Sinornithosaurus
Sinornithosaurus ("Ave-reptil chino") es un género con dos especies conocidas de dinosaurios terópodos dromeosáuridos, que vivieron a principios del período Cretácico, hace aproximadamente entre 124,6 a 122 millones de años, durante el Aptiense, en lo que es hoy Asia. Encontrado en la Formación Yixian de China.[1] Cuando se encontró la primera especie, Sinornithosaurus millenii, en 1999 causó gran sensación porque era el quinto dinosaurio no aviar con plumas descubierto. Fue recogido del sitio de Sihetun en Liaoning occidental. Estaba en la capa 6, Chaomidianzi, en la porción inferior de Yixian, en el Grupo Jehol. La segunda especie Sinornithosaurus haoiana fue descrita en 2004.
| Sinornithosaurus | ||
|---|---|---|
| Rango temporal: 124,6 Ma - 122 Ma Cretácico inferior (Aptiense) | ||
 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| Infraorden: | Coelurosauria | |
| (sin rango): | Maniraptora | |
| Superfamilia: | Deinonychosauria | |
| Familia: | Dromaeosauridae | |
| Subfamilia: | Microraptorinae | |
| Género: |
Sinornithosaurus Xu, Wang y Wu, 1999 | |
| Especie tipo | ||
|
Sinornithosaurus millenii Xu, Wang, & Wu, 1999 | ||
| Otras Especies | ||
| ||
Descripción
Como todos los dromeosáuridos, estaba muy estrechamente emparentado con las aves, aunque era incapaz de volar. Sinornithosaurus estaba entre los dromeosáuridos más pequeños, con una longitud de unos 90 centímetros.[2] En 2010, Gregory S. Paul dio estimaciones más altas de 1,2 metros y tres kilogramos.[3] Sus brazos y manos eran proporcionalmente muy largos. La capa de protoplumas que cubría su piel constaba de dos tipos de estructuras, fibras simples organizadas en haces y plumas con un eje central y barbas, pero sin las bárbulas de las plumas más evolucionadas. Sinornithosaurus sugiere que otros dromeosáuridos corredores, como el Velociraptor, también pudieron haber poseído plumas.
Descubrimiento e investigación

Sinornithosaurus fue descubierto por Xu Xing, Wang Xiaolin y Wu Xiaochun del Instituto de Paleontología de Vertebrados y Paleoantropología de Pekín. Un casi completo fósil con impresiones de plumas, recobrado de la Provincia de Liaoning, China, en la Formación Yixian. En está formación, que increíblemente rica, previamente habían aparecido otros cuatro dinosaurios con plumas, Protarchaeopteryx, Sinosauropteryx, Caudipteryx, y Beipiaosaurus.
Los restos fósiles fueron hallados en la Formación Yixian, en China. Se han descrito dos especies: Sinornithosaurus millenii en el año 1999 y Sinornithosaurus haoiana en el 2004. Existe también un ejemplar pequeño que quizá termine asignándose también al género Sinornithosaurus. Este fósil, probablemente una cría, se identifica como NGMC 91, y ha sido apodado "Dave". Está completo y magníficamente preservado, y de sus brazos surgen largas plumas que forman aparentemente un ala o protoala.
El holotipo IVPP V12811, se encuentra en la colección del Instituto de Paleontología de Vertebrados y Paleoantroplogía en Pekín, China. Otro espécimen, NGMC-91, es probablemente un juvenil Sinornithosaurus. Qiang et al. en 2001 eran renuente nombrar NGMC-91 debido a que aunque el espécimen este articulado totalmente, casi todos los huesos se rompieron cuando las losas con los fósiles se partieron, de modo que solamente las siluetas de los huesos están claras en la mayor parte de la pieza y las contrapartes. Esto obscureció las características esqueléticas de diagnóstico, que hicieron el especien de género incierto.[4] NGMC-91, también llamado "Dave", se encuentra en la colección del Museo Geológico Nacional de. Fue recogido de la mina Fanzhangzi, cerca de la ciudad de Lingyuan, Provincia de Liaoning, China. Esta se encuentra aproximadamente 130 kilómetros de la localidad de Sihetun. Un espécimen de un pez Lycoptera también se encuentra cerca del pie de NGMC-91.
