Scipionyx samniticus
Scipionyx samniticus es la única especie conocida del género extinto Scipionyx (lat. "garra de Escipión") de dinosaurio terópodo compsognátido que vivió a mediados del período Cretácico, hace aproximadamente 113 millones de años, en el Albiense, en la actual Europa. El nombre proviene de la palabra latina Scipio, por Scipione Breislak,[1] el géologo que hiciera la descripción del lugar donde se encontró el fósil y por Escipión el Africano, vencedor de Aníbal Barca en las segunda guerra púnica, unida al término onyx del griego que significa garra, traduciéndose como “la Garra de Escipión”[1] mientras que el nombre de la especie samniticus se debe al Samnio, región donde fue encontrado. El espécimen es popularmente apodado "Skippy".[1]

| Scipionyx samniticus | ||
|---|---|---|
| Rango temporal: 113 Ma Cretácico inferior | ||
 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| Infraorden: | Coelurosauria | |
| Familia: | Compsognathidae | |
| Género: |
Scipionyx dal Sasso & Signore, 1998 | |
| Especie: |
S. samniticus dal Sasso & Signore, 1998 | |
Sólo se conoce un fósil de Scipionyx, descubierto en 1981 por un paleontólogo aficionado y llamado la atención de la ciencia en 1993. En 1998 se nombró la especie tipo Scipionyx samniticus, cuyo nombre genérico significa "garra de Scipio". El hallazgo generó mucha publicidad debido a la preservación única de grandes áreas de tejido blando petrificado y órganos internos como músculos e intestinos. El fósil muestra muchos detalles de estos, incluso la estructura interna de algunas células musculares y óseas. También fue el primer dinosaurio encontrado en Italia. Debido a la importancia del ejemplar, se ha estudiado intensamente.
El fósil es el de un juvenil de solo medio metro de largo y quizás solo de tres días de edad. Se desconoce su tamaño adulto. Scipionyx era un depredador bípedo, su grupa horizontal equilibrada por una cola larga. Su cuerpo probablemente estaba cubierto por plumas primitivas, pero estas no se han encontrado en el fósil, es decir, sin restos de piel.
En las entrañas del fósil todavía están presentes algunas comidas a medio digerir, lo que indica que Scipionyx comió lagartijas y pescado. Quizás sus padres se los habían dado al animal joven. Varios científicos han intentado aprender de la posición de los órganos internos cómo respiraba Scipionyx, pero sus conclusiones a menudo no concuerdan.
La clasificación de Scipionyx es incierta, debido a las dificultades de clasificar un taxón conocido solo de un espécimen tan joven. La mayoría de los paleontólogos lo han clasificado como miembro de Compsognathidae, una familia de pequeños celurosaurios, pero un investigador ha propuesto que puede pertenecer a Carcharodontosauridae , una familia de grandes carnosaurios.
Descripción
Sólo se ha descubierto un ejemplar juvenil, notable por la preservación del tejido blando de sus órganos internos. El ejemplar medía 23 centímetros de largo y 15 de alto, por lo que se estima que un adulto llegó a los 2 metros de largo, 0,70 de alto y a pesar 20 kilogramos.

El holotipo de Scipionyx representa un individuo muy pequeño, la longitud conservada es de solo 237 milímetros. En 2011, dal Sasso & Maganuco estimó su longitud total, incluida la sección faltante de la cola, en 461 milímetros. El espécimen no era mucho más pequeño que los embriones conocidos o las crías de Lourinhasaurus y Allosaurus, terópodos de considerable magnitud. Sin embargo, dadas sus afinidades con Compsognathidae, es probable que el tamaño adulto de Scipionyx no supere al del compsognátido más grande conocido, Sinocalliopteryx de 237 centímetros de largo. Como la cría habría encajado dentro de un huevo de unos once centímetros de largo y seis centímetros de ancho, esto habría implicado un tamaño de huevo bastante alto en comparación con la longitud corporal del adulto.[2]
Debido a que el holotipo es una cría de quizás solo unos pocos días de edad, es difícil determinar la constitución del animal adulto, pero se pueden sacar algunas conclusiones generales de manera confiable. Scipionyx era un pequeño depredador bípedo. Su rabadilla horizontal estaba equilibrada por una cola larga. El cuello era relativamente largo y delgado. Las patas traseras y especialmente las anteriores eran bastante alargadas. Dal Sasso & Maganuco consideraron probable la presencia de una capa de protoplumas primitivas, conocidas también por algunos parientes directos.[2]
El estudio de 2011 estableció ocho rasgos derivados únicos o autapomorfías en las que Scipionyx difería de sus parientes más cercanos. El premaxillar tiene cinco dientes. Donde el hueso parietal y frontal hacen contacto, la depresión en la que estaba presente la fenestra supratemporal, una abertura del techo del cráneo, muestra una cresta sinuosa en el postorbital. La rama inferior del escamosoTiene un final rectangular. La muñeca consta de solo dos huesos superpuestos, un elemento radial y otro inferior formado por una fusión del primer y segundo carpo. Este último elemento tiene forma de lente, no de media luna; está aplanado y se fusiona a la perfección. El primer dedo está notablemente alargado, un 23% más largo que el tercer dedo. La muesca en el borde frontal del ilion se dirige hacia el frente y solo se desarrolla débilmente. El borde frontal del eje del isquion tiene un largo proceso obturador con un extremo rectangular.[2]
Cráneo
