un proceso muy complejo, que consta de varias fases: glucólisis (fase anterior), descarboxilación del piruvato, ciclo de Krebs, transporte de electrones en la cadena de transporte de electrones y fosforilación oxidativa.
Características
Se produce en la mitocondria. La respiración celular, como componente del metabolismo, es un proceso catabólico, en el cual la energía contenida en los sustratos usados como combustible es liberada de manera controlada. Durante la misma, buena parte de la energía libre desprendida en estas reacciones exotérmicas es incorporada a la molécula de ATP (o de nucleótidos trifosfato equivalentes), que puede ser a continuación utilizada en los procesos endotérmicos, como son los de mantenimiento y desarrollo celular (anabolismo).
Los substratos habitualmente usados en la respiración celular son la glucosa, otros hidratos de carbono, ácidos grasos,[8] incluso aminoácidos, cuerpos cetónicos u otros compuestos orgánicos. En los animales estos combustibles pueden provenir del alimento, de los que se extraen durante la digestión, o de las reservas corporales. En las plantas su origen puede ser asimismo las reservas, pero también la glucosa obtenida durante la fotosíntesis.
La mayor parte del ATP producido en la respiración celular se produce en tres etapas: glucólisis, ciclo de Krebs y cadena de transporte de electrones.
Glucólisis
Tiene lugar en el citosol de la mayoría de las células vivas, incluyendo muchas bacterias anaerobias.[9] Puede ocurrir en condiciones anaeróbicas (sin oxígeno) y es similar a la fermentación. Consta de 10 reacciones enzimáticas en las que la glucosa (molécula de 6 carbonos) se convierte en dos moléculas de piruvato (molécula de 3 carbonos), por lo que en la glucólisis no hay pérdida de carbono en forma de CO2.[9] El piruvato es una molécula más oxidada que la glucosa. Durante estas reacciones se producen dos fosforilaciones a nivel de sustrato, gracias a las cuales se generan 2 ATP netos, que es el único ATP que se produce directamente en la glucólisis.[9] También se producen dos moléculas de la coenzima reducida NADH,[9] que luego pueden ser llevadas a la mitocondria por sistemas de transporte llamados lanzaderas (lanzadera del malato-aspartato y lanzadera del glicerol-fosfato) y allí pueden transferir sus electrones a la cadena de transporte de electrones y generar más ATP (hasta 3 ATP por NADH, un total de 6 ATP). También se forman dos moléculas de agua. Luego, en condiciones aeróbicas, el piruvato será llevado a la mitocondria para continuar con la siguiente fase de la respiración.[5][10]
La ecuación global de la glucólisis es la siguiente:
Descarboxilación oxidativa del piruvato
En los eucariotas aeróbicos, el piruvato ingresa a la mitocondria a través de un sistema de transportadores de membrana. Una vez allí, la enzima piruvato deshidrogenasa cataliza su descarboxilación y unión del resto de la molécula a la coenzima A. La reacción produce una molécula de CO2, que procede del grupo carboxilo del piruvato, y es el primer carbono de la glucosa que se pierde. En los humanos saldrá de la célula, pasará al torrente sanguíneo y será expulsado por los pulmones durante la respiración pulmonar (en otros seres saldrá por las branquias, tráquea, estomas vegetales...). Los dos carbonos restantes del piruvato se unen a la coenzima A, formando acetil-CoA, que luego liberará esos dos carbonos al ciclo de Krebs en la siguiente fase. También se produce una molécula de la coenzima reducida NADH, que va a la cadena de transporte de electrones, donde dará lugar a la formación de 3 ATP por molécula. Recuerda que cada glucosa dio lugar a dos piruvatos, por lo que todos los productos de esta fase en realidad se duplican si los contamos como glucosa. La descarboxilación del piruvato es, por tanto, una reacción de conexión entre la glucólisis y el ciclo de Krebs. la reacción es:[11]
Ciclo de Krebs
