Sistema somatosensorial
El sistema somatosensorial comprende un complejo del organismo que consiste en centros de recepción y proceso, cuya función es producir modalidades de estímulo tales como el tacto, la temperatura, la propiocepción (posición del cuerpo) y la nocicepción (nociceptores: informan del dolor). Los receptores sensoriales actúan en la piel, el epitelio, el músculo esquelético, los huesos y articulaciones, órganos internos. Mientras que el tacto es considerado uno de los cinco sentidos tradicionales, la impresión del tacto está formada por varias modalidades. En medicina, el término coloquial "tacto" es usualmente reemplazado por "sentidos somáticos", con el objeto de reflejar en forma apropiada la variedad de mecanismos involucrados.

El sistema reacciona a los estímulos usando diferentes receptores: termorreceptores, mecanorreceptores y quimiorreceptores. La transmisión de información desde los receptores pasa por vía de los nervios sensoriales a través de tractos en la médula espinal y en el cerebro.
El proceso tiene lugar principalmente en el área somatosensorial primaria ubicada en el giro poscentral del lóbulo parietal de la corteza cerebral.
En resumen, el sistema se activa cuando una neurona sensorial es impulsada por algún estímulo específico, como el calor; esta neurona pasa a un área del cerebro especialmente atribuida al área del cuerpo que ha recibido el estímulo.
Anatomía
El sistema somatosensorial se extiende a través de todas las regiones importantes del cuerpo del mamífero (y otros vertebrados). Se compone de receptores sensoriales y neuronas sensoriales (aferentes) en la periferia (piel, músculo y órganos), hasta neuronas más profundas dentro del sistema nervioso central.
Vía somatosensorial
Un recorrido somatosensorial por lo general consta de tres neuronas sensitivas: primaria, secundaria y terciaria.
- La neurona sensorial primaria tiene su soma en el ganglio espinal del nervio espinal (si el estímulo está en la cabeza o cuello, se trata del ganglio de Gasser o ganglios nerviosos situados en otros nervios craneales).
- La neurona sensorial secundaria tiene su soma en la médula espinal y en el tronco del encéfalo. El axón ascendente atraviesa en forma de decusación hacia el lado opuesto en la médula espinal o el tronco del encéfalo. Los axones de muchas de estas neuronas terminan en el tálamo (p.ej. el núcleo ventral posterior), otros terminan en el sistema de activación reticular o en el cerebelo.
- En el caso del tacto y ciertos tipos de dolor, la neurona sensorial terciaria tiene su soma en el núcleo ventral posterior del tálamo y finaliza en el giro poscentral situado en el lóbulo parietal.
Periferia
En la periferia, el sistema somatosensorial detecta varios estímulos por medio de los receptores sensoriales, por ejemplo, los mecanorreceptores que detectan la sensación de tacto y los nociceptores que detectan el dolor. La información sensorial (tacto, dolor, temperatura, etc.) es entonces transmitida al sistema nervioso central por medio de las neuronas aferentes. Existen diferentes tipos de éstas, los cuales varían en tamaño, estructura y propiedades. Generalmente existe una correlación entre el tipo de modalidad sensorial detectada y el tipo de neurona aferente involucrada. Por ejemplo, neuronas lentas, delgadas, no mielinizadas conducen el dolor, en tanto que neuronas veloces, gruesas y mielinizadas conducen el tacto superficial.
Médula espinal
En la médula espinal, el sistema somatosensorial incluye conductos ascendentes desde el cuerpo hacia el cerebro. El giro poscentral en la corteza cerebral constituye un destino primordial como receptor sensorial. Hacia él se proyectan las neuronas de la vía dorsal-lemniscal medial y la vía espinotalámica. Adviértase que numerosas vías somatosensoriales incluyen sinapsis tanto en el tálamo como en la formación reticular antes de llegar a la corteza. Otras vías ascendentes, particularmente aquellas involucradas en el control de la postura son proyectadas hacia el cerebelo. Estas incluyen los tractos espinocerebelosos ventral y dorsal. Otro destino importante de neuronas somatosensoriales aferentes que entran en la médula espinal son aquellas neuronas involucradas en los reflejos segmentales.