El cráneo del holotipo es grande, en comparación con el tamaño del cuerpo, y cuenta con cuencas oculares muy grandes. Esto se debe en gran parte a su corta edad. En consecuencia, la fenestra antorbital semicircular, la abertura del cráneo normalmente más grande, es también corta y más pequeña que la cuenca del ojo. Frente a él hay dos aberturas más pequeñas, la maxilar y la promaxilar. El hocico es puntiagudo con una punta redondeada baja. La premaxila, el hueso que forma la parte frontal del hocico, tiene cinco dientes. El maxilar detrás de él es profundo con una rama frontal muy corta. Lleva siete dientes. La depresión en su superficie para la abertura antorbital está delimitada por una cresta. El lagrimal es robusto y carece de cuerno, su costado no está perforado por un foramen. Los prefrontales son excepcionalmente grandes y forman una gran parte del borde superior frontal de la cuenca del ojo. Los huesos frontales tienen una cresta transversal atrás. Entre los frontales y los parietales, el techo del cráneo en una distancia limitada aún no se ha cerrado, lo que resulta en una llamativa abertura en forma de diamante, una fontanela que se confundió por primera vez con el daño infligido al fósil durante la primera preparación. En su lado interno, la fenestra supratemporal no tiene depresión, estando delimitada por un borde alto del parietal. El yugal no tiene rama vertical frontal hacia el lagrimal. El hueso cuadrado tiene en su borde frontal una gran expansión en forma de ala, tocando el pterigoideo. Los huesos de la caja cerebral son en gran parte inaccesibles, pero se ve una pequeña abertura en el oído interno, el recessus tympanicus dorsalis . La parte inferior de la caja craneana carece de una parte inflada o bulla .[2]

La mandíbula inferior es recta y alargada. El hueso de la mandíbula es bastante bajo, la muestra crea la ilusión de una mandíbula fuerte porque la mandíbula izquierda es visible debajo de la derecha. Tiene diez dientes. En la descripción de 1998, una parte del esplenio se confundió con un supradentario y el angular se identificó erróneamente como surangular porque en el fósil se había desplazado hacia arriba, creando la falsa impresión de una fenestra mandibular externa, una abertura en el lado externo de la mandíbula, estaría presente.[2]
Scipionyx tiene cinco dientes en el premaxilar, siete en el maxilar y diez en el dentario de la mandíbula inferior para un total de veintidós por lado y un gran total para la cabeza de cuarenta y cuatro. El número de cinco dientes premaxilares es sorprendente, ya que un total de cuatro es normal para los compsognátidos, por lo contrario, solo algunos carnosaurios tener cinco. Debido a la corta edad de la pieza, el ciclo de reemplazo dentario aún no había comenzado, provocando una perfecta simetría dental entre los maxilares izquierdo y derecho. Los dientes carecen de la forma compsognátida típica con un vértice de la corona del diente que se repite repentinamente. En cambio, en general se curvan gradualmente, sólo los dientes más grandes muestran algo de "torcedura". Excepcionalmente, la fila de dientes del maxilar inferior se extiende más hacia atrás que la del maxilar superior. Los dientes premaxilares son puntiagudos y carecen de dentículos. Los primeros cuatro tienen una sección transversal ovalada, el quinto está más aplanado cerca de su vértice. El segundo y quinto diente son los más grandes. Los dientes superiores están aplanados con dentículos en sus bordes posteriores. El segundo y cuarto dientes maxilares son los más grandes; siendo este último el diente más grande de todos. De los diez dientes de la mandíbula inferior, los dos primeros son bastante rectos con una sección transversal ovalada y carecen de dentículos. El tercer diente tiene dentículos en su base y una parte superior más plana; los otros siete son más recurvados y aplanados en toda su altura; gradualmente los dentículos alcanzan el ápice.[2]
Postcraneo
La columna vertebral de Scipionyx probablemente incluye diez vértebras cervicales y trece vértebras dorsales. Debido al hecho de que el espécimen es solo una cría, la diferenciación entre las dos categorías no se ha desarrollado completamente, lo que hace que cualquier distinción sea bastante arbitraria. Con certeza están presentes cinco vértebras sacras. El fósil ha conservado solo nueve vértebras de la cola, probablemente cincuenta o más habían estado presentes originalmente. Las vértebras del cuello son opistocelulares. El axis está neumatizado como un pneumatoporo, una abertura a través de la cual un divertículo del saco de aire de la base del cuello podría llegar a su interior hueco, es visible de lado. La tercera, cuarta y quinta vértebras también presentan pneumatoporos pero la serie consecutiva carece de ellos, lo que es sorprendente ya que se suponía que el proceso de neumatización se habría iniciado por la parte posterior, avanzando por sí mismo. Contrariamente a lo afirmado por el estudio de 1998, las costillas cervicales son muy alargadas, con una longitud de hasta tres centros vertebrales.[2]

Las vértebras de la espalda no están neumatizadas. Son anfiplatas de sección ovalada y tienen espinas bajas de perfil hexagonal. Justo debajo de la parte superior de la columna, en el borde frontal y posterior, hay un pequeño proceso en forma de pico. En 1998, interpretado como un complejo hiposfeno-hipántro reducid , un sistema de articulaciones vertebrales secundarias mostrado por muchos terópodos, fue visto por el estudio de 2011 como un par de puntos de unión para tendones, como se identificó en 2006 en Compsognathus . Excepcionalmente, con la decimotercera vértebra las dos apófisis costales, la parapófisis y la diapófisis, se colocan al mismo nivel. Las cinco vértebras sacras aún no se han fusionado en un verdadero sacro. Las vértebras de la cola son platicolas con espinas bajas y chevrones inclinados hacia atrás.[2]
Hay al menos doce pares de costillas dorsales, algunos elementos desplazados podrían representar un decimotercer par. La tercera y la cuarta costilla tienen extremos inferiores expandidos que en vida probablemente estaban adheridos a las costillas cartilaginosas del esternón, a su vez conectados al esternón que en el espécimen holotipo no se han osificado todavía. La grupa inferior está cubierta por una canasta de dieciocho pares de gastralia o costillas del vientre. Dal Sasso & Maganuco interpretan las misteriosas partes del fuste presentes cerca de las extremidades anteriores como los restos de un decimonoveno elemento frontal que consta de dos fustes completamente fusionados homólogos a los elementos mediales normales de un par de gastralia, un hueso con forma de galón también se ha informado con Juravenator. Los gastralia forman un patrón en espiga, los elementos mediales izquierdo y derecho se superponen entre sí en sus extremos bifurcados para que la canasta pueda expandirse y contraerse para acomodar los movimientos respiratorios del abdomen.[2]