También llamado ciclo del ácido cítrico o ciclo de los ácidos tricarboxílicos o, es un ciclo de reacciones enzimáticas que tiene lugar en la matriz mitocondrial (y el citosol bacteriano) en el que el grupo de dos carbonos proporcionado por el acetil-CoA se condensa con oxalacetato para formar ácido cítrico, que luego sufre varias reacciones en ciclo en las que finalmente se regenera el oxalacetato.[12] El acetil-CoA que entra en el ciclo proviene del piruvato de la glucólisis o del catabolismo de lípidos y aminoácidos.[13] Durante el ciclo del ácido cítrico, se liberan dos moléculas de CO2 por cada acetil-CoA, por lo que finalmente se eliminan todos los carbonos entrantes (entre la descarboxilación del piruvato y el ciclo de Krebs, se pierden los 6 carbonos de la glucosa inicial).[14] En el ciclo hay una fosforilación a nivel de sustrato en la que cada acetil-CoA produce una molécula de GTP (que luego origina ATP enzimáticamente).[14] También hay cuatro deshidrogenaciones en las que se forman 3 moléculas de NADH y una de FADH2 (esta última unida a la enzima como grupo prostético).[14] Las coenzimas transferirán sus electrones a la cadena de transporte de electrones, que servirá para producir alrededor de 11 ATP. Sumando el ATP producido a partir de GTP serían 12 ATP por cada acetil-CoA. Si lo contamos por molécula de glucosa, el rendimiento en el ciclo de Krebs sería de 24 ATP y, contando la descarboxilación de piruvato y glucólisis, sería de hasta 38 ATP totales.[2][15]
Cadena de transporte de electrones y fosforilación oxidativa
La cadena de transporte de electrones o cadena respiratoria o se encuentra en la membrana mitocondrial interna o membrana bacteriana.[16] Consiste en un conjunto de transportadores que se pueden pasar electrones entre sí en reacciones redox. La cadena recibe estos electrones de las coenzimas formadas durante la respiración.[16] Estas coenzimas, en el caso de la respiración de glucosa, se forman en: el ciclo de Krebs, la descarboxilación del piruvato, y en la fase previa de la glucólisis. Otras deshidrogenaciones biológicas y la beta-oxidación de ácidos grasos también pueden producir coenzimas reducidas. Los electrones que producen las coenzimas circulan a través de la cadena respiratoria liberando gradualmente energía, que se utiliza para bombear protones al espacio intermembrana mitocondrial (o espacio periplásmico de la superficie bacteriana).[16] Al final de la cadena está el oxígeno, que recoge electrones y se une a los protones para formar agua (½ O2 + 2e- + 2H + → H2O). Necesitamos respirar oxígeno para que realice esta función en la cadena respiratoria mitocondrial. Los protones bombeados en la cadena se acumulan en el espacio intermembrana creando un gradiente, ya que la membrana es impermeable a ellos y una vez bombeados no pueden regresar.[4][17]
En la membrana mitocondrial interna (y en la membrana bacteriana) también hay una compleja nanomáquina llamada ATP sintasa que puede sintetizar ATP a partir de ADP y fosfato. Esta máquina, formada por más de 23 subunidades proteicas, tiene un canal para dejar pasar los protones acumulados. Cuando este flujo de protones pasa a través de la ATP sintasa, esta se activa y comienza a sintetizar ATP. Este proceso se denomina fosforilación oxidativa y produce la mayor parte del ATP de la célula. Por tanto, el funcionamiento de la cadena de transporte electrónico se combina con la fosforilación oxidativa.[18]
En definitiva, en la respiración los electrones que se arrancan de los sustratos durante su oxidación pasan a las coenzimas, para luego circular por la cadena respiratoria y llegar al oxígeno. A medida que circulan crean un gradiente de protones, esto activa la producción de ATP en la ATP sintasa.[18] Además, las coenzimas se reciclan (reoxidan) cediendo electrones en la cadena y están listas para su reutilización.[4]
Balance energético
De hecho, estrictamente hablando, la respiración sería a partir del ciclo de Krebs, ya que las primeras etapas dependen del tipo de sustancia que se “respira”. Si oxidamos los ácidos grasos por beta-oxidación, por ejemplo, la glucólisis y la descarboxilación del piruvato ya no se producirían de antemano, sino que serían necesarias en la oxidación de una glucosa. En la siguiente tabla podemos ver las fases y el balance energético total de la respiración completa de una molécula de glucosa hasta CO2 y H2O. Se indica la máxima producción teórica de ATP.