Cerebro
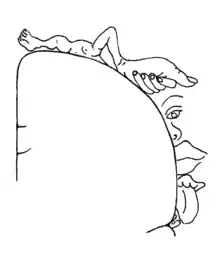
El área somatosensorial primaria en la corteza humana se localiza en el giro poscentral del lóbulo parietal. Aquella constituye el área de recepción sensorial principal del tacto. Al igual que otras áreas sensoriales, existe un mapa que refleja el espacio sensorial de la zona, mediante un homúnculo. Para la corteza somatosensorial primaria, podemos hablar del homúnculo sensorial. Áreas de esta parte del cerebro humano representan ciertas regiones del cuerpo, dependiendo del nivel de importancia de entrada somatosensorial. Por ejemplo, existe una gran región de la corteza dedicada a la sensación en las manos, mientras que a la espalda corresponde un área considerablemente menor (ver imagen). Como dato interesante, un estudio reveló que la corteza somatosensorial resultó ser un 21% más gruesa en 24 individuos que padecían migraña, contra un 12% en individuos de control, aunque aún se desconoce el significado de tal diferencia. Asimismo, la información somatosensorial involucrada en la propiocepción y la postura se proyecta hacia una parte enteramente diferente del cerebro, el cerebelo.
Fisiología

El proceso de «somatosensación» se inicia con la activación de un «receptor» físico. Estos receptores somatosensoriales yacen en la piel, órganos y músculos. La estructura de estos receptores es muy similar en todos los casos, consistiendo en terminaciones nerviosas libres o almacenadas en cápsulas especiales. Pueden ser activadas por movimiento (mecanorreceptor), presión (mecanorreceptor), estímulos químicos (quimiorreceptor), y/o temperatura. La activación también surge por medio de vibraciones generadas por roces, p.ej un dedo recorriendo una superficie. De esta forma podemos sentir aquellas texturas delicadas en las cuales la escala espacial es menor a 200 µm. Tales vibraciones son del orden de 250Hz, lo cual constituye la sensibilidad de frecuencia óptima en los corpúsculos de Pacini. En cada caso, el principio general de activación es similar; el estímulo causa depolarización en el extremo del nervio y luego una acción potencial es iniciada. En tanto ésta (usualmente) viaja desde adentro hacia la médula.
Vía somatosensorial general
Toda la información aferente al tacto/vibración asciende a la médula espinal a través de la vía columna dorsal-lemnisco medial- vía gracilis (T7 y por debajo) o cuneatus (T6 y por encima). El cuneatus envía señales al núcleo coclear indirectamente a través de la sustancia gris espinal, esta información se utiliza para determinar si un sonido percibido es sólo ruido/irritación de las vellosidades. Todas las fibras se cruzan (de izquierda a derecha) en la médula.
Una vía somatosensorial suele tener tres neuronas:[1] de primer orden, de segundo orden y de tercer orden.[2]
- La neurona de primer orden es un tipo de neurona pseudounipolar y siempre tiene su cuerpo celular en el ganglio de la raíz dorsal del nervio espinal con un axón periférico que inerva los mecanorreceptores táctiles y un axón central que hace sinapsis en la neurona de segundo orden. Si la vía somatosensorial se encuentra en partes de la cabeza o del cuello no cubiertas por los nervios cervicales, la neurona de primer orden será el ganglios del nervio trigémino o los ganglios de otros nervios craneales sensoriales).
- La neurona de segundo orden tiene su cuerpo celular en la médula espinal o en el tronco encefálico. Los axones ascendentes de esta neurona se cruzarán (decusarán) hacia el lado opuesto, ya sea en la médula espinal o en el tronco encefálico.
- En el caso del tacto y ciertos tipos de dolor, la neurona de tercer orden tiene su cuerpo celular en el núcleo ventral posterior del tálamo y termina en el giro postcentral del lóbulo parietal en la corteza somatosensorial primaria (o S1).