En 2009, un equipo de científicos liderado por Enpu Gong examinó un cráneo bien conservado de Sinornithosaurus y encontró evidencias que sugerían que se trata del primer dinosaurio venenoso identificado. Gong et al. sugieren que Sinornithosaurus podría haberse especializado en cazar presas pequeñas, por ejemplo pájaros, como hacen las serpientes actualmente.[5]
Clasificación
Xu Xing describe a Sinornithosaurus y presenta un análisis filogenético en el que demuestran que se trata de un basal, o primitivo, dromeosáurido.[6] ellos demostraron que algunas características del hombro y cráneo son muy similares a "Archaeopteryx" y otros Avialae. Estas particularidades juntas demuestran que los primeros dromeosáuridos eran más parecidos a las aves que los últimos. Esto contradice un argumento de los críticos de la teoría de que los pájaros se desarrollaron a partir de dinosaurios.[7]

Sinornithosaurus es miembro de la familia Dromaeosauridae, un grupo de ágiles dinosaurios cazadores con una distintiva garra en forma de hoz en cada pie; éstos incluyen a Deinonychus y Utahraptor. Sinornithosaurus vivió hace 125 millones de años en el Barremiense (Cretácico Inferior), lo que lo hace uno de los más antiguos y primitivos miembros de esta familia. La presencia de plumas en Sinornithosaurus es consistente con lo visto en otros dromeosáuridos como Microraptor, Velociraptor y Rahonavis.

Sinornithosaurus es conocido por al menos dos especies. S. millenii, "lagarto ave chino del milenio", es la especie tipo, descrito en 1999. Una segunda especie S. haoiana, "lagarto ave chino de Hao ", fue descrita por Liu et al. en 2004 basándose en un espécimen que difiere de S. millenii en caracteres de la cabeza y cadera.[8] Un increíblemente bien preservado microrraptorino "Dave", espécimen NGMC 91, podría representar una tercera especie de Sinornithosaurus, o un juvenil. Sin embargo, según Turner, Makovicky y Norell en 2012, las supuestas características distintivas de S. haoiana "están presentes en Sinornithosaurus millenii o son variables entre el número de especímenes de Sinornithosaurus ". Los autores consideraron S. haoiana como un sinónimo menor de S. millenii.[9]
Un microraptorino increíblemente bien conservado apodado "Dave", espécimen NGMC 91, fue descrito por primera vez en un artículo publicado en la revista Nature por Ji Qiang y sus colegas en 2001. Se negaron a nombrar el espécimen porque, aunque está completamente articulado, casi todos los huesos se rompieron cuando se partieron las losas fósiles, por lo que solo las siluetas de estos huesos son claras en la mayor parte de la parte y la contraparte. Esto oscureció las características esqueléticas de diagnóstico, lo que hizo que el género del espécimen fuera incierto. Notaron que era similar en algunos aspectos a Sinornithosaurus millenii, y sugirieron que las diferencias entre los dos podrían deberse a la edad.[4] Ji, junto con otro equipo de científicos, enfatizó aún más esta similitud en un artículo de 2002, en el que formalmente refirieron el espécimen a Sinornithosaurus , aunque consideraron cuestionable la especie exacta.[10] Mientras tanto, Stephen Czerkas y sus colegas consideraron que el espécimen representaba un ejemplo de su especie recién descrita Cryptovolans pauli, ahora generalmente considerada un sinónimo de Microraptor gui, basándose en las supuestas proporciones de las alas.