Miembro superior

La escápula es relativamente recta y de seis a siete veces más larga que ancha, falta su extremo superior. Su extremo inferior está conectado a un coracoides semicircular. La furcula es ancha y más o menos en forma de U con sus dos ramas en ángulo de 125°. La extremidad anterior es bastante larga. Su longitud es igual al 48% de la longitud del cuerpo por delante de la pelvis. Especialmente la mano se alarga como es típico de los compsognátidos, sin embargo, para un miembro de ese grupo, Scipionyx tiene una mano relativamente corta. El húmero es recto con una cresta deltopectoral moderadamente desarrollada. El cúbito es delgado y cilíndrico con una longitud del 70% de la del húmero. La muñeca consta de dos elementos únicamente, un hueso radial que cubre el extremo inferior del radio y un hueso en forma de disco debajo de él, este es el primer carpo inferior agrandado o una fusión perfecta y sin costuras del primer y segundo carpo inferior. El metacarpo es compacto y moderadamente alargado. Sus tres elementos reflejan la forma de los dedos que llevan, el primero es el más corto y el más grueso, el segundo el más largo y el tercero es intermedio en longitud y espesor. El tercer dedo es excepcionalmente largo para un comspognátido, con un 123 % de la longitud del pulgar. Como la articulación inferior del primer metacarpiano está biselado, el pulgar diverge medialmente. Su garra no es más grande que la del segundo dedo. Las garras de las manos están moderadamente curvadas.[2]
Miembro inferior
En la pelvis, el Hueso ilion es corto y plano con un perfil superior ligeramente convexo. El extremo posterior es rectangular, el borde frontal tiene un punto adjunto en forma de gancho y cerca de su parte superior una muesca circular, un rasgo que generalmente se considera una sinapomorfia de Tyrannosauroidea. El hueso púbico apunta casi verticalmente hacia abajo y, por lo tanto, es "mesopúbico" u "ortopúbico". Es relativamente corto con aproximadamente dos tercios de la longitud del fémur. Tiene un "pie" corto con forma de palo de golf. El isquion tiene tres cuartas partes de la longitud del pubis y forma un ángulo de 54°. Termina en una pequeña expansión. En la parte delantera de su eje está presente un gran proceso obturador en forma de hacha, el accesorio para elMusculus puboischiofemoralis externus , que carece de una pequeña muesca circular entre su borde inferior y el eje, aunque esta falta se asocia normalmente con la posesión de un proceso obturatorio triangular inferior.[2]
De la extremidad trasera, falta la parte inferior de la pierna. El fémur o fémur es recto y robusto. El trocánter menor es notablemente más bajo que el trocánter mayor y está separado de él por una hendidura estrecha. Tiene la forma de una expansión en forma de ala hacia el frente. Falta un trocánter accesorio o posterior; asimismo, falta un cuarto trocánter en la diáfisis posterior. La tibia tiene sólo una cresta cnemial débil, separada de su cóndilo externo por una ranura profunda y estrecha, la incisura tibial . El peroné es ancho en la parte superior pero tiene un eje delgado.[2]
Tejidos
El holotipo conserva un conjunto excepcionalmente grande de tejidos blandos para un dinosaurio fósil. Aunque se ha informado de algunos tejidos musculares, por ejemplo Santanaraptor , Pelecanimimus, cartílago, Juravenator, Aucasaurus[3] o intestino, Mirischia de otros dinosaurios, Scipionyx es único en la conservación de algunos ejemplos de la mayoría de los principales grupos de órganos internos, sangre, vasos sanguíneos, cartílagos, tejidos conectivos, tejido óseo, tejido muscular, vainas córneas, sistema respiratorio y digestivo. El tejido nervioso y la piel externa, incluidas las posibles escamas o plumas, están ausentes.[2]
Los tejidos blandos no están presentes en forma de huellas sino como petrificaciones tridimensionales, habiendo sido reemplazados por fosfato cálcico con asombroso detalle, incluso a nivel subcelular o como restos transformados de los componentes biomoleculares originales.[2]
Tejido óseo
El tejido óseo original ya no está presente, pero la mineralización de fosfato de calcio ha conservado la estructura de las células óseas originales, mostrando osteocitos individuales, incluidos sus espacios huecos internos y los canalículos. También se han conservado los vasos sanguíneos internos del hueso, en algunos casos todavía vacíos por dentro. En algunos huesos, incluidos algunos del cráneo y la mandíbula inferior, el periostio aún es visible.[2]
Tejido cartilaginoso y ligamentos
Desde la novena vértebra cervical hasta la espalda, las articulaciones vertebrales muestran restos de cápsulas articulares. Entre las espinas en algunos lugares se ven ligamentos interespinales y supraespinales muy delgados. Seis vértebras están visiblemente cubiertas por sincondrosis cartilaginosas, una característica juvenil típica. Los casquetes cartilaginosos también están presentes en todas las articulaciones de las extremidades, incluso en las más pequeñas, y son especialmente gruesas en las articulaciones de los hombros, codos y muñecas. Además, se tapa el pie púbico y el hueso ilíaco y púbico están separados por cartílago.[2]
Sistema respiratorio
Del sistema respiratorio se ha conservado poco. No ha sobrevivido ningún rastro de los pulmones, ni de ningún saco de aire. El único elemento aún presente consiste en una pieza de la tráquea de siete milímetros de largo de la cual se ven unos diez anillos traqueales, los más anteriores de los cuales están abiertos en la parte superior, dándoles forma de C. Tienen una longitud media de 0,33 milímetros y están separados por espacios intermedios de 0,17 milímetros de espesor. La tráquea es bastante delgada, con un ancho conservado de un milímetro aproximadamente la mitad de ancho de lo que se esperaría para un animal del tamaño del holotipo, y está colocada bastante baja en la base del cuello, incrustada en tejido conectivo.[2]
Hígado, corazón, bazo y timo