Los fotorreceptores, similares a los que se encuentran en la retina del ojo, detectan la radiación ultravioleta potencialmente dañina (ultravioleta A específicamente), induciendo una mayor producción de melanina por parte de los melanocitos.[3] Así pues, el bronceado ofrece potencialmente a la piel una protección rápida frente a los daños en el ADN y las quemaduras solares causadas por la radiación ultravioleta (daños en el ADN causados por el ultravioleta B). Sin embargo, es discutible si esto ofrece protección, porque la cantidad de melanina liberada por este proceso es modesta en comparación con las cantidades liberadas en respuesta al daño del ADN causado por la radiación ultravioleta B.[3]
Retroalimentación táctil
La retroalimentación táctil de la propiocepción deriva de los propioceptores de la piel, los músculos y las articulaciones.[4]
Equilibrio
El receptor del sentido del equilibrio reside en el sistema vestibular del oído (para la orientación tridimensional de la cabeza y, por inferencia, del resto del cuerpo). El equilibrio también está mediado por el reflejo cinestésico alimentado por la propiocepción (que percibe la ubicación relativa del resto del cuerpo con respecto a la cabeza).[5] Además, la propiocepción estima la ubicación de los objetos que son percibidos por el sistema visual (que proporciona la confirmación del lugar de esos objetos en relación con el cuerpo), como entrada a los reflejos mecánicos del cuerpo.
Tacto fino y tacto grueso
El tacto fino (o tacto discriminativo) es una modalidad sensorial que permite a un sujeto sentir y localizar el tacto. La forma de tacto en la que la localización no es posible se conoce como tacto burdo. La vía columna posterior-lemnisco medial es la vía responsable del envío de la información del tacto fino a la corteza cerebral del cerebro.
El tacto crudo (o tacto no discriminativo) es una modalidad sensorial que permite al sujeto percibir que algo le ha tocado, sin poder localizar dónde ha sido tocado (a diferencia del "tacto fino"). Sus fibras se transportan por el tracto espinotalámico, a diferencia del tacto fino, que se transporta por la columna dorsal.[6] Como el tacto fino funciona normalmente en paralelo al tacto grosero, una persona será capaz de localizar el tacto hasta que las fibras que transportan el tacto fino (Vía columna posterior-lemnisco medial) hayan sido interrumpidas. Entonces el sujeto sentirá el tacto, pero será incapaz de identificar dónde le han tocado.
Procesamiento neuronal del tacto social
El córtex somatosensorial codifica la información sensorial entrante procedente de receptores de todo el cuerpo. El tacto afectivo es un tipo de información sensorial que provoca una reacción emocional y suele ser de naturaleza social, como un contacto físico humano. En realidad, este tipo de información se codifica de forma diferente a otras informaciones sensoriales. La intensidad del tacto afectivo se sigue codificando en el córtex somatosensorial primario y se procesa de forma similar a las emociones invocadas por la vista y el oído, como ejemplifica el aumento de adrenalina provocado por el tacto social de un ser querido, frente a la incapacidad física de tocar a alguien a quien no se quiere.
Por otra parte, la sensación de placer asociada al tacto afectivo activa el córtex cingulado anterior más que el córtex somatosensorial primario. Los datos de imágenes por resonancia magnética funcional (fMRI) muestran que el aumento de la señal de contraste del nivel de oxígeno en sangre (BOLD) en el córtex cingulado anterior, así como en el córtex prefrontal, está altamente correlacionado con las puntuaciones de agradabilidad de un toque afectivo. La estimulación magnética transcraneal (EMT) inhibitoria del córtex somatosensorial primario inhibe la percepción de la intensidad del tacto afectivo, pero no de su agrado. Por lo tanto, el S1 no está directamente implicado en el procesamiento del tacto afectivo socialmente placentero, pero sigue desempeñando un papel en la discriminación de la ubicación y la intensidad del tacto.[7]
La interacción táctil es importante en la mayoría de los animales. Normalmente, el contacto táctil entre dos animales se produce mediante caricias, lamidos o acicalamiento. Estos comportamientos son esenciales para la salud social del individuo, ya que en el hipotálamo inducen la liberación de oxitocina, una hormona que disminuye el estrés y la ansiedad y aumenta el vínculo social entre animales.[8].
