Retículo endoplasmático
El retículo endoplasmático o endoplásmico[1] es un orgánulo que se encuentra en el citoplasma de la célula eucariota, se presenta como una compleja red dispuesta en forma de túbulos, sacos aplanados y cisternas, que están interconectadas entre sí, con una organización variada en los diferentes tipos celulares.
El retículo muestra sus membranas organizadas en regiones, las que realizan diferentes funciones. Existen dos regiones extensas y permanentes que son: el retículo endoplasmático rugoso, con sus membranas formando túbulos más o menos rectos, sacos aplanados o cisternas, con numerosos ribosomas asociados, y el retículo endoplasmático liso, sin ribosomas asociados y con membranas organizadas formando túbulos muy curvados e irregulares y cisternas.[2]
El retículo endoplasmático (RE) interviene en funciones relacionadas con la síntesis proteica, el metabolismo de lípidos y algunos esteroides, así como el transporte intracelular, el metabolismo de carbohidratos, el almacenamiento de calcio y la metabolización de toxinas.
El retículo es multifuncional y necesita gran coordinación con el medio intracelular y una respuesta rápida a los cambios (estrés) en el medio intrarreticular. Frente al estrés, el RE mantiene su homeostasis mediante la activación de las vías de respuesta metabólicas. La activación prolongada de la vías de respuesta al estrés en el RE, es habitual en la diabetes, la enfermedad inflamatoria intestinal y varios cánceres.[3]

del Núcleo
Rugoso
Liso

Características generales
El retículo endoplasmático (RE), es un complejo sistema de membranas, que establece dos compartimentos uno externo (citoplasmático) y otro interno intraluminal (intratubular o intracisternal).[4]
El RE es característico de la célula eucariota con núcleo verdadero, ya sean organismos unicelulares o bien organismos multicelulares.
La cantidad de retículo endoplasmático que se encuentra presente en una célula varia dependiendo del estado metabólico de la misma.[5]
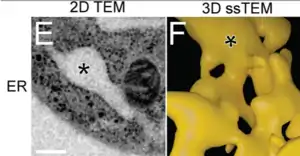
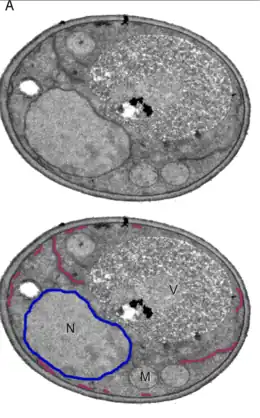

El retículo endoplásmico (RE), forma una extensa red de cisternas y túbulos.
Es una red dinámica en constante cambio y adaptación a las necesidades funcionales de la célula.
El RE se ha mostrado como un centro de red y forma conexiones físicas con casi todos los orgánulos unidos a su membrana.
El concepto de retículo endoplásmico como compartimientos aislados, propia de libros de texto del siglo XX, ha cambiado. Se entiende ahora al RE como estructuras que se comunican con otros orgánulos, permitiendo la transferencia de lípidos e iones, la señalización y la segmentación de orgánulos mediante sus sitios de contacto de membrana (Membrane contact site (MCS) en inglés).
El RE regula una amplia variedad de funciones clave, mediante gran cantidad de procesos celulares a través de sus MCS con varios orgánulos y con la membrana plasmática. Los sitios de contacto del retículo endoplásmico regulan la dinámica de otros orgánulos.
[6]
[7][8]
Historia
Mediante la microscopía óptica se podía identificar dentro de las células secretoras, teñidas con hematoxilina o con azul de toluidina, un 'componente basófilo' o cromófilo en el sector basal infranuclear.
Una estructura o componente de aspecto filamentoso estriado, en el sector basal de las células secretoras, había sido notada por varios investigadores, entre ellos: Pflüger en 1875, Heidenhain en 1875, y Ebertih et Muller en 1892.
