Sarahsaurus aurifontanalis
Sarahsaurus aurifontanalis es la única especie conocida del género extinto Sarahsaurus de dinosaurio saurópodo prosaurópodo que vivió a principios del período Jurásico, hace aproximadamente entre 200 y 188 millones de años, entre el Sinemuriano al Pliensbachiano, en lo que hoy es Norteamérica. Sus restos aparecen en lo que hoy es el noreste de Arizona, EE. UU., en la Formación Kayenta cerca de Gold Spring. Fue nombrado por primera vez por Timothy B. Rowe, Hans-Dieter Sues y Robert R. Reisz en 2010 y la especie tipo es llamada Sarahsaurus aurifontanalis. Sarahsaurus es el tercer dinosaurio sauropodomorfo basal que se ha identificado en América del Norte. Se cree que llegaron a través de un evento de dispersión que se originó en América del Sur, distinto del de los otros dos sauropodomorfos.Sarahsaurus se conoce por el holotipo, un esqueleto casi completo articulado denominado TMM 43646-2, otro esqueleto parcial denominado TMM 43646-3, y un cráneo casi completo, pero en mal estado denominado MCZ 8893.[1]
| Sarahsaurus aurifontanalis | ||
|---|---|---|
| Rango temporal: 200 Ma - 188 Ma Jurásico inferior | ||
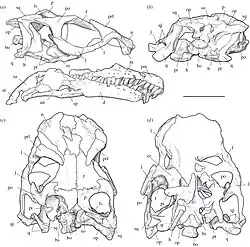 Ilustración del cráneo en vistas múltiples | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Sauropodomorpha | |
| Género: |
Sarahsaurus Rowe, Sues & Reisz, 2010 | |
| Especie: |
S. aurifontanalis Rowe, Sues & Reisz, 2010 | |
Descripción
Sarahsaurus era un miembro basal, es decir "primitivo", del linaje sauropodomorfo, que también incluyó los más derivados, o "avanzados", saurópodos gigantes de cuatro patas herbívoros con cuellos largos y colas. Los saurópodos basales como Sarahsaurus ya se parecen a los saurópodos en varios aspectos, incluido el cuello alargado y la estructura robusta, pero recuerdan a los dinosaurios ancestrales en otros. Sarahsaurus era bípedo, de dos patas, con extremidades anteriores mucho más cortas que las traseras. extremidades y equipado con poderosas manos agarradoras.[2][3] Sarahsaurus era de tamaño medio para un sauropodomorfo basal, con una columna vertebral que medía más de 3,1 metros de longitud.[3] El paleontólogo Gregory Paul , en un libro de divulgación de 2016, estimó su longitud en 4 metros y su peso corporal en 200 kilogramos.[4]

Cráneo
La naris externa, abertura del cráneo para la fosa nasal, tenía menos de la mitad del tamaño de la órbita, cuenca del ojo. Estaba bordeado por la nariz, el premaxilar y en pequeña medida, el maxilar, los dos últimos formando la mandíbula superior. Entre el premaxilar y el maxilar había una gran abertura, el agujero subnarial. La fosa antorbital, una abertura adicional del cráneo que se ve en la mayoría de los dinosaurios y que estaba situada entre la nariz externa y la órbita, tenía menos de la mitad de la longitud de la órbita. Gran parte del margen superior de la órbita estaba formado por el hueso frontal del techo del cráneo. Este hueso, sin embargo, no contribuyó al margen frontal de la fenestra temporal superior en el extremo posterior del techo del cráneo. En cambio, este margen frontal estaba formado por el los huesos postorbitarios y parietales, con la parte frontal del parietal bifurcada para conectarse con este último. Entre el parietal y el supraoccipital, que sigue detrás del parietal en la parte posterior del cráneo, había otra abertura, la fenestra posparietal, que era grande en Sarahsaurus. Una característica única del género se puede encontrar en la parte inferior de la caja del cráneo, donde una cresta poco profunda se extendía entre los procesos basipterigoideos, proyecciones óseas hacia abajo de la caja del cráneo que apuntalan a este último contra el paladar. La mandíbula inferior se curvó hacia abajo hacia su punta, y la articulación de la mandíbula estaba ubicada muy por debajo del nivel de la fila de dientes. No hay evidencia de pico.[1][3]
Cada lado de la mandíbula superior estaba revestido con 20 dientes, cuatro en la premaxilar, el hueso frontal de la mandíbula superior y 16 en el maxilar que seguía detrás. El hueso dentario de la mandíbula inferior también tenía 20 dientes. Los dientes eran moderadamente heterodontos, es decir, la forma de los dientes variaba dentro de la mandíbula y se sentaban ligeramente oblicuamente en la mandíbula, superponiéndose ligeramente entre sí. Sus bordes cortantes fueron dentados con hasta 20 dentículos por diente.[1]
Postcráneo