Más concretamente, se ha observado la consistencia de la activación de neuronas de oxitocina en ratas acariciadas por humanos, especialmente en el núcleo paraventricular caudal.[9] Se descubrió que esta relación afiliativa inducida por el contacto táctil es común independientemente de la relación entre los dos individuos (madre-bebé, macho-hembra, humano-animal). También se ha descubierto que el nivel de liberación de oxitocina a través de este comportamiento se correlaciona con el transcurso temporal de la interacción social, ya que las caricias más prolongadas inducían una mayor liberación de la hormona.[10]
También se ha observado la importancia de la estimulación somatosensorial en animales sociales como los primates. El acicalamiento forma parte de la interacción social que los primates ejercen sobre sus congéneres. Esta interacción es necesaria entre los individuos para mantener la relación afiliativa dentro del grupo, evitar conflictos internos y aumentar el vínculo grupal.[11] Sin embargo, dicha interacción social requiere el reconocimiento de cada miembro del grupo. Por ello, se ha observado que el tamaño del neocórtex está positivamente correlacionado con el tamaño del grupo, lo que refleja un límite en el número de miembros reconocibles entre los que puede producirse el acicalamiento.[11] Además, el curso temporal del acicalamiento está relacionado con la vulnerabilidad debida a la depredación a la que están expuestos los animales mientras realizan dicha interacción social. La relación entre la interacción táctil, la reducción del estrés y el vínculo social depende de la evaluación de los riesgos que se producen durante la realización de tales comportamientos en la vida salvaje, y se requieren más investigaciones para desvelar la conexión entre el cuidado táctil y el nivel de aptitud.
Variación individual
Diversos estudios han medido e investigado las causas de las diferencias entre individuos en el sentido del tacto fino. Un área bien estudiada es la agudeza espacial táctil pasiva, la capacidad de resolver los detalles espaciales finos de un objeto presionado contra la piel inmóvil. Se han utilizado diversos métodos para medir la agudeza espacial táctil pasiva, siendo quizás el más riguroso la tarea de orientación con rejilla.[12] En esta tarea los sujetos identifican la orientación de una superficie estriada presentada en dos orientaciones diferentes,[13] que puede aplicarse manualmente o con equipos automatizados.[14] Muchos estudios han demostrado una disminución de la agudeza espacial táctil pasiva con la edad;[15][16][17] se desconocen las razones de este declive, pero puede incluir la pérdida de receptores táctiles durante el envejecimiento normal. Sorprendentemente, la agudeza espacial táctil pasiva del dedo índice es mejor entre los adultos con dedos índices más pequeños;[18] se ha demostrado que este efecto del tamaño de los dedos subyace a la mejor agudeza espacial táctil pasiva de las mujeres, por término medio, en comparación con los hombres.[18] La densidad de corpúsculos táctiles, un tipo de mecanorreceptor que detecta vibraciones de baja frecuencia, es mayor en los dedos más pequeños;[19] lo mismo puede ocurrir con las células de Merkel, que detectan las hendiduras estáticas importantes para la agudeza espacial fina.[18] Entre los niños de la misma edad, los que tienen dedos más pequeños también tienden a tener mejor agudeza táctil.[20] Muchos estudios han demostrado que la agudeza espacial táctil pasiva es mayor en las personas ciegas que en las videntes de la misma edad,[17][21][22][23][24] posiblemente debido a la plasticidad modal cruzada en la corteza cerebral de los individuos ciegos. Tal vez también debido a la plasticidad cortical, los individuos que han sido ciegos desde el nacimiento, según se informa, consolidan la información táctil más rápidamente que las personas videntes.[25]
Tecnología
La nueva área de investigación denominada tecnología háptica puede proporcionar sensación de tacto en entornos reales y virtuales. Esta nueva disciplina ha comenzado a proveer ideas cruciales en lo que concierne a capacidad de tacto.
Sociedad y cultura
La tecnología háptica puede proporcionar sensación táctil en entornos virtuales y reales.[26] En el campo de la logopedia, la retroalimentación táctil puede utilizarse para tratar trastornos del habla.
El tacto afectuoso está presente en la vida cotidiana y puede adoptar múltiples formas. Estas acciones, sin embargo, parecen conllevar funciones específicas aunque no se comprenda del todo el beneficio evolutivo de una gama tan amplia de comportamientos. Los investigadores estudiaron los patrones de expresión y las características de 8 acciones de tacto afectuoso diferentes (abrazar, coger, besar, inclinarse, acariciar, apretar, acariciar y hacer cosquillas) en un estudio de autoinforme.[27] Se encontró que el toque afectuoso tiene distintas áreas objetivo en el cuerpo, diferente afecto asociado, valor de confort y frecuencia de expresión basada en el tipo de acción de toque que se realiza.