| Fase | Producción de coenzimas | Producción de ATP | Modo de producción do ATP | |
|---|---|---|---|---|
| Glucólisis | 2 netos | Fosforilación a nivel de substrato (se producen 4 pero se gastan 2, por lo que el balance neto es de 2) | ||
| 2 NADH | 4-6 | Fosforilación oxidativa - Cada NADH citosólico produce de 2 a 3 ATP dependiendo del sistema de lanzadera usado para el transporte a la mitocondria (lanzadera del glicerol-fosfato o lanzadera del malato-aspartato) | ||
| Descarboxilación oxidativa del piruvato | 2 NADH | 6 | Fosforilación oxidativa | |
| Ciclo de Krebs | 2 | Fosforilación a nivel de substrato | ||
| 6 NADH | 18 | Fosforilación oxidativa | ||
| 2 FADH2 | 4 | Fosforilación oxidativa | ||
| Rendimiento total | 36-38 ATP | Obtenidas de la oxidación completa de una molécula de glucosa y la oxidación posterior de todos los coenzimas reducidos formados | ||
Aunque el rendimiento teórico es de hasta 38 ATP por glucosa, generalmente es menor debido a la pérdida de electrones en la cadena respiratoria y los costos del transporte activo de moléculas a las mitocondrias. En general, se estima que la producción neta real no es más de 28 a 30 ATP por glucosa, pero se desconoce la cifra exacta.
Desacoplamiento de la respiración mitocondrial y síntesis de ATP
En algunas células, la respiración mitocondrial y la síntesis de ATP puede desacoplarse por la proteína desacopladora, con lo que la mayor parte de la energía producida no se transforma en ATP sino que se disipa en forma de calor. Este proceso ocurre en la denominada grasa parda y se pone en marcha el despertar de la hibernación de ciertos animales o para proteger del frío al recién nacido.[19][20]
Fermentación
Sin oxígeno, el piruvato (ácido pirúvico) no es metabolizado por la respiración celular sino que sufre un proceso de fermentación. El piruvato no se transporta a la mitocondria sino que permanece en el citoplasma, donde se convierte en productos de desecho que pueden eliminarse de la célula. Esto tiene el propósito de oxidar los transportadores de electrones para que puedan realizar la glucólisis nuevamente y eliminar el exceso de piruvato. La fermentación oxida NADH a NAD+ para que pueda reutilizarse en la glucólisis. En ausencia de oxígeno, la fermentación evita la acumulación de NADH en el citoplasma y proporciona NAD+ para la glucólisis. Este producto de desecho varía según el organismo. En los músculos esqueléticos, el producto de desecho es el ácido láctico. Este tipo de fermentación se llama fermentación láctica. En el ejercicio extenuante, cuando las demandas de energía superan el suministro de energía, la cadena respiratoria no puede procesar todos los átomos de hidrógeno unidos por NADH. Durante la glucólisis anaeróbica, el NAD+ se regenera cuando los pares de hidrógeno se combinan con el piruvato para formar lactato. La formación de lactato es catalizada por la lactato deshidrogenasa en una reacción reversible. El lactato también se puede utilizar como precursor indirecto del glucógeno hepático. Durante la recuperación, cuando el oxígeno está disponible, el NAD+ se une al hidrógeno del lactato para formar ATP. En levaduras, los productos de desecho son etanol y dióxido de carbono. Este tipo de fermentación se conoce como fermentación alcohólica o etanólica. El ATP generado en este proceso se produce por fosforilación a nivel de sustrato, que no requiere oxígeno.[21]
La fermentación es menos eficiente en el uso de la energía de la glucosa: solo se producen 2 ATP por glucosa, en comparación con los 38 ATP por glucosa producidos nominalmente por la respiración aeróbica. Esto se debe a que la mayor parte de la energía de la respiración aeróbica se deriva del O2 con su doble enlace relativamente débil y de alta energía. El ATP glicolítico, sin embargo, se crea más rápidamente. Para que los procariotas continúen con una tasa de crecimiento rápida cuando pasan de un ambiente aeróbico a un ambiente anaeróbico, deben aumentar la tasa de las reacciones glucolíticas. Para los organismos multicelulares, durante períodos breves de actividad extenuante, las células musculares usan la fermentación para complementar la producción de ATP de la respiración aeróbica más lenta, por lo que una célula puede usar la fermentación incluso antes de que se agoten los niveles de oxígeno, como es el caso en los deportes que no requiere que los atletas controlen su propio ritmo, como en las carreras de velocidad.