En la parte anterior del tórax se ve un llamativo halo rojo que forma una mancha más o menos circular de diecisiete milímetros de diámetro. En 1998 se sugirió que esto podría representar los restos del hígado descompuesto, un órgano rico en sangre. En 2011, se confirmó que el pigmento rojo se derivaba de la sangre. Un análisis de microscopio electrónico de barrido indicó que la sustancia consistía en limonita, óxido de hierro hidratado, un probable producto de transformación de la hemoglobina original. También estaba presente biliverdina, un componente biliar esperable en el hígado. La sangre también podría haberse originado en parte en el corazón y el bazo, dos órganos igualmente ricos en sangre, con reptiles colocados entre los dos lóbulos del hígado.[2]
Otro órgano del tórax, del cual pueden existir vestigios, es el timo, que pudo haber contribuido a una masa grisácea de origen orgánico visible en la base del cuello, esto también contiene tejido conectivo y muscular.[2]
Sistema digestivo
El tracto digestivo se puede rastrear principalmente, ya sea porque los intestinos todavía están presentes o por la presencia de alimentos. La posición del esófago está indicada por una serie de pequeñas partículas de alimentos de cinco milímetros de largo. Debajo de la novena vértebra dorsal, la ubicación del estómago se muestra mediante un grupo de huesos de animales de presa y es probable que el propio órgano haya sido disuelto por su propio ácido poco después de la muerte. La posición bastante hacia atrás del grupo sugiere que el estómago era de estructura dual, con un proventrículo que secreta enzimas hacia adelante precediendo a una molleja muscular. No se han reportado gastrolitos.[2]
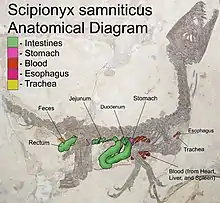
Justo detrás de la supuesta posición del estómago se ve una tripa gruesa y muy llamativa, que se ha identificado como duodeno. Se conserva en parte en forma del molde natural , en parte como una petrificación que aún muestra la estructura celular, incluida la mucosa y el tejido conectivo. Algo mesentérico. Los vasos sanguíneos cubren el intestino en forma de tubos huecos de hasta un centímetro de largo y de 0.02 a 0.1 milímetros de ancho. El duodeno forma un gran bucle, cuya parte descendente se dirige primero hacia las gastralias y luego corre hacia el lomo. Allí, en una curva cerrada, cuyos pliegues son claramente visibles, gira hacia el frente, procediendo como una vía ascendente, su parte visible termina cerca del estómago. En este punto, el tracto se dirige hacia la izquierda del cuerpo, perpendicular a la losa fósil, y por lo tanto ya no se puede seguir su curso. Cerca y ligeramente por encima, emerge una parte posterior del intestino que se ha interpretado como el yeyuno. Este intestino más delgado gira hacia la espalda, corre paralelo al tracto ascendente del duodeno y finalmente desaparece debajo de él, al nivel de la duodécima vértebra dorsal. Aparentemente, se hace un bucle hacia el frente porque resurge debajo de la décima vértebra dorsal, primero corriendo hacia arriba y luego girando hacia atrás debajo de la columna vertebral trasera, o en lugares incluso sobre ella. Probablemente después de la muerte, su posición se desplazó parcialmente hacia arriba. El yeyuno parece fusionarse con un íleon excepcionalmente corto. Una contracción debajo de la decimotercera vértebra dorsal podría indicar la transición al recto. Un ciego parece ausente. El recto corre hacia la parte posterior entre los ejes superiores del pubis y el isquion. Luego se dobla hacia abajo en paralelo al eje del isquion, al final girando hacia arriba nuevamente. En esta parte final todavía están presentes las heces. La cloaca es insuficiente. Dal Sasso & Maganuco sugirieron que la salida de la cloaca era bastante baja, al nivel de los pies isquiáticos y que una válvula rectocoprodal separaba las heces y la orina.[2]
Entre el borde anterior de los ejes púbicos y la parte posterior de los intestinos hay un gran espacio vacío. Además, el recto parece correr en una posición muy alta, como si algo lo empujara hacia arriba. Según Dal Sasso & Maganuco, en vida este espacio habría sido llenado por el saco vitelino de la cría, al nacer, los juveniles de reptiles no han absorbido normalmente toda la yema y utilizan los nutrientes residuales para complementar la ingesta de alimentos durante sus primeras semanas.[2]
Tejido Muscular
En varios lugares el tejido muscular fósil está presente. El grado de conservación es a menudo excepcional, no solo las fibras individuales aún son discernibles, sino también las células individuales e incluso los sarcómeros subcelulares. Entre los fósiles de dinosaurios, estos sarcómeros solo se conocen de Santanaraptor, cuyas fibras musculares son cuatro veces más gruesas. El material orgánico original ha sido reemplazado por pequeños globos huecos, cuyas paredes están formadas por cristales euédricos de apatita.[2]
En la masa orgánica gris en la base del cuello, las fibras musculares están presentes que han sido identificados como pertenecientes al músculo esternohioideo y el músculo esternotraqueal. Entre la sexta y séptima vértebra dorsal es visible un parche de fibras musculares que pertenecen al músculo transversospinal o al músculo longissimus dorsi. Delante del isquion derecho hay fibras musculares que van desde el pie isquiático en dirección al fémur. Su identidad es incierta: podrían pertenecer al Musculus puboisquofemoral pars medialis, el Musculus aductor femoris I de cocodrilos, pero en ese caso este músculo en algunos terópodos no aviares no estaría anclado en el proceso obturador. Las fibras también podrían representar un músculo desconocido. En cualquier caso, refutan una conjetura de Gregory S. Paul de que no habría ninguna conexión muscular entre el isquion y el fémur en absoluto. Por encima del tracto recto hay una gran área de fibras musculares horizontales no segmentadas, probablemente representando el Musculus caudofemoral largo no segmentado de la base de la cola, el principal músculo retractor que opera en el fémur. Estas fibras son poligonales en sección transversal y también muestran los espacios intercelulares. Debajo de algunas vértebras de la base de la cola están presentes los ligamentos conectivos entre los chevrones, formando el ligmamento interhemal, pero también algunas pequeñas fibras musculares y unos misteriosos tubos huecos dispuestos en forma de espiga. Estos últimos quizás representan el myosepta de los myotomes, los segmentos del músculo iliocaudal o el músculo ischiocaudal.[2]