En 1894 el investigador alemán Solger, describió una estructura basal de aspecto filamentoso, en las células secretoras de las glándulas salivales. Por extensión, la denominación de 'filamentos de Solger' se aplicó en todas las células secretoras.[9][10][11][12]
Entre 1897 y 1900 Garnier denominó a ese componente basófilo como 'ergastoplasma' (del griego ergastés: trabajo, función) por su actividad funcional, asociada a la síntesis. También describió que el ergastoplasma, visto a través de su microscopio, mostraba un aspecto fibrilar y que era observable en el polo basal de las células en la etapa presecretora.[13][14][15][10]
<<...les filaments... participent d'une façon active au processus de la sécrétion, qu'ils sont en quelque sorte un véritable organe dans la cellule secrétrice.>> ...los filamentos... participan de una manera activa en los procesos de secreción, siendo, de alguna manera, un verdadero órgano dentro de la célula secretora. —Charles Garnier, 1897.[10] |
En 1951 Bernhard planteó la estructura fibrilar del Ergastoplasma en los hepatocitos con el microscopio electrónico.[16]
En 1953 Porter aclaró mediante microscopía electrónica, que el componente conocido como 'ergastoplasma', que mostraba con la citoquímica un alto contenido de ARN y por tanto basófilo, era una red compleja de hebras, que llamó retículo endoplásmico.[17]
En 1954 Palade y Porter, sobre células de aves cultivadas in vitro, establecían que el 'retículo endoplásmico' era una red de cavidades. Que estas podían agrandarse hasta convertirse en vesículas aplanadas relativamente vastas, que denominaron 'cisternas'.
En 1955 Palade planteaba que la membrana que limitaba las cavidades del retículo endoplásmico, parecía ser continua con la membrana celular y con las membranas nucleares.[18]
[19]
Estructura
Microarquitectura del retículo
Con el microscopio óptico los investigadores identifican dentro de las células secretoras, teñidas con hematoxilina o con azul de toluidina, un 'componente basófilo' o cromófilo en el sector basal.
Esta estructura basófila, observada con aumentos medianos, presenta un aspecto fibrilar grueso y corresponde al retículo endoplasmático rugoso asociado con ribosomas y ácido ribonucleico.[10][20]
Son necesarios grandes aumentos para poder observar los túbulos, sacos y cisternas del retículo, en el corte teñido de una célula.
El retículo endoplasmático rugoso y el retículo liso, suelen ocupar espacios celulares diferentes en los hepatocitos, en las neuronas y en las células que sintetizan esteroides.
Ultraestructura del retículo

El carácter universal de estas estructuras en los eucariotes, fue reconocido finalmente con el desarrollo del microscopio electrónico (ME).
El ME permite observar un abundante sistema de membranas, que limita cavidades aplanadas y a menudo superpuestas también llamadas láminas o planchas (sheet en inglés), o que forman tubos contorneados en el interior del hialoplasma.
Este conjunto polimórfico de cavidades que se encuentran en toda la célula, se denominó por su aspecto de red como retículo endoplásmico.
Puede representar por sí solo de 50-60 % de la superficies de las membranas celulares totales, más o menos dilatadas y limitadas por una membrana de 5 a 6 nanómetros (nm) de espesor.[21]
La disposición espacial del retículo endoplasmático en las células animales depende de sus interacciones con los microtúbulos, mientras que en las vegetales los responsables son los filamentos de actina.[2]
Dominios del Retículo endoplasmático
El retículo endoplasmático tiene zonas que se diferencian por su estructura y funciones, no se distribuyen de forma homogénea en todo el RE, sino que se limitan a distintas subregiones que se denominan dominios, e incluyen: la envoltura nuclear (EN), el retículo Rugoso (RER), el retículo Liso (REL) y las regiones que entran en contacto con otros orgánulos.
Además se describen Nanodominios funcionales a lo largo de los túbulos, formados por alternancias y agrupamientos de moléculas.[22][23][24]

Retículo endoplasmático rugoso.
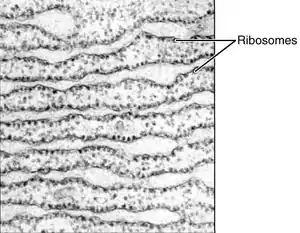
El retículo endoplasmático rugoso (RER), recibe este nombre debido a los numerosos ribosomas, adheridos del lado del citosol de su membrana (hemimembrana citosólica), mediante sus subunidades mayores con coeficiente de sedimentación de 60 S.
El RER tiene unos sáculos interconectados de aspecto redondeado, cuyo interior se conoce como luz del retículo o lumen, donde caen las proteínas sintetizadas por el ribosoma. Está muy desarrollado en las células que por su función, deben realizar una activa labor de síntesis, como las células hepáticas o las células del páncreas.[25][26]
Retículo endoplasmático liso.
El retículo endoplásmico liso (REL) no presenta ribosomas en su estructura y entre sus funciones principales está: participar en el metabolismo de lípidos, el almacenamiento de calcio y la desintoxicación de drogas. Debido a esta última función es muy abundante en hepatocitos que aumenta con la ingesta de sustancias tóxicas como el alcohol.