Los estudios filogenéticos no respaldaron la idea de que NGMC 91 fuera un pariente cercano de S. millenii. En un análisis de 2004, Phil Senter y sus colegas encontraron que, de hecho, estaba más estrechamente relacionado con Microraptor.[11] Estudios posteriores, también realizados por Senter, han seguido demostrando apoyo a este hallazgo a pesar de que posteriormente se descubrió que algunos datos utilizados en el estudio original eran erróneos.[12]
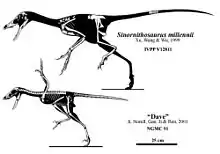
Sin embargo, en una publicación de 2011, Senter declaró que un examen personal del holotipo de S. millenii lo llevó a concluir que su separación anterior de NGMC 91 de S. millenii se basó en interpretaciones anatómicas erróneas. Como los dos especímenes eran "idénticos en todos los estados de caracteres" utilizados en su análisis filogenético, eran de niveles estratigráficos similares y "compartían de forma única la presencia de una coracoides triangular con un gran foramen ovalado", Senter concluyó que NGMC 91 pertenece a la especie Sinornithosaurus millenii.[13]
Turner, Makovicky y Norell, en 2012, también llegaron a la misma conclusión independientemente de Senter. Según estos autores, NGMC 91 comparte varias apomorfias tanto con Microraptor zhaoianus como con Sinornithosaurus millenii, sin embargo, como "carece de caudales medios alargados que son de tres a cuatro veces la longitud de las vértebras dorsales ", no se puede referir a M. zhaoianus. Por otro lado, posee un dentario bifurcado posteriormente, que es una apomorfia de Sinornithosaurus. Los autores concluyeron que NGMC 91 era un espécimen subadulto de S. millenii.[9]
Paleobiología
Las comparaciones entre los anillos esclerales de Sinornithosaurus y las aves y reptiles modernos indican que pudo haber sido catemeral, activo durante el día y la noche a intervalos cortos.[14]
Plumas

Los especímenes de Sinornithosaurus poseen impresiones de plumas que cubren el contorno del cuerpo y forman las alas. Estas plumas eran indistinguibles de las halladas en las aves de los mismos depósitos geológicos.[15] Las plumas corporales medían generalmente entre 3 a 4,5 centímetros, estaban compuestas de filamentos, y mostraron dos características que indican que son plumas tempranas. Primero, varios filamentos fueron ensamblados juntos en " penachos". En segundo lugar, una fila de los filamentos (barbas) fueron ensamblados junto a un eje (raquis), haciéndolos similares en estructura normal de la pluma de los pájaros. Sin embargo, no tienen la ramificación secundaria y los pequeños ganchos minúsculos (barbulas) que las plumas modernas tienen, lo que permite que las plumas de las aves modernas formen una paleta continúa. En NGMC - 91 las plumas cubrieron el cuerpo entero, incluyendo la cabeza por delante de los ojos, el cuello, brazos, plumas largas en los muslos, y una la cola como la de Archaeopteryx.[15] Algunos científicos han sugerido que las plumas pueden haberle permitido a Sinornithosaurus planear por breves trayectos tras saltar de los árboles.[16]
Color
Un estudio de 2010 indicó que Sinornithosaurus puede haber tenido plumas que variaban en color significativamente a través de distintas zonas del cuerpo, basándose en análisis de estructuras celulares microscópicas en los fósiles preservadas.[17] Un estudio posterior de 2012 mostró que los colores de Sinornithosaurus eran pardo-rojizo, amarillo, negro y gris, los cuales estaban dispersos por su cuerpo.[16]
Locomoción aérea
La posibilidad de que Sinornithosaurus fuera capaz de planear se ha presentado varias veces, debido a su estrecha relación con dromeosaurios voladores o planeadores como Microraptor. Chatterjee y Templin en 2004 encontraron que S. millenii se agrupaba dentro de los dinosaurios con potencial aerodinámico para la locomoción aérea,[18] una última opinión también expresada por Darren Naish,[16] mientras que Longrich y Currie 2009 expresaron que probablemente era demasiado pesado para volar,[19] aunque vale la pena señalar que este último estudio se publicó antes de la descripción formal de Changyuraptor, una microraptorina más grande con evidentes capacidades de vuelo.[20]
Posible mordida venenosa