Integumento

En todas las garras conservadas en el fósil, las de los pies se han perdido, las vainas de las garras son visibles. Estos tienen una coloración más oscura en la parte superior que en la parte inferior, lo que sugiere que el material original de la garra todavía está presente, pero esto aún no ha sido probado directamente por un análisis químico por temor a dañar estas delicadas estructuras que se consideraban una parte esencial de la integridad del precioso espécimen. Las vainas córneas de las garras de las manos extienden los núcleos óseos en aproximadamente un 40%, como una guadaña, continuando la curva del hueso y terminando en puntas afiladas. En algunas garras, las vainas se han desprendido parcialmente. En otros, se han aplanado o dividido.[2]
El fósil no conserva rastros de piel, escamas o plumas. En 1999, Philip J. Currie planteó la hipótesis de que esto podría ser de otra manera, sugiriendo que los tubos que se encuentran en la base de la cola representarían los filamentos de las protoplumas. En 2011, sin embargo, Dal Sasso & Maganuco rechazaron esta interpretación porque los tubos se estrechan en ambos extremos, mientras que se espera que los filamentos del tegumento solo tengan un extremo superior ahusado. Sin embargo, consideraron probable que Scipionyx en vida tuviera protoplumas, ya que se sabe que están presentes con los compsognátidos Sinosauropteryx y Sinocalliopteryx.[2]
Descubrimiento e investigación
Scipionyx es considerado uno de los más importante fósiles de vertebrados descubiertos, luego de largos y complejos estudios se comprobó que poseía impresiones fósiles de los órganos internos.[4] Parte de la tráquea, intestinos, hígado, y músculos fueron bien preservados en el limo. El hígado del espécimen está tan bien preservado que retiene la forma y el color original. La importancia del descubrimiento se debe a que la posición de los órganos solo podía suponerse antes de este descubrimiento. El intestino era más corto de lo esperado, indicando que el Scipionyx pudo poseer un eficiente proceso digestivo.

Scipionyx fue descubierto en la primavera de 1981 por Giovanni Todesco, un paleontólogo aficionado , en la pequeña cantera de Le Cavere en las afueras del pueblo de Pietraroja, aproximadamente a 70 kilómetros al noreste de Nápoles.[1] El espécimen se conservó en la marina formación Pietraroja, conocida por sus fósiles inusualmente bien conservados. Todesco pensó que los restos pertenecían a un ave extinta. Preparó el extraño descubrimiento en el sótano de su casa en San Giovanni Ilarione cerca de Verona, retirando, sin el uso de ningún instrumento óptico, parte de la matriz de tiza de la parte superior de los huesos y cubriéndolos con cola vinílica. Fortaleció la placa de piedra agregando piezas a su borde y en una de ellas agregó una cola falsa hecha de resina de poliéster, ya que la del fósil faltaba en gran medida porque no la había recuperado por completo. A principios de 1993, Todesco, quien había apodado al animal cagnolino, "perrito", por sus mandíbulas dentudas, llamó la atención del paleontólogo Giorgio Teruzzi del Museo Civico di Storia Naturale di Milano, quien lo identificó como un juvenil de un dinosaurio terópodo y lo apodó Ambrogio en honor al santo patrón de Milán, Ambrose. No siendo él mismo un experto en el campo de los estudios de dinosaurios, recurrió a la ayuda de su colega, el padre Giuseppe Leonardi. En Italia, estos hallazgos son propiedad del Estado por ley y Todesco fue convencido por el periodista científico Franco Capone de informar del descubrimiento a las autoridades. El 15 de octubre de 1993, Todesco entregó personalmente el fósil a la Dirección de Arqueología en Nápoles. El espécimen fue agregado a la colección de la Soprintendenza per i Beni Archeologici di Salerno, Avellino, Benevento e Caserta en Salerno, a la que oficialmente todavía pertenece. El 19 de abril de 2002 se exhibió en el Museo Archeologico di Benevento.
En 1993, Teruzzi y Leonardi informaron científicamente del hallazgo,[5] que generó cierta publicidad ya que fue el primer dinosaurio encontrado en Italia.[6] La popular revista Oggi apodó simultáneamente al animal Ciro, el nombre típico de un niño napolitano, una idea del editor en jefe Pino Aprile.[7] En 1994 Leonardi publicó un artículo más extenso sobre el descubrimiento.[8] En 1995 Marco Signore de la Universidad de Nápoles Federico II[1] presentó una tesis que contenía una extensa descripción del fósil, en la que lo llamó "Dromaeodaimon irene".[9] Debido a que la tesis no se publicó, esto siguió siendo un nomen ex dissertatione y considerado inválido . Mientras tanto, en Salerno, Sergio Rampinelli había comenzado una nueva preparación del fósil, durante 300 horas de trabajo quitando la cola falsa, reemplazando el pegamento vinílico con un moderno conservante de resina y terminando el destapado de los huesos. En esta ocasión se descubrió que se habían conservado gran parte de los tejidos blandos.

En 1998, Ciro por esto apareció la portada de Nature, cuando la especie tipo Scipionyx samniticus fue nombrada y descrita por Marco Signore y Cristiano Dal Sasso.[10] El nombre genérico Scipionyx proviene del nombre latino Scipio y del griego ὄνυξ, ónix, la combinación que significa "garra de Scipio". "Escipión" se refiere tanto a Scipione Breislak,[1] el geólogo del siglo XVIII que escribió la primera descripción de la formación en la que se encontró el fósil como a Escipión Africano, el famoso cónsul romano que luchó contra Hannibal. El nombre específico samniticus significa "De Samnium ", el nombre latino de la región alrededor de Pietraroja. Se habían considerado varios otros nombres, pero se habían rechazado, como "Italosaurus", "Italoraptor" y "Microraptor".[11] Desde entonces, este último nombre se ha utilizado para un género de dromeosáurido de "cuatro alas" descubierto en China unos años más tarde.