En las células musculares lisas y estriadas se encontraron una forma especializada de retículo endoplásmico liso conocida como retículo sarcoplásmico el cual es un importante almacén del calcio que se utiliza en el proceso de contracción muscular.
En el retículo endoplasmático liso también se sintetizan lípidos como los triacilgliceroles que serán almacenados en el propio retículo o en gotas lipídicas citosólicas. Este proceso es muy activo en los adipocitos, células que almacenan grasa, con dos funciones: reserva alimenticia y aislamiento térmico. También es el principal responsable de la síntesis de la parte lipídica de las lipoproteínas, de la producción de hormonas esteroideas y de ácidos biliares.[2]
Funciones
- Biosíntesis proteica: El ARN mensajero proviene de la transcripción del ADN nuclear y es su imagen especular. Al llegar al retículo endoplasmático, se fija a unas estructuras específicas llamadas ribosomas, adheridas al retículo endoplasmático, gracias a las riboforinas. Allí participa en la síntesis de proteínas, determinando el orden en que se unirán los aminoácidos. La información está codificada en forma de tripletes: cada tres bases constituyen un codón que determina un aminoácido. Las reglas de correspondencia entre codones y aminoácidos constituyen el código genético. Los aminoácidos son enviados por el ARN de transferencia, específico para cada uno de ellos, y son trasportados hasta el ARN mensajero, donde se aparean el codón de este y el anticodón del ARN de transferencia, por complementar de bases, y de esta forma se sitúan en la posición que les corresponde. Una vez finalizada la síntesis de una proteína, el ARN mensajero queda libre y puede ser leído de nuevo. De hecho, es muy frecuente que antes de que finalice una proteína ya está comenzando otra, con lo cual, una misma molécula de ARN mensajero, está siendo utilizada por varios ribosomas simultáneamente.
- Metabolismo de lípidos: Dado que no tiene ribosomas, en el retículo endoplasmático liso no se sintetizan proteínas. Pero tiene un papel esencial en la síntesis de lípidos de la membrana plasmática, colesterol y derivados de este, como los ácidos biliares o las hormonas esteroideas.
- Desintoxicación: Es un proceso que se lleva a cabo principalmente en las células del hígado y que consiste en la inactivación de productos tóxicos como drogas, medicamentos o los propios productos del metabolismo celular, por ser liposolubles (hepatocitos).
- Glicosilación: Son reacciones de transferencia de un oligosacárido a las proteínas sintetizadas. Se realiza en la membrana del retículo endoplasmático. De este modo, la proteína sintetizada se transforma en una proteína periférica externa del glucocálix en la reproducción de lisosomas.
- Desfosforilación de la glucosa-6 fosfato.- La glucosa generalmente se guarda como glucógeno, básicamente en el hígado. Este órgano es el principal encargado de administrar glucosa a la sangre, debido al control realizado por las hormonas glucagón e insulina. El desbasamiento del glucógeno produce glucosa-6-fosfato que no puede cruzar las películas y de esta manera no puede salir de las células. La glucosa 6-fosfatasa se encarga de disponer de ese depósito de fosfato, permitiendo que la glucosa sea transportada a la célula exterior.[2]
- Reservorio intracelular de calcio.- Las cisternas del retículo endoplásmico liso también están especializadas en el secuestro y almacenamiento de calcio del citosol, gracias a las bombas de calcio ubicadas en sus membranas. La concentración de ion Ca2+ dentro del retículo está en el orden milimolar (mM, 1 × 10-3), mientras que en el citosol su concentración es nanomolar (nM), 1 × 10-9). Este calcio puede emerger masivamente en respuesta a señales extra o intracelulares a través de cascadas de segundo mensajero, y desencadenar respuestas celulares tales como exocitosis. Otro ejemplo notable es el retículo sarcoplásmico (un nombre que recibe el retículo endoplasmático liso en las células musculares) que secuestra el calcio a través de una bomba de calcio en sus membranas. La secuenciación y la salida de calcio del retículo sarcoplásmico se produce en cada ciclo de contracción de la célula muscular.[2]
- Acumulación de productos.- El retículo almacena una gran cantidad de sustancias en el espacio cisternal o lumen.
Como por ejemplo:
-Las proteínas que sintetizan sustancias que se modificarán posteriormente. Las mismas que se almacenan temporalmente en el retículo hasta que cumplan con su objetivo final.