En 2009, un equipo de científicos dirigido por Empu Gong examinó un cráneo de Sinornithosaurus bien conservado y notó varias características que sugerían que era el primer dinosaurio venenoso identificado. Gong y sus colegas notaron que los dientes inusualmente largos y con forma de colmillo de la mitad de los maxilares tenían surcos prominentes que bajaban por la superficie exterior, hacia la parte posterior del diente, una característica que solo se ve en animales venenosos. También interpretaron una cavidad en el hueso de la mandíbula justo encima de estos dientes como el sitio posible para la glándula venenosa de tejido blando. Gong y sus colegas sugirieron que estas características únicas indicaban que Sinornithosaurus puede haberse especializado en cazar presas pequeñas como aves, usando sus largos colmillos para penetrar las plumas y envenenar y aturdir a la presa, como una serpiente moderna. También sugirieron que los dientes cortos que apuntan ligeramente hacia adelante en la punta de la mandíbula podrían haber sido utilizados para despojar a las aves de las plumas.[5]

Sin embargo, en 2010, otro equipo de científicos dirigido por Federico Gianechini publicó un artículo que ponía en duda la afirmación de que Sinornithosaurus era venenoso. Señalaron que los dientes ranurados no son exclusivos de este género y, de hecho, los dientes ranurados se encuentran en muchos otros terópodos, incluidos otros dromeosáuridos. También demostraron que los dientes no eran anormalmente largos como afirmaban Gong y su equipo, sino que se habían salido de sus alvéolos, un artefacto de conservación común en los fósiles triturados y aplanados. Finalmente, no pudieron verificar de forma independiente la presencia de supuestas cámaras para las glándulas venenosas citadas por el equipo de Gong, encontrando solo los senos normales del cráneo.[21]
En la misma edición de la revista, Gong y su equipo presentaron una nueva evaluación del estudio de 2010, poniendo en duda sus hallazgos. Admitieron que los dientes acanalados eran comunes entre los terópodos, aunque sugirieron que en realidad solo prevalecían entre los maniraptoranos emplumados, y plantearon la hipótesis de que el veneno puede haber sido un rasgo primitivo para todos los arcosaurios, si no para todos los reptiles, que se mantuvo en ciertos linajes. También cuestionaron la afirmación de que los dientes estaban significativamente fuera de sus alvéolos en el espécimen holotipo de Sinornithosaurus, aunque admitieron que no estaban en una posición completamente natural. La revaluación de Gong también afirmó que ciertos especímenes no descritos tenían dientes completamente articulados que mostraban una longitud similar.[22] Sin embargo, estos dientes ranurados no son evidencia directa de veneno, ya que las especies de animales no venenosos, como los babuinos, tienen dientes ranurados de manera similar.[23]
Referencias
- Swisher, Carl C., Wang, Yuan-qing, Wang, Xiao-lin, Xu, Xing, Wang, Yuan. (1999). "Cretaceous age for the feathered dinosaurs of Liaoning, China". Nature 400:58-61 1 July 1999.
- Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2010 Appendix.
- Paul, G.S., 2010, The Princeton Field Guide to Dinosaurs, Princeton University Press p. 132
- Qiang, Ji, Norell, Mark A., Gao, Ke-Qin, Ji, Shu-An, Ren, Dong. (2001) "The distribution of integumentary structures in a feathered dinosaur" "Nature" 410:1084-1087 26 April 2001.
- Gong, E., L.D. Martin, D.E. Burnham, and A.R. Falk. (2009). "The birdlike raptor Sinornithosaurus was venomous." Proceedings of the National Academy of Sciences, (Published online before print December 14, not yet published in paper)
- Xu, Xing, Wang, Xiao-Lin, Wu, Xiao-Chun. (1999) "A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China" "Nature" 401:262-266 16 September 1999.
- Xu, Xing, Wu, Xiao-Chun. (2001). "Cranial morphology of Sinornithosaurus millenii Xu et al. 1999 (Dinosauria:Theropoda:Dromaeosauridae) from the Yixian Formation of Liaoning, China". Canadian Journal of Earth Sciences 38:1739-1752 (2001)
- Liu, J., Ji, S., Tang, F. & Gao, C. (2004). «A new species of dromaeosaurids from the Yixian Formation of western Liaoning». Geological Bulletin of China 23 (8): 778-783. (abstract Archivado el 28 de septiembre de 2007 en Wayback Machine.)
- Alan Hamilton Turner, Peter J. Makovicky y Mark Norell (2012). "Una revisión de la sistemática de dromeosáuridos y la filogenia paraviana". Boletín del Museo Americano de Historia Natural. 371: 1–206. doi:10.1206/748.1
- Ji, Q.; Ji, S.-A.; Yuan, C.-X.; Ji, X.-X. (2002). «Restudy on a small dromaeosaurid dinosaur with feathers over its entire body». Earth Science Frontiers 9 (3): 57-63.
- Senter, Phil; Barsbold, R.; Britt, Brooks B.; Burnham, David B. (2004). «Systematics and evolution of Dromaeosauridae (Dinosauria, Theropoda)». Bulletin of the Gunma Museum of Natural History 8: 1-20.