El holotipo, SBA-SA 163760, data del Albiense temprano , tiene unos 110 millones de años, y consiste en un esqueleto casi completo de un individuo juvenil, al que le falta solo el extremo de la cola, la parte inferior de las patas y la garra del segundo derecho. dedo. Se han conservado extensos tejidos blandos, pero ninguna parte de la piel ni ningún tegumento como escamas o plumas.[12] Se cree que el Scipionix vivió en una región de muchos lagos. Por lo que se calcula que el cuerpo cayó en una región pobre de oxígeno, lo que preservó tan bien el espécimen de Scipionyx, de igual manera que el fósil de Archaeopteryx de Alemania. En vista de la importancia excepcional del hallazgo, entre diciembre de 2005 y octubre de 2008 el fósil se estudió intensamente en Milán, lo que resultó en una monografía de dal Sasso y Simone Maganuco publicada en 2011,[2] que contiene la descripción más extensa de una sola especie de dinosaurio. siempre. Scipionyx es clasificado dentro del clado Coelurosauria y por lo tanto es un terópodo. Debido a que solo se recobraron restos de un individuo juvenil, es difícil asignarlo a un infraorden en particular. Las características de celurosaurioano incluyen el sacro largo más que en otros dinosaurios, una cola rígida, y una ulna combada. La tibia es tan larga como el fémur. En 2021, el paleontólogo italiano Andrea Cau propuso que el holotipo de Scipionyx es un carcharodontosáurido recién nacido.[13]
Clasificación
Scipionyx fue asignado por sus descriptores a Coelurosauria, un grupo de terópodos. Debido a que sus únicos restos conocidos pertenecen a un juvenil, ha sido difícil asignar a este dinosaurio a un grupo particular. El problema yace en que su constitución inmadura es más probable que se expresen rasgos pertenecientes a sus antecesores, lo que sugeriría a su vez una posición más primitiva en un árbol evolutivo de lo que fue. Parte de la monografía realizada sobre este género en 2011 fue in análisis cladístico el cual indicaba que Scipionyx era un miembro basal de la familia Compsognathidae y la especie hermana de Orkoraptor. Dal Sasso & Maganuco enfatizaron que, debido a sus restos escasos, la posición de Orkoraptor es tentativa.[2] Otra hipótesis es que Scipionyx es un carcharodontosaurio recién nacido.[13]
Filogenia
Este cladograma muestra la posición filogenética de Scipionyx entre los celurosaurios, de acuerdo con el estudio de 2011.

| Tyrannoraptora |
| ||||||||||||||||||||||||||||||||||||||||||||||||
Paleoecología

La ubicación donde se encontró Scipionyx, en el Albiano, era parte de la Placa de Apulia, en ese momento cubierta en gran parte por el mar poco profundos de Paratethys. Sin embargo, había algo de tierra seca, pero no se sabe cuán extensos o conectados estaban los varios terrenos. Los sedimentos marinos de la formación Pietraroja probablemente se depositaron cerca de un trozo de la Plataforma de los Apeninos, que posiblemente formaba una pequeña isla entre el actual centro de Italia y Túnez. De esto se ha concluido que el hábitat de Scipionyx en general consistía en pequeñas islas y representaba uno de los animales más grandes de su ecosistema.[2]
Sin embargo, también hay indicios de que los terrenos se interconectan regularmente para formar islas mucho más extensas, puentes terrestres que permiten la dispersión de animales mucho más grandes, como saurópodos y grandes terópodos. De ser así, no estuvieron presentes por mucho tiempo cuando la superficie terrestre se fragmentó nuevamente, porque no hay signos de enanismo insular, una reducción de tamaño como una adaptación a la disminución de los recursos. Del mismo modo, Scipionyx en sí no es un enano entre sus parientes. Debido a su pequeño tamaño absoluto, Scipionyx habría podido mantenerse cuando la tierra seca se redujo. Sin embargo, Dal Sasso & Maganuco no consideraron a Scipionyx como un residente permanente de pequeñas islas durante decenas de millones de años, pero más probablemente un inmigrante reciente que llegó durante una ola de dispersión, probablemente del norte de África. Admitieron que esto estaba en desacuerdo con su propio análisis filogenético, mostrando que Scipionyx es un compsognátido basal, pero señalaron que la filogenia encontrada era incierta debido al estado juvenil del fósil.[2]
Los animales terrestres que se encuentran realmente en los depósitos de Pietraroja son todos pequeños. Entre ellos se encuentran los lagartos Chometokadmon y Eichstaettisaurus gouldi, un pariente del alemán Eichstaettisaurus schroederi cuarenta millones de años mayor, el rinchocefálo Derasmosaurus y el anfibio Celtedens megacephalus.[2]
Paleobiología
Alimentación
El fósil proporciona información directa sobre la dieta de Scipionyx porque se han conservado restos de una serie completa de comidas consecutivas, quizás todo lo que el animal comió durante su corta vida. Estos confirman lo que ya se pudo concluir a partir de sus afinidades filogenéticas y estructura general, de que Scipionyx era un depredador.[2]
En el tracto del esófago se encuentran presentes unas ocho escamas y algunos fragmentos óseos. Dal Sasso & Maganuco consideraron probable que estos no hubieran sido ingeridos como elementos sueltos sino como restos de una comida, en parte regurgitados desde el estómago en la agonía final. En la propia posición del estómago, se ve un grupo de huesos pequeños. Estos incluyen un tobillo con un metatarso de tres milímetros de ancho que consta de cinco metatarsianos unidos, una vértebra de la cola y el extremo superior de un cúbito. Si los restos representan un solo animal de presa, es probable que sea un miembro de la Mesoeucrocodylia o algún lepidosaurio, un animal parecido a un lagarto. El tamaño indica la última posibilidad. En el tracto descendente del duodeno están presentes dos grupos de escamas de lagarto y, más abajo, una vértebra de pez. El yeyuno muestra un grupo de docenas de vértebras de peces, probablemente pertenecientes a un miembro de Clupeomorpha. Se encontró un segundo grupo de vértebras en el límite yeyuno-íleon. El tracto final del recto todavía contiene heces en las que se ve un trozo de piel que muestra diecisiete escamas de un pez Osteoglosiformes que tenía nueve temporadas, a juzgar por las líneas de crecimiento en las escamas.[2]