La columna estaba compuesta por 10 vértebras cervicales, 14 vértebras dorsales, tres vértebras sacras y aproximadamente 50 vértebras caudales. A diferencia de los saurópodos posteriores, los lados de las vértebras dorsales no mostraban cavidades profundas causadas por sacos de aire que invaden el hueso, neumática esquelética poscraneal. La escápula tenía forma de reloj de arena. En la muestra de holotipo, la escápula y la coracoides no estaban fusionadas, probablemente conectadas sólo a través del cartílago, esto podría indicar que el individuo aún no estaba completamente maduro. La muestra de holotipo incluye el esternón y la clavícula, que rara vez se encuentran con esqueletos de dinosaurios. La clavícula es un hueso en forma de varilla que se articula con el extremo inferior de la escápula en una articulación inmóvil. Ha sido controvertido si las clavículas izquierda y derecha de los sauropodomorfos estaban unidas entre sí en sus extremos, conectando así las cinturas escapular izquierda y derecha, o si estaban presionadas contra sus escápulas a lo largo de su longitud. La orientación de la clavícula en Sarahsaurus confirma que lo primero fue el caso. Los extremos de ambas clavículas probablemente habrían estado conectados entre sí por un ligamento, aunque no se fusionaron. En vista lateral, la escápula se inclinó entre 45 ° y 65 ° con respecto a la horizontal. Esto habría permitido que los coracoideos izquierdo y derecho casi se tocaran entre sí en la línea media del cuerpo, mientras que los esternones izquierdo y derecho, que estaban unidos entre sí en la línea media, se habrían ubicado directamente detrás de los coracoides.[1][3]
El húmero, hueso de la parte superior del brazo, tenía más de la mitad de la longitud del fémur, hueso de la parte superior del muslo y tenía una gran cresta en la parte frontal para la inserción de los músculos, la cresta deltopectoral , que se extendía por la mitad de la longitud del hueso y tenía un borde en forma de S. En la mano, el ungual, hueso de la garra del dedo índice era el más grande.[1]
En la cadera, el hueso ilíaco tenía dos proyecciones óseas orientadas hacia abajo, el pedúnculo isquiático y púbico, que conectaban con los dos huesos inferiores de la cadera, el isquion y el pubis, respectivamente, como en otros dinosaurios. En Sarahsaurus, el pedúnculo isquiático tenía solo la mitad de la longitud del pedúnculo púbico. El acetábulo, articulación de la cadera, estaba formado por los tres huesos de la cadera, como en otros dinosaurios, aunque en Sarahsaurus el isquion contribuyó con menos de la mitad que el pubis. El pubis era único en tener dos aberturas separadas, el foramen obturador y el foramen púbico, este último faltaba en la mayoría de los demás arcosaurios. El fémur era recto y elíptico en sección transversal. La tibia, espinilla, alcanzó el 84% de la longitud del fémur.[1]
Descubrimiento e investigación
Todos los especímenes de Sarahsaurus fueron recolectados de la Formación Kayenta del Jurásico Inferior cerca de Gold Spring, Arizona. El género se basa en un esqueleto casi completo y articulado con un cráneo fragmentario y desarticulado, holotipo , número de muestra TMM 43646-2. Además, se asignó al género un esqueleto parcial, número de muestra TMM 43646-3 y un cráneo casi completo, número de muestra MCZ 8893.[1] Este último espécimen se denominó originalmente Massospondylus sp..[5] El cráneo completo esta aplastado y dividido horizontalmente, separando el techo del cráneo del paladar, esta división fue causada por la hinchazón y la contracción periódicas de la arcilla circundante después del entierro. Mientras que el individuo holotipo estaba maduro, el segundo cráneo indica un individuo menos maduro como lo indican las líneas de sutura abiertas que separan los huesos del cráneo de la caja del cráneo.[1][3]
Sarahsaurus fue descrito por primera vez por Timothy B. Rowe, Hans-Dieter Sues y Robert R. Reisz en 2011, la especie tipo es Sarahsaurus aurifontanalis. El nombre genérico honra a Sarah Butler, una filántropa de Austin. Ayudó a financiar la exhibición "Dino Pit" en el Austin Nature and Science Center, que Rowe ayudó a crear. Este fue citado diciendo: "Le dije a Sarah que si realmente recaudaba un millón de dólares para construir el Dino Pit, le pondría su nombre a un dinosaurio"."[6] El nombre específico se deriva de aurum del latín, "oro" y fontanalis también del Latín, "de la primavera" en referencia a Gold Spring, Arizona, donde se encontró el holotipo. Sarahsaurus es el cuarto dinosaurio sauropodomorfo basal que se ha identificado en América del Norte. Los otros tres son Anchisaurus y Ammosaurus del Jurásico Temprano del Valle del Río Connecticut y Seitaad del posterior Navajo Sandstone del Jurásico Temprano de Utah. Se cree que apareció a través de un evento de dispersión que se originó en América del Sur y estaba separado de los otros sauropodomorfos.[1] El animal se destaca por poseer manos muy grandes y poderosas, lo que posiblemente sugiere que era un omnívoro.[7]
Clasificación
En un análisis cladístico, presentado por Apaldetti y sus colegas en noviembre de 2011, se encontró que Sarahsaurus estaba más estrechamente relacionado con Ignavusaurus dentro de Massopoda. Se encontró que su grupo era intermedio entre los plateosáuridos y los massospondílidos, siendo más derivado que el primero y más primitivo que el segundo.[8] Un análisis cladístico de 2018 realizado por Chapelle y Choiniere recuperó a Sarahsaurus e Ignavusaurus como massospondílidos, reflejando la asignación original de Sarahsaurus a Massospondylus.[9]
Filogenia
Cladograma siguiendo lo encontrado por Apaldetti y sus colegas en noviembre de 2011.