- Senter, P. (2007). «A new look at the phylogeny of Coelurosauria (Dinosauria: Theropoda)». Journal of Systematic Palaeontology 5 (4): 429-463. S2CID 83726237. doi:10.1017/S1477201907002143.
- Senter, Phil (2011). «Using creation science to demonstrate evolution 2: morphological continuity within Dinosauria (supporting information)». Journal of Evolutionary Biology 24 (10): 2197-2216. PMID 21726330. doi:10.1111/j.1420-9101.2011.02349.x. Parámetro desconocido
|doi-access=ignorado (ayuda) - Schmitz, L.; Motani, R. (2011). «Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology». Science 332 (6030): 705-8. Bibcode:2011Sci...332..705S. PMID 21493820. S2CID 33253407. doi:10.1126/science.1200043.
- Xu, X.; Zhou, Z.; Prum, R.O. (2001). «Branched integumental structures in Sinornithosaurus and the origin of feathers». Nature 410 (6825): 200-204. PMID 11242078. doi:10.1038/35065589.
- Naish, Darren (2012). Planet Dinosaur : The Next Generation of Killer Giants. Firefly Books. p. 192. ISBN 978-1-77085-049-1.
- Zhang, Fucheng; Kearns, Stuart L.; Orr, Patrick J.; Benton, Michael J.; Zhou, Zhonghe; Johnson, Diane; Xu, Xing and Wang, Xiaolin (2010). Fossilized melanosomes and the colour of Cretaceous dinosaurs and birds" Nature 463(7284), p. 1075. http://oro.open.ac.uk/22432/2/41064696.pdf
- Chatterjee, S., and R. J. Templin. 2004. Feathered coelurosaurs from China: new light on the arboreal origin of avian flight. In: Feathered dragons (P. J. Currie, E. B. Koppelhus, M. A. Shugar, and J. L. Wright, eds.), pp. 251-281. Indiana University Press, Bloomington, IN.
- Longrich, N.R.; Currie, P.J. (2009). «A microraptorine (Dinosauria–Dromaeosauridae) from the Late Cretaceous of North America». Proceedings of the National Academy of Sciences 106 (13): 5002-5007. Bibcode:2009PNAS..106.5002L. PMC 2664043. PMID 19289829. doi:10.1073/pnas.0811664106. Parámetro desconocido
|doi-access=ignorado (ayuda) - Gang Han, Luis M. Chiappe,Shu-An Ji, Michael Habib,Alan H. Turner, Anusuya Chinsamy, Xueling Liu & Lizhuo Han (15 de julio de 2014). «A new raptorial dinosaur with exceptionally long feathering provides insights into dromaeosaurid flight performance». Nature Communications. 5, Article number: 4382. Consultado el 15 de julio de 2014.
- Gianechini, F.A.; Agnolín, F.L.; Ezcurra, M.D. (2010). «A reassessment of the purported venom delivery system of the bird-like raptor Sinornithosaurus». Paläontologische Zeitschrift 85: 103-107. S2CID 84259940. doi:10.1007/s12542-010-0074-9.
- Gong, E.; Martin, L.D.; Burnham, D.A.; Falk, A.R. (2010). «Evidence for a venomous Sinornithosaurus». Paläontologische Zeitschrift 85: 109-111. S2CID 129716572. doi:10.1007/s12542-010-0076-7.
- «Re: Venom in Sinornithosaurus». Archivado desde el original el 10 de noviembre de 2011.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikispecies tiene un artículo sobre Sinornithosaurus.
Wikispecies tiene un artículo sobre Sinornithosaurus. Wikimedia Commons alberga una categoría multimedia sobre Sinornithosaurus.
Wikimedia Commons alberga una categoría multimedia sobre Sinornithosaurus.- New Chinese dromaeosaur covered with downy fibers: Sinornithosaurus millenii, de Zoom Dinosaurs at Enchanted Learning, September 24, 1999. (para niños)
- Wang, L. (10 de marzo de 2001). Dinosaur fossil yields feathery structures. Science News. Archivado desde el original el 16 de julio de 2007. Consultado el 27 de mayo de 2006.
- Sinornithosaurus: "Chinese bird-lizard", por T. Mike Keesey from The Dinosauricon. (técnico)
| Control de autoridades |
|
|---|
 Datos: Q131267
Datos: Q131267- Multimedia: Sinornithosaurus / Q131267
- Especies: Sinornithosaurus