.jpg.webp)
Los alimentos encontrados permiten reconstruir una secuencia de ingestas de alimentos. Primero un pez de cuatro a cinco centímetros de largo, en segundo lugar, un pez más pequeño de dos a tres centímetros, luego un lagarto de diez a doce centímetros de largo, luego una lagartija lepidosauriana de quince a cuarenta centímetros de largo, según la identificación y finalmente algunos vertebrados indeterminados. Juntos representan una dieta variada que muestra que Scipionyx era un generalista oportunista. El hecho de que se hubieran capturado lagartos veloces y que se hubieran recolectado peces de mar arrastrados a la orilla requiriendo un patrullaje prolongado de la línea de inundación, ambos indican una buena movilidad. Si el animal de presa en el estómago realmente tenía cuarenta centímetros de largo, es muy poco probable que la cría del mismo tamaño hubiera podido someterlo, lo que indica el cuidado de los padres.[2]
Fisiología
Scipionyx es considerado uno de los vertebrados fósiles más importantes jamás descubiertos, después de que una larga y minuciosa "autopsia" revelara la fosilización única de partes de sus órganos internos. Se cree que Scipionyx vivía en una región llena de lagunas poco profundas. Estos cuerpos de agua tenían deficiencia de oxígeno, lo que llevó al espécimen de Scipionyx sea bien conservado, muy parecido a la excelente conservación de fósiles que se observa en el Archaeopteryx de Alemania.[1][14] Partes de la tráquea , los intestinos , el hígado, vasos sanguíneos, cartílagos, vainas córneas, tendones y músculos estaban fosilizados en la fina piedra caliza. El hígado del espécimen se conservó en forma de halo rojo de hematita conservando la forma que tenía cuando el animal estaba vivo. El hallazgo tiene gran importancia porque las posiciones relativas de los órganos internos de los dinosaurios solo podían adivinarse antes de este descubrimiento. El espécimen holotipo proporciona así información directa única sobre la fisiología de los dinosaurios no aviares, especialmente en lo que respecta a la digestión, respiración y ontogénesis.
Digestión
El tracto digestivo de Scipionyx es en generalcorto pero ancho. La longitud total de los intestinos, más corta de lo esperado, indica que Scipionyx podría procesar los alimentos de manera muy eficiente.[1] La eficacia mejoraría con los pliegues intestinales visibles, las plicae circulares, agrandando la superficie de absorción. Dal Sasso & Maganuco enfatizó que un tramo corto no necesariamente implica que el tiempo de procesamiento también fue corto, la retención podría haberse prolongado para optimizar la digestión. La mayoría de los depredadores vertebrados existentes son capaces de extraer aproximadamente el 75% del valor energético de la carne de la presa.[2]
Los huesos de la región del estómago no habían sido afectados por el ácido del estómago, lo que indica que esta comida tenía menos de un día. Es posible que la salida del estómago fuera demasiado estrecha para dejar pasar estos restos y que el ácido se haya utilizado para extraer calcio de los huesos, un nutriente muy necesario para un animal joven en crecimiento. El tracto digestivo subsiguiente con la mayoría de los vertebrados existentes es incapaz de digerir más huesos. Posteriormente, los restos habrían sido regurgitados. Sin embargo, la presencia de vértebras en los intestinos sugiere que la salida era amplia y que los terópodos no aviares a este respecto se parecían más a los actuales Lepidosauromorpha que a los actuales Archosauria que regurgitan. Esto también se confirma por un alto contenido de hueso en los coprolitos.de grandes terópodos. El pescado contiene mucho calcio y es posible que la cría lo haya buscado instintivamente, alternativamente, podría haber sido alimentado preferentemente con pescado por sus padres. Era de esperar la presencia de piel en las heces, ya que no se digiere fácilmente.[2]
Las partes del cuerpo del gran lepidosaurio en el estómago se hicieron más digeribles al morderlas en pedazos. Según Dal Sasso & Maganuco, la cría sin duda habría sido incapaz de lograr esto y lo consideraron un fuerte indicio de cuidado parental, ya que era improbable que el animal hubiera encontrado por casualidad un cadáver perfectamente desgarrado en pedazos fáciles de tragar por depredadores o carroñeros.[2]
Respiración
Aunque la mayor parte del sistema respiratorio no se ha conservado, se han extraído algunas conclusiones de gran alcance a partir de la evidencia indirecta disponible. En 1999 John Ruben ea infirió que Scipionyx tenía un sistema respiratorio diferente al de las aves y más similar al de los cocodrilos, basándose en un análisis de fotografías del fósil que parecían indicar la presencia de un diafragma. El hígado grande habría dividido completamente la cavidad corporal en una sección anterior para el corazón y los pulmones y una sección posterior para los intestinos. Esto habría indicado la presencia de pulmones septados, ventilados por un diafragma de pistón hepático, impulsado por el hígado y un músculo diafragmático, que en el fósil estaba visiblemente adherido al pubis. Tal sistema sería un argumento en contra de la idea de que las aves, cuyos pulmones están ventilados por sacos de aire, son terópodos celurosaurios y una indicación de que los terópodos eran de sangre fría.[15]