Diferencias entre fermentación y respiración aerobia
La fermentación es muy diferente de la respiración, siendo las principales diferencias:
- No utiliza un aceptor de electrones externo (el papel que juega el oxígeno en la respiración aeróbica). En las reacciones de fermentación se suelen formar coenzimas en las reacciones previas que recogen los electrones.
- La cadena de transporte electrónico no está involucrada.
- La ATP sintasa no está involucrada y no hay fosforilación oxidativa, ya que todo el ATP se forma por fosforilación a nivel de sustrato y siempre en cantidades mucho más pequeñas.
- Los productos finales de la fermentación siguen siendo moléculas relativamente complejas y ligeramente oxidadas. La respiración, por otro lado, produce productos finales muy simples y muy oxidados (CO2 y H2O).
Respiración anaeróbica
La respiración anaeróbica es utilizada por algunos microorganismos en los que ni el oxígeno (respiración aeróbica) ni los derivados del piruvato (fermentación) son los aceptores finales de electrones de alta energía. En su lugar, se usa un aceptor inorgánico como sulfato (SO42-), nitrato (NO3-) o azufre (S).[19][22]. Los transportadores electrónicos usados por estas bacterias son muy similares a los encontrados en la cadena respiratoria mitocondrial.[19] Dichos organismos se encuentran típicamente en lugares inusuales, como cuevas submarinas o cerca de respiraderos hidrotermales en el fondo del océano.
Véase también
Referencias
- Biología III. Los códigos de la vida, pag 72, en Google libros
- Lehninger, 1988, p. 435-460.
- Alberts et al., Raff, p. 70-71.
- Lehninger, 1988, p. 467-505.
- Lehninger, 1988, p. 397-429.
- Lehninger, 1988, p. 511-527.
- Lehninger, 1988, p. 531-557.
- Iglesias, Vanesa Álvarez. Estudio multidisciplinar de la variabilidad del ADN mitocondrial en poblaciones humanas.. Univ Santiago de Compostela. Consultado el 21 de marzo de 2018.
- Alberts et al., Raff, p. 71.
- Alberts et al., Raff, p. 71-75, 102-103.
- Lehninger, 1988, p. 437-441.
- Alberts et al., Raff, p. 80.
- Alberts et al., Raff, p. 81.
- Alberts et al., Raff, p. 80-81.
- Alberts et al., Raff, p. 80-82, 104-105.
- Alberts et al., Raff, p. 82-83.
- Alberts et al., Raff, p. 82-83, 761-766.
- Alberts et al., Raff, p. 766-787.
- Alberts et al., Raff, p. 785.
- Paniagua Gómez-Álvarez, 2002, p. 230.
- Alberts et al., Raff, p. 72-73.
- Lumen Boundless Microbiology. «Anaerobic Respiration-Electron Donors and Acceptors in Anaerobic Respiration». courses.lumenlearning.org (en inglés). Boundless.com. Consultado el 19 de noviembre de 2020. «Anaerobic respiration is the formation of ATP without oxygen. This method still incorporates the respiratory electron transport chain, but without using oxygen as the terminal electron acceptor. Instead, molecules such as sulfate (SO42-), nitrate (NO3–), or sulfur (S) are used as electron acceptors ».
Bibliografía
- Lehninger, Albert L. (1988). Principios de bioquímica. Barcelona: Omega. ISBN 84-282-0738-0.
- Alberts, Bruce; Johnson, Alexander; Lewis, Julian; Morgan, David; Raff, Martin; Roberts, K.; Walter, P. (2016). Biología molecular de la célula (6ª edición). Barcelona: Omega S.A. ISBN 978-84-282-1638-8.
- Paniagua Gómez-Álvarez, Ricardo (2002). Citología e histología vegetal y animal (3ª edición). Madrid: McGraw-Hill - Interamericana de España, S.A.U. ISBN 84-481-9984-7.
Enlaces externos
| Control de autoridades |
|
|---|
 Datos: Q912751
Datos: Q912751 Multimedia: Cellular respiration / Q912751
Multimedia: Cellular respiration / Q912751