Además de las consecuencias sensoriales bastante obvias del tacto, también puede afectar a aspectos cognitivos de mayor nivel, como los juicios sociales y la toma de decisiones. Este efecto podría deberse a un proceso de andamiaje físico-mental en el desarrollo temprano, en el que las experiencias sensoriomotoras están vinculadas a la aparición del conocimiento conceptual.[28] Estos vínculos pueden mantenerse durante toda la vida, de modo que tocar un objeto puede indicar la sensación física de su procesamiento conceptual relacionado. De hecho, se ha descubierto que diferentes propiedades físicas -peso, textura y dureza- de un objeto tocado pueden influir en el juicio social y la toma de decisiones.[29] Por ejemplo, los participantes describieron un pasaje de una interacción social como más duro cuando tocaban un bloque de madera duro en lugar de una manta blanda antes de la tarea. A partir de estos hallazgos, la capacidad del tacto para influir inconscientemente en estos pensamientos de orden superior puede proporcionar una herramienta novedosa para las estrategias de marketing y comunicación.
Referencias
- Saladin KS. Anatomía y Fisiología 3ª edd. 2004. McGraw-Hill, Nueva York.
- 1007/978-3-642-28753-4_201964 Neurona de segundo orden. «Enciclopedia del dolor». Enciclopedia del dolor (Springer). 2013. p. 3448. ISBN 978-3-642-28752-7. doi:10.1007/978-3-642-28753-4_201964. Consultado el 2 de diciembre de 2022.
- Zukerman, Wendy. newscientist.com/article/dn21127-skin-sees-the-light-to-againt-sunshine.html#.VMFiPC6sXXE «La piel 've' la luz para protegerse de los rayos solares». newscientist.com. New Scientist. Consultado el 22 de enero de 2015.
- Proske U, Gandevia SC (October 2012). «Los sentidos propioceptivos: sus funciones en la señalización de la forma del cuerpo, la posición y el movimiento corporal, y la fuerza muscular». Physiological Reviews 92 (4): 1651-97. PMID 23073629. doi:10.1152/physrev.00048.2011.
- Proske U, Gandevia SC (September 2009). «The kinaesthetic senses». The Journal of Physiology 587 (Pt 17): 4139-46. PMC 2754351. PMID 19581378. doi:10.1113/jphysiol.2009.175372.
- Case LK, Laubacher CM, Olausson H, Wang B, Spagnolo PA, Bushnell MC (May 2016). «Encoding of Touch Intensity But Not Pleasantness in Human Primary Somatosensory Cortex». The Journal of Neuroscience 36 (21): 5850-60. PMC 4879201. PMID 27225773. doi:10.1523/JNEUROSCI.1130-15.2016.
- Case LK, Laubacher CM, Olausson H, Wang B, Spagnolo PA, Bushnell MC (May 2016). «Encoding of Touch Intensity But Not Pleasantness in Human Primary Somatosensory Cortex». The Journal of Neuroscience 36 (21): 5850-60. PMC 4879201. PMID 27225773. doi:10.1523/JNEUROSCI.1130-15.2016.
- Knobloch, H. Sophie; Grinevich, Valery (2014). «Evolución de las vías de la oxitocina en el cerebro de los vertebrados». Frontiers in Behavioral Neuroscience 8: 31. ISSN 1662-5153. PMC 3924577. PMID 24592219. doi:10.3389/fnbeh.2014.00031.
- Okabe, Shota; Takayanagi, Yuki; Yoshida, Masahide; Onaka, Tatsushi (2020-06- 04). «Los estímulos de caricias suaves inducen la respuesta afiliativa hacia los humanos en ratas macho». Scientific Reports 10 (1): 9135. Bibcode:2020NatSR..10.9135O. ISSN 2045-2322. PMC 7272613. PMID 32499488. doi:10.1038/s41598-020-66078-7.