Las conclusiones de John Ruben, sin embargo, han sido cuestionadas por algunos científicos, como Lawrence Witmer, quien afirmó que el estudio tenía fallas.[16] El estudio de 2011 concluyó que debido al hecho de que el hígado se había conservado como un halo vago, que representa los fluidos corporales que después de la muerte podrían haber cubierto una superficie más grande que los órganos de los que se originaron, no se pueden determinar sus dimensiones y extensión exactas. En cualquier caso, un diafragma en sí o su posición no se pudieron observar directamente. Muchos hígados de aves también son grandes, lo que demuestra que tal rasgo es compatible con un sistema de sacos de aire. La pequeña cavidad del cuerpo frente al halo parecía indicar la presencia de pequeños pulmones rígidos como de pájaro. El presunto M. diafragmatico se demostró que era un artefacto causado por el pulido y grabado de nódulos de calcita de origen no orgánico durante la preparación, creando la ilusión de fibras musculares.[2]
G. S. Paul y David Martill han planteado la hipótesis de que el espacio vacío entre los huesos púbicos y los intestinos era la ubicación de un gran saco de aire. Dal Sasso & Maganuco, sin embargo, rechazaron esta interpretación porque en las aves vivas el saco aéreo del abdomen posterior no fuerza a los intestinos hacia adelante. Consideraron que era más probable que el espacio estuviera ocupado por un gran saco vitelino. No obstante, los sacos de aire eran probables dada la neumatización de las vértebras. Las vértebras sin pneumatoporos habrían indicado los límites entre tres sistemas de sacos aéreos, los de la base del cuello, los pulmones y el abdomen. Las cabezas de doble costilla indicarían un tórax bastante rígido, ventilado por la gastralia. Un sistema de procesos uncinados en forma de gancho en las costillas como con los Maniraptoriformes, permitiendo que la caja torácica se mueva de manera flexible, en articulación con un esternón osificado, estaba ausente en Scipionyx.[2]
Crecimiento
El holotipo de Scipionyx es un raro ejemplo de cría de terópodo no aviar. Los otros ejemplares muy jóvenes más importantes son los polluelos de Byronosaurus que sin embargo son mucho menos completos. La juventud se refleja en las proporciones y el bajo grado de osificación y fusión de varios elementos esqueléticos. El rasgo juvenil más obvio es la cabeza relativamente grande y corta.[2]
Dal Sasso & Maganuco han intentado determinar la edad absoluta de la cría. El hecho de que la fontanela aún no se haya cerrado, plantea un límite superior de edad de unas cinco semanas. Un límite aún más bajo está indicado por la falta de reemplazo de dientes, que con Archosauria comienza después de unas pocas semanas a más tardar. La edad más exacta viene dada por el tamaño del saco vitelino, que indica una edad probable de tres días, con un límite superior de una semana. A pesar de su corta edad, la cría podía caminar, como lo demuestra la completa osificación del hueso ilíaco. Sin embargo, esto no implica que Scipionyx fuera precoz, ya que incluso en las aves altriciales este hueso de la pelvis se osifica por completo unos días después de la eclosión.[2]
Referencias
- Poling, Jeff (1998). «Skippy the dinosaur». Journal of Dinosaur Paleontology. Archivado desde el original el 4 de febrero de 2007. Consultado el 1 de marzo de 2007.
- Cristiano dal Sasso & Simone Maganuco, 2011, Scipionyx samniticus (Theropoda: Compsognathidae) from the Lower Cretaceous of Italy — Osteology, ontogenetic assessment, phylogeny, soft tissue anatomy, taphonomy and palaeobiology, Memorie della Società Italiana de Scienze Naturali e del Museo Civico di Storia Naturale di Milano XXXVII(I): 1-281
- Carrano, M.T. & Sampson S.D. 2008. "The phylogeny of Ceratosauria". Journal of Systematic Palaeontology 6(2): 183-236
- Dal Sasso, C. and Signore, M. (1998). "Exceptional soft tissue preservation in a theropod dinosaur from Italy." Nature, 392: 383-387.
- Leonardi, G. & Teruzzi, G., 1993, "Prima segnalazione di uno scheletro fossile di dinosauro (Theropoda, Coelurosauria) in Italia (Cretacico di Pietraroia, Benevento)", Paleocronache 1993: 7-14
- Dal Sasso, C. and Signore, M., 1998, "Scipionyx samniticus (Saurischia, Theropoda): the first Italian dinosaur", Third European Workshop on Vertebrate Paleontology, Abstract: 23
- Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages.
- Leonardi, G. & Avanzini, M., 1994, "Dinosauri in Italia", Le Scienze (Quaderni), 76: 69-81
- Signore, M., 1995, Il teropode del Plattenkalk della Civita di Pietraroia (Cretaceo inferiore, Bn). Thesis, Dip. Paleont. Univ. Napoli "Federico II"
- Dal Sasso, C. and Signore, M. (1998). "Exceptional soft tissue preservation in a theropod dinosaur from Italy." Nature, 392: 383-387.
- Cristiano Dal Sasso & Giuseppe Brillante, 2001, Dinosauri italiani, Marsilio pp 256
- Dal Sasso, C. and Signore, M., 1998, "Scipionyx samniticus (Theropoda: Coelurosauria) and its exceptionally well preserved internal organs", Journal of Vertebrate Paleontology 18 (3): 37A
- Cau, Andrea (2021). «Comments on the Mesozoic theropod dinosaurs from Italy». Atti della Società dei Naturalisti e Matematici di Modena 152: 81-95.
- Reisdorf, A.G., and Wuttke, M. (2012). "Re-evaluating Moodie's Opisthotonic-Posture Hypothesis in fossil vertebrates. Part I: Reptiles - The taphonomy of the bipedal dinosaurs Compsognathus longipes and Juravenator starki from the Solnhofen Archipelago (Jurassic, Germany)." Palaeobiodiversity and Palaeoenvironments, doi 10.1007/s12549-011-0068-y
- John A. Ruben, Cristiano Dal Sasso, Nicholas R. Geist, Willem J. Hillenius, Terry D. Jones and Marco Signore (1999), «Pulmonary Function and Metabolic Physiology of Theropod Dinosaurs», Science 283 (5401): 514-516, Bibcode:1999Sci...283..514R, PMID 9915693, doi:10.1126/science.283.5401.514Abstract and citations at Science journal site.
- Browne, Malcolm W (26 de enero de 1999), «Spectacular fossil reveals dinosaur soft part anatomy and supports cold-blooded dinos not related to birds», New York Times, archivado desde el original el 11 de mayo de 2012, consultado el 11 de noviembre de 2021.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikispecies tiene un artículo sobre Scipionyx samniticus.
Wikispecies tiene un artículo sobre Scipionyx samniticus. Wikimedia Commons alberga una categoría multimedia sobre Scipionyx samniticus.
Wikimedia Commons alberga una categoría multimedia sobre Scipionyx samniticus.- "Baby dinosaur from Italy with guts preserved"