-Existen otro tipo de proteínas que son residentes del retículo es decir nunca lo dejan. Estas proteínas tienen una secuencia de aminoácidos que las unen al retículo. Por lo que a pesar de que escapen del retículo, volverán a ser receptadas.
Función del retículo endoplasmático en la síntesis de proteínas
La mayoría de las células son capaces de secretar proteínas de alto peso molecular hacia el exterior. En los animales las proteínas sintetizadas aseguran funciones muy diversas.
Varios compartimientos intracelulares especializados están implicados en los procesos de secreción de proteínas, y el primero de ellos es el retículo endoplasmático.[21]
Un estudio científico de 2013 muestra hallazgos que respaldan un modelo en el que una función secretora no canónica está coordinada por la función COPI en células de mamíferos bajo el agotamiento de Sar1 (A+B), como una alternativa al modelo vesicular clásico dependiente de COPII. Por lo tanto, a partir de nuestros datos, es evidente que pueden surgir compartimentos de exportación no canónicos a expensas de los clásicos, para respaldar el transporte de una serie de proteínas que se sintetizan en el retículo endoplasmático sin pasar por la función de COPII.[27]
Biogénesis del retículo endoplasmático.
El retículo endoplásmico liso y el retículo rugoso pierden membrana de manera constante, debido a la formación de vacuolas de secreción, por lo que necesitan regenerarse constantemente.
Las proteínas de la membrana del retículo endoplásmico rugoso, son fabricadas por los ribosomas adheridos a ella.[28]
- Las enzimas translocadoras organizan la membrana.
- Las proteínas se sintetizan en el citoplasma y en el lumen.[2]
Véase también
Referencias
- OMS,OPS (ed.). «Retículo endoplásmico». Descriptores en Ciencias de la Salud. Biblioteca virtual de salud.
- Megías M.; Molist P.; Pombal M.A. (2020). «5: Tráfico vesicular. Retículo endoplasmático, Ampliado». Atlas de histología vegetal y animal. Galicia: Departamento de Biología Funcional y Ciencias de la Salud, Facultad de Biología, Universidad de Vigo.
- Dianne S. Schwarz; Michael D. Blower (2016). «The endoplasmic reticulum: structure, function and response to cellular signaling.». Cellular and Molecular Life Sciences 73: 79-94. Consultado el 12 de julio de 2021.
- Cooper, Geoffrey (2000). «The Endoplasmic Reticulum». The Cell: A Molecular Approach. Sinauer Associates, Inc. Consultado el mes de julio de 2021.
- Welsch U. (2009). «2: La célula». Histología de Sobotta. Médica Panamericana. pp. 45-46. Consultado el 13 de julio de 2021.
- Wu H.; Carvalho P.; Voeltz G.K. (2018). «Here, there, and everywhere: The importance of ER membrane contact sites». Science (Revisión) (en inglés) 361 (6401): eaan5835. Consultado el 14 de julio de 2021.
- Lee J.E.; Cathey P.I.; Wu H.; Parker R.; Voeltz G.K. (2020). «Endoplasmic reticulum contact sites regulate the dynamics of membraneless Organelles». Science (en inglés) 367 (6477): eaay7108. doi:10.1126/science.aay7108. Consultado el 14 de julio de 2021.
- Scorrano, L.; De Matteis, M.A.; Emr, S.; Giordano F.; Hajnóczky G.; Kornmann B.; Lackner L.L.; Levine T.P.; Pellegrini L.; Reinisch K.; Rizzuto R.; Simmen T.; Stenmark H.; Ungermann C.; Schuldiner M. (2019). «Coming together to define membrane contact sites.». Nat Commun 10 (1287). Consultado el 17 de julio de 2021.

- Santiago Ramón y Cajal (1897). «XII:Tejidos compuesto. Tejido glandular.». Elementos de histología normal y de técnica micrográfica para uso de estudiantes. Nicolás Moya. p. 454.
- Georges Grignon; Monique Grignon (1989). «Charles Garnier et la découverte de l'ergastoplasme.» (PDF). Société Française d'Histoire de la Médecine. (en francés) (Faculté B de médecine de Nancy): 205-209.
- Crespo Ferrer P.V. (12 de marzo de 2010). Biología y Terapia Celular. Mitos y Realidades. Real Academia de Medicina y Cirugía de Granada. p. 13.
- Georges Grignon. «A Nancy, une découverte prophetique, un nom oublié: Charles Garnier (1875-1958)» (en francés).