- Tang, Yan; Benusiglio, Diego; Lefevre, Arthur; Hilfiger, Louis; Althammer, Ferdinand; Bludau, Anna; Hagiwara, Daisuke; Baudon, Angel; Darbon, Pascal; Schimmer, Jonas; Kirchner, Matthew K.; Roy, Ranjan K.; Wang, Shiyi; Eliava, Marina; Wagner, Shlomo (Septiembre 2020). ncbi.nlm.nih.gov/32719563/ «El tacto social promueve la comunicación interfemenina a través de la activación de las neuronas parvocelulares de oxitocina». Nature Neuroscience 23 (9): 1125-1137. ISSN 1546-1726. PMID 32719563. S2CID 220810651. doi:10.1038/s41593-020-0674-y.
- Lehmann, J.; Korstjens, A. H.; Dunbar, R. I. M. (1 de diciembre de 2007). «Tamaño del grupo, acicalamiento y cohesión social en primates». Animal Behaviour 74 (6): 1617-1629. ISSN 0003-3472. S2CID 14866172. doi:10.1016/j.anbehav.2006.10.025.
- Van Boven, R. W.; Johnson, K. O. (1 de diciembre de 1994). «El límite de la resolución espacial táctil en humanos: Discriminación de la orientación de la rejilla en el labio, la lengua y el dedo». Neurology 44 (12): 2361-6. PMID 7991127. S2CID 32255147. doi:10.1212/WNL.44.12.2361.
- Craig JC (1999). «Grating orientation as a measure of tactile spatial acuity». Somatosensory & Motor Research 16 (3): 197-206. PMID 10527368. doi:10.1080/08990229970456.
- Goldreich D, Wong M, Peters RM, Kanics IM (June 2009). «A Tactile Automated Passive-Finger Stimulator (TAPS)». Journal of Visualized Experiments (28). PMC 2726582. PMID 19578327. doi:10.3791/1374.
- Stevens JC, Alvarez-Reeves M, Dipietro L, Mack GW, Green BG (2003). «Disminución de la agudeza táctil en el envejecimiento: a study of body site, blood flow, and lifetime habits of smoking and physical activity». Somatosensory & Motor Research 20 (3-4): 271-9. PMID 14675966. S2CID 19729552. doi:10.1080/08990220310001622997.
- Manning H, Tremblay F (2006). «Age differences in tactile pattern recognition at the fingertip». Somatosensory & Motor Research 23 (3-4): 147-55. PMID 17178550. S2CID 24407285. doi:10.1080/08990220601093460.
- Goldreich D, Kanics IM (April 2003). «La agudeza táctil aumenta en la ceguera». The Journal of Neuroscience 23 (8): 3439-45. PMC 6742312. PMID 12716952. doi:10.1523/jneurosci.23-08-03439.2003.
- Peters RM, Hackeman E, Goldreich D (December 2009). «Diminutive digits discern delicate details: El tamaño de la yema del dedo y la diferencia de sexo en la agudeza espacial táctil». The Journal of Neuroscience 29 (50): 15756-61. PMC 3849661. PMID 20016091. doi:10.1523/JNEUROSCI.3684-09.2009.
- Dillon YK, Haynes J, Henneberg M (November 2001). «La relación del número de corpúsculos de Meissner con los caracteres dermatoglíficos y el tamaño de los dedos». Journal of Anatomy 199 (Pt 5): 577-84. PMC 1468368. PMID 11760888. doi:10.1046/j.1469-7580.2001.19950577.x.
- Peters RM, Goldreich D (2013). «Agudeza espacial táctil en la infancia: efectos de la edad y el tamaño de la yema del dedo». PLOS ONE 8 (12): e84650. Bibcode:2013PLoSO...884650P. PMC 3891499. PMID 24454612. doi:10.1371/journal.pone.0084650.
- Stevens, Joseph C.; Foulke, Emerson; Patterson, Matthew Q. (1996). «Agudeza táctil, envejecimiento y lectura braille en la ceguera de larga duración.». Journal of Experimental Psychology: Applied 2 (2): 91-106. doi:10.1037/1076-898X.2.2.91.
- Van Boven RW, Hamilton RH, Kauffman T, Keenan JP, Pascual-Leone A (June 2000). «Tactile spatial resolution in blind braille readers». Neurology 54 (12): 2230-6. PMID 10881245. S2CID 12053536. doi:10.1212/wnl.54.12.2230.
- Goldreich D, Kanics IM (November 2006). «Performance of blind and sighted humans on a tactile grating detection task». Perception & Psychophysics 68 (8): 1363-71. PMID 17378422. doi:10.3758/bf03193735.