| Plateosauria |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleoecología
Aún no se ha realizado una datación radiométrica definitiva de la Formación Kayenta, y la correlación estratigráfica disponible se ha basado en una combinación de fechas radiométricas de fósiles de vertebrados, magnetoestratigrafía y evidencia de polen.[10] Se ha conjeturado que la Formación Kayenta fue depositada durante las etapas Sinemuriense y Pliensbachiense del período Jurásico Temprano, hace aproximadamente 199 a 182 millones de años.[11] La Formación Kayenta es parte del Grupo Glen Canyon que incluye formaciones no solo en el norte de Arizona sino también en partes del sureste de Utah, el oeste de Colorado y el noroeste de Nuevo México. La formación fue depositada principalmente por ríos. Durante el Jurásico Temprano, la tierra que ahora es la Formación Kayenta experimentó veranos lluviosos e inviernos secos. En el período Jurásico medio, un campo de dunas arenosas lo invadía desde el norte que se convertiría en la arenisca de Navajo.[12] Los animales aquí se adaptaron a un clima estacional y se podía encontrar abundante agua en arroyos, estanques y lagos.
Sarahsaurus compartió su paleoambiente con otros dinosaurios, como varios terópodos, incluidos Dilophosaurus, Kayentavenator,[13] Coelophysis kayentakatae y el terópodo "Shake N Bake", y los dinosaurios blindados Scelidosaurus y Scutellosaurus. La Formación Kayenta ha producido un conjunto de organismos pequeño pero creciente.[14] Los vertebrados presentes en la Formación Kayenta en la época de Saharasaurus incluían tiburones hibodontes, peces óseos indeterminados, peces pulmonados, salamandras y la rana Prosalirus, la cecilia Eocaecilia, la tortuga Kayentachelys, un reptil esfenodontido, varios lagartos y el pterosaurio Rhamphinion. También estaban presentes los sinápsidos Dinnebitodon, Kayentatherium, Oligokyphus, morganucodóntidos, el posible mamífero primitivo Dinnetherium y un mamífero haramiyido.[15] Varios crocodilomorfos tempranos estuvieron presentes, incluidos Calsoyasuchus, Eopneumatosuchus, Kayentasuchus y Protosuchus.[14][15][16][17] Los fósiles de rastros de vertebrados de esta área incluyeron coprolitos[18] y los rastros de terápsidos, animales parecidos a lagartos y dinosaurios, lo que proporcionó evidencia de que estos animales también estaban presentes.[19] Los no vertebrados en este ecosistema incluían piedra caliza microbiana o "alga",[18] bivalvos de agua dulce, mejillones y caracoles de agua dulce[12] y ostrácodos.[20] La vida vegetal conocida en esta área incluía árboles que se conservaron como madera petrificada.[16]
Referencias
- Timothy B. Rowe, Hans-Dieter Sues and Robert R. Reisz (2010). «Dispersal and diversity in the earliest North American sauropodomorph dinosaurs, with a description of a new taxon». Proc. R. Soc. B. In press. doi:10.1098/rspb.2010.1867.
- Yates, Adam M. (2012). «Basal Sauropodomorpha: The "Prosauropods"». En M. K. Brett-Surman; James O. Farlow; Thomas R. Holtz, eds. The Complete Dinosaur (2 edición). Indiana University Press. pp. 424-443. ISBN 978-0-253-35701-4.