- Garnier, Charles (1897). «Les filaments basaux des cellules glandulaires. Note préliminaire.». Bibliographie anatomique. 5: 278-289.
- Garnier, Charles (1900). «Contribution à l’étude de la structure et du fonctionnement des cellules glandulaires (du rôle de l’ergastoplasme dans la sécrétion).». J. anat. et physiol. (Paris) 36 (22). Weiss J.M., The Ergastoplasm.
- Weiss J.M. (1953). «The Ergastoplasm.». J Exp Med (PDF) (en inglés) 98 (6): 607-618. PMID 13109112. doi:10.1084/jem.98.6.607. Consultado el 27 de junio de 2021.

- Bernhard W.; Gautier A.; Oberling C. (1951). «Fibrillary Elements of Probable Ergastoplasmic Nature in Cytoplasm of Hepatic Cells Revealed by Electron Microscopy». Comptes rendus des seances de la Societe de biologie et de ses filiales 145 (7-8): 566-569.
- Porter K.R. (1953). «Observations on a submicroscopic basophilic component of Cytoplasm». J Exp Med 97 (5): 727-750. PMID 13052830. doi:10.1084/jem.97.5.727. Consultado el 27 de junio de 2021.
- Palade G.E.; Porter K.R. (1954). «Studies on the Endoplasmic Reticulum: I. Its Identification in cells in situ». J Exp Med. 100 (6): 641-656. PMID 13211920. doi:10.1084/jem.100.6.641. Consultado el 27 de junio de 2021.
- Palade G.E. (1955). «Studies on the endoplasmic reticulum. II. Simple dispositions in cells in situ». J Biophys Biochem Cytol (PubMed) 1 (6): 567-582. PMID 13278367. doi:10.1083/jcb.1.6.567. Consultado el 27 de junio de 2021.
- Castellanos N.; Martínez L.C.; Silva E.H.; Teodoro A.V.; Serrão J.E.; Oliveira E.E. (2017). «Ultrastructural analysis of salivary glands in a phytophagous stink bug revealed the presence of unexpected muscles.». PLoS ONE 12 (6): e0179478. doi:10.1371/journal.pone.0179478.
- Callen, Jean Claude. «capítulo 9». Biología celular de las moléculas a los organismos. ISBN 2-10-003197-X.
- Voeltz G.K.; Rolls M.M.; Rapoport T.A. (2002). «Structural organization of the endoplasmic reticulum». EMBO Reports (Revisión) 3: 944-950. Consultado el 17 de julio de 2021.
- Baumann O.; Walz B. (2001). «Endoplasmic reticulum of animal cells and its organization into structural and functional domains.». Int Rev Cytol (Revisión) 205: 149-214. doi:10.1016/s0074-7696(01)05004-5. (requiere suscripción).
- Gao G.; Zhu C.; Liu E.; Nabi I.R. (2019). «Reticulon and CLIMP-63 regulate nanodomain organization of peripheral ER tubules.». PLoS Biology 17 (8): e3000355. doi:10.1371/journal.pbio.3000355.
- «Tema 11: Ribosomas.». Organización general de las células: Citosol y sistema de endomembranas. Departamento de Histología, Universidad Miguel Hernández.
- Salvador Resino (2013). «Retículo endoplásmico». Epidemiología Molecular de Enfermedades Infecciosas (EMEI). Consultado el 27 de junio de 2021.
- Cutrona M., Beznoussenko G.V., Fusella A., Martella O., Moral P., Mironov A. (2013). «Silencing of Mammalian Sar1 Isoforms Reveals COPII-Independent Protein Sorting and Transport». Traffic 14 (6): 691-708. doi:10.1111/tra.12060. Consultado el 12 de mayo de 2020.
- La gaveta (29 de mayo de 2013). «Biogénesis de las estructuras de la célula eucariota animal». http://gavetasdemiescritorio.blogspot.com/2013/05/biogenesis-de-las-estructuras-de-la.html. Consultado el 24 de febrero de 2017.
Bibliografía
- Alberts, Bruce (2004). Biología Molecular de la Célula 4ªEdición. Barcelona: Ediciones Omega. ¡¡j77577589+69.
Enlaces externos.
 Wikimedia Commons alberga una categoría multimedia sobre Retículo endoplasmático.
Wikimedia Commons alberga una categoría multimedia sobre Retículo endoplasmático.
 Datos: Q79927
Datos: Q79927- Multimedia: Endoplasmic reticulum / Q79927