- Wong M, Gnanakumaran V, Goldreich D (May 2011). «Mejora de la agudeza espacial táctil en la ceguera: evidencia de mecanismos dependientes de la experiencia». The Journal of Neuroscience 31 (19): 7028-37. PMC 6703211. PMID 21562264. doi:10.1523/JNEUROSCI.6461-10.2011.
- Bhattacharjee A, Ye AJ, Lisak JA, Vargas MG, Goldreich D (October 2010). «Experimentos de enmascaramiento vibrotáctil revelan un procesamiento somatosensorial acelerado en lectores de braille congénitamente ciegos». The Journal of Neuroscience 30 (43): 14288-98. PMC 3449316. PMID 20980584. doi:10.1523/JNEUROSCI.1447-10.2010.
- Gabriel Robles-De-La-Torre. «Sociedad Internacional de Háptica: Tecnología háptica, una explicación animada». Isfh.org. Archivado desde el original el 7 de marzo de 2010. Consultado el 26 de febrero de 2010.
- Schirmer, Annett; Chiu, Man Hey; Croy, Ilona (September 2021). «Más de un tipo: Diferentes firmas sensoriales y funciones dividen el toque afectuoso». Emotion 21 (6): 1268-1280. ISSN 1931-1516. PMID 34435843. doi:10.1037/emo0000966.
- Williams, Lawrence E.; Huang, Julie Y.; Bargh, John A. (1 de diciembre de 2009). «La mente andamiada: los procesos mentales superiores se basan en la experiencia temprana del mundo físico». European Journal of Social Psychology 39 (7): 1257-1267. doi:10.1002/ejsp.665.
- Ackerman, Joshua M.; Nocera, Christopher C.; Bargh, John A. (25 de junio de 2010). «Las sensaciones hápticas incidentales influyen en los juicios y decisiones sociales». Science 328 (5986): 1712-1715. Bibcode:2010Sci...328.1712A. ISSN 1095-9203. PMC 3005631. PMID 20576894. doi:10.1126/science.1189993.
Bibliografía
- Croy, Ilona; Sehlstedt, Isac; Wasling, Helena Backlund; Ackerley, Rochelle; Olausson, Håkan (febrero de 2019). «Gentle touch perception: From early childhood to adolescence». Developmental Cognitive Neuroscience (en inglés) 35: 81-86. doi:10.1016/j.dcn.2017.07.009.
- Tiffany Field, Les bienfaits du toucher, Petite Bibliothèque Payot, 2006. (en francés)
- Ashley Montagu, La peau et le toucher, un premier langage, Seuil, 1979. (en francés)
- Jean Folan-Curran, M-J-T, FitzGerald, Neuro-anatomie clinique et neurosciences connexes, Maloine, Paris, 2003. (en francés)
- François Boureau, Jean-françois Vibert, Alain Sébille, Marie-Claude Lavallard-Rousseau, Neurophysiologie: De la physiologie à l'exploration fonctionnelle, Elsevier, Paris, 2e édition, 2011. (en francés)
- Leonard E. White, George J. Augustine, David Fitzpatrick, Cally Hall, Antony-Samuel Lamantia, James O. McNamara, Dale Purves, Neurosciences, De Boeck Université, Bruxelles, 4e édition, 2011. (en francés)
- Marc Crommelinck, Nicole Boisacq-Shepens, Neurosciences, Dunod, Paris, 4e édition, 2004. (en francés)
- Michael A. Paradiso, Barry W. Connors, Mark F. Bear, Neurosciences : À la découverte du cerveau, Éditions Pradel, Paris, 3e édition, 2007. (en francés)
- Mark R. Rosenzweig, S. Marc Breedlove, Neil V. Watson, Psychobiologie - De la biologie du neurone aux neurosciences comportementales, cognitives et cliniques, De Boeck Université, Bruxelles, 6e édition, 2012. (en francés)
Véase también
Enlaces externos
- Artículo en PDF sobre el sistema somatosensorial.
- Explorador cerebral - Atlas del encéfalo
| Control de autoridades |
|
|---|
 Datos: Q328835
Datos: Q328835 Multimedia: Somatosensation / Q328835
Multimedia: Somatosensation / Q328835