- Adam D. Marsh, Timothy B. Rowe (2018):Anatomy and systematics of the sauropodomorph Sarahsaurus aurifontanalis from the Early Jurassic Kayenta Formation:https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0204007
- Paul, Gregory S. (2016). The Princeton Field Guide to Dinosaurs (2nd Edition). Princeton, New Jersey: Princeton University Press. p. 189. ISBN 9780691167664.
- Attridge, J.; A.W. Crompton; Farish A. Jenkins Jr (1985). «The southern Liassic prosauropod Massospondylus discovered in North America». Journal of Vertebrate Paleontology 5 (2): 128-132. doi:10.1080/02724634.1985.10011850.
- https://cns.utexas.edu/news/sarahsaurus
- http://news.nationalgeographic.com/news/2010/10/101006-new-dinosaur-north-america-science/
- Cecilia Apaldetti, Ricardo N. Martinez, Oscar A. Alcober and Diego Pol (2011). «A New Basal Sauropodomorph (Dinosauria: Saurischia) from Quebrada del Barro Formation (Marayes-El Carrizal Basin), Northwestern Argentina». En Claessens, Leon, ed. PLoS ONE 6 (11): e26964. PMC 3212523. PMID 22096511. doi:10.1371/journal.pone.0026964.
- Chapelle KEJ, Choiniere JN. (2018) A revised cranial description of Massospondylus carinatus Owen (Dinosauria: Sauropodomorpha) based on computed tomographic scans and a review of cranial characters for basal Sauropodomorpha. PeerJ 6:e4224 https://doi.org/10.7717/peerj.4224
- J. M. Clark and D. E. Fastovsky. 1986. Vertebrate biostratigraphy of the Glen Canyon Group in northern Arizona. The Beginning of the Age of the Dinosaurs: Faunal change across the Triassic-Jurassic boundary, N. C. Fraser and H.-D. Sues (eds.), Cambridge University Press 285–301
- Padian, K (1997) Glen Canyon Group In: Encyclopedia of Dinosaurs, edited by Currie, P. J., and Padian, K., Academic Press.
- Harshbarger, J. W.; Repenning, C. A.; Irwin, J. H. (1957). Stratigraphy of the uppermost Triassic and the Jurassic rocks of the Navajo country. Professional Paper 291. Washington, D.C.: U.S. Geological Survey.
- Gay, R. 2010. Kayentavenator elysiae, a new tetanuran from the early Jurassic of Arizona. Pages 27–43 in Gay, R. Notes on early Mesozoic theropods. Lulu Press (on-demand online press).
- Lucas, S. G.; Heckert, A. B.; Tanner, L. H. (2005). «Arizona's Jurassic fossil vertebrates and the age of the Glen Canyon Group». En Heckert, A. B.; Lucas, S. G., eds. Vertebrate paleontology in Arizona. Bulletin 29. Albuquerque, NM: New Mexico Museum of Natural History and Science. pp. 95-104.
- Jenkins, F. A., Jr., Crompton, A. W., and Downs, W. R. 1983. Mesozoic mammals from Arizona: new evidence in mammalian evolution. Science 222(4629):1233–1235.
- Jenkins, F. A., Jr. and Shubin, N. H. 1998. Prosalirus bitis and the anuran caudopelvic mechanism. Journal of Vertebrate Paleontology 18(3):495–510.
- Curtis, K., and Padian, K. 1999. An Early Jurassic microvertebrate fauna from the Kayenta Formation of northeastern Arizona: microfaunal change across the Triassic-Jurassic boundary. PaleoBios 19(2):19–37.
- Luttrell, P. R., and Morales, M. 1993. Bridging the gap across Moenkopi Wash: a lithostratigraphic correlation. Aspects of Mesozoic geology and paleontology of the Colorado Plateau. Pages 111–127 in Morales, M., editor. Museum of Northern Arizona, Flagstaff, AZ. Bulletin 59.
- Hamblin, A. H., and Foster, J. R. 2000. Ancient animal footprints and traces in the Grand Staircase-Escalante National Monument, south-central Utah. Pages 557–568 in Sprinkel, D. A., Chidsey, T. C., Jr., and Anderson, P. B. editors. Geology of Utah's parks and monuments. Utah Geological Association, Salt Lake City, UT. Publication 28.
- Lucas, S. G., and Tanner L. H. 2007. Tetrapod biostratigraphy and biochronology of the Triassic-Jurassic transition on the southern Colorado Plateau, USA. Palaeogeography, Palaeoclimatology, Palaeoecology 244(1–4):242–256.
| Control de autoridades |
|
|---|
 Datos: Q3950358
Datos: Q3950358 Especies: Sarahsaurus aurifontanalis
Especies: Sarahsaurus aurifontanalis