Transportador de monoamina
Los transportadores de monoaminas (MATs, del inglés: monoamine transporters) son estructuras proteicas que funcionan como transportadores integrales de la membrana plasmática para regular las concentraciones de neurotransmisores monoamina. Tres clases principales de MATs (SERT, DAT, NET) son responsables de la recaptación de sus neurotransmisores aminas correspondientes (serotonina, dopamina, norepinefrina). Los MATs están ubicados justo afuera de la hendidura sináptica (peri-sinápticamente), transportando el rebose de transmisores monoamina desde la hendidura sináptica devuelta hacia el citoplasma de la neurona presináptica. La regulación de las MAT ocurre generalmente a través de fosforilación y modificación postraduccional. Debido a su importancia en la señalización neuronal, los MATs son el objetivo de muchos fármacos terapéuticos asociado con trastornos del estado de ánimo, tales como fluoxetina (Prozac) y metilfenidato (Ritalin). Los compuestos sintéticos tales como la anfetamina y cocaína también pueden dirigirse hacia los MATs.
Estimulantes
Hay varios transportadores de monoaminas diferentes, cada uno perteneciente a la familia de transportadores de membrana neuronales de sustratos específicos Na+/Cl- dependientes.[1]
- El transportador de dopamina, DAT.
- El transportador de norepinefrina, NET.
- El transportador de serotonina, SERT.
Función
Transportador de dopamina (DAT)
El transportador de dopamina (DAT) es responsable de la recaptación de dopamina (DA) extracelular Na+/Cl- dependiente.[1] El DAT también puede transportar norepinefrina extracelular. Los DATs pueden ser encontrados en el sistema nervioso central (SNC), donde se localizan en la sustancia negra y en el área tegmental ventral (ATV). Los DATs también se encuentran en el sistema nervioso periférico (SNP) donde se localizan en el estómago, páncreas, y en linfocitos.[1] Varias quinasas han sido relacionadas con la regulación de DAT incluyendo PKA, PKC, PI3K, MAPK3, MAPK1, PKB, CaM quinasas II, CDK5, y MAPK.[1]
Transportador de norepinefrina (NET)
El NET es responsable de la recaptación de norepinefrina (NE) extracelular Na+/Cl- dependiente.[1] El NET también pueden recapturar DA extracelular. Dentro del SNC, los NETs están ubicados en las dendrita y axones ubicados ambos en el hipocampo y córtex. Periféricamente, los NETs pueden ser encontrados en las neuronas simpáticas periféricas, la médula adrenal, el pulmón, la placenta, y conducto deferente.[2][1] La regulación de NET ha sido vinculado a MAPKs, insulina, PKC, y angiotensina II.[1]
Transportador de serotonina (SERT)
El SERT es responsable de la recaptación de serotonina (5-HT) extracelular en un proceso Na+/Cl- dependiente.[1] En el SNC, el SERT se ubica en la corteza cerebral, las regiones CA1 y CA3 del hipocampo, como también en los núcleos del rafe mediano y dorsal. En el SNP, el SERT se ubica en el tracto intestinal, glándulas suprarrenales, placenta, pulmones, y plaquetas.[2][1] La expresión de SERT en las plaquetas es usada como un medio de readquirir 5-HT desde el medio extracelular posteriormente utilizado en la activación de plaquetas. La regulación de SERT ha sido vinculado a la depleción aguda de CaNa2+ intracelular, inhibición de calmodulina, CaMKII, Src (gen), p38 MAP quinasa, PKC, y activación de NOS/GMPc.[1]
Estructura y mecanismo
Los transportadores de monoaminas (MAT en inglés) son miembros del grupo de transportadores neuronales de membrana de sustrato específico Na+/Cl--dependiente pertenecientes a la familia de genes SLC6.[1] Los MAT son grandes proteínas integrales de membrana compuestas de 12 dominios transmembranales conectados por bucles intracelulares y extracelulares. Los terminales NH2 y COOH de las proteínas MAT se encuentran dentro del citoplasma de las células presinápticas. Todos los MAT contienen sitios para la fosforilación de proteína quinasa mediante la proteína quinasa AMPc-dependiente, la proteína quinasa C (PKC) y la proteína quinasa Ca2+/calmodulina-dependiente.[2]
Los MAT son responsables de la captación de monoaminas mediante la unión secuencial y co-transporte de iones Na+ y Cl-. El gradiente de concentración de iones generado por la membrana plasmática Na+/K+ ATPasa proporciona la fuerza impulsora para la captación de monoamina mediada por transportadores.[2][3]
En el caso de NET y SERT un ion de Na+ y uno de Cl- son transportados junto con un NE o un 5-HT respectivamente. Cuando los gradientes iónicos son alterados (el K+ extracelular aumenta o el Na+ o Cl- disminuye) estos transportadores pueden funcionar en reversa resultando en un flujo de salida neto de substratos e iones de una neurona.[2]
Para regresar a una conformación que mira hacia fuera el SERT requiere el transportador de K+ intracelular. No existe evidencia de que los otros transportadores tengan tal requerimiento.[2]
La fosforilación juega un papel clave en la función de los MATs. Cuando el SERT es fosforilado mediante la vía PKC-dependiente ocurre la internalización del SERT. La internalización del SERT reduce la captación de 5-HT.[1] Eventos similares de fosforilación ocurren en DAT y NET, disminuyendo la capacidad de transporte de monoaminas de las células.
| MAT | Gen | Tamaño | Cromosoma Humano |
|---|---|---|---|
| DAT | hDAT | 620 aminoácidos | 5p15.3[1] |
| SERT | hSERT | 630 aminoácidos | 17q11.2[1] |
| NET | hNET | 617 aminoácidos | 16q12.2[1] |
Trastornos asociados y tratamientos
Se cree que los transportadores de monoaminas son factores en varias condiciones neurológicas debido a su rol en la recaptación de las monoaminas dopamina, noradrenalina, y 5-hidroxitriptamina. Estas condiciones incluyen TDAH, depresión, abuso de drogas, enfermedad de Parkinson, esquizofrenia, y síndrome de Tourette. La evidencia que apoya esta creencia incluye que los transportadores de monoaminas, DAT, NET, y SERT, son importantes sitios objetivo para fármacos terapéuticos utilizados en el tratamiento de trastornos del humor. Varios medicamentos se usan para tratar síntomas de enfermedades mediante el bloqueo de los transportadores de monoaminas, lo que resulta en un aumento de monoaminas extracelulares.[4] Además, se ha demostrado que los niveles de transportadores de monoamina son alterados en muchas de estas enfermedades psiquiátricas y neurológicas. Finalmente, las variaciones polimórficas en los genes de transportadores de monoaminas han sido propuestas de estar asociadas con condiciones tales como el TDAH y la depresión.[2]
Trastorno por déficit de atención con hiperactividad
Se ha observado que la hiperactividad, inatención e impulsividad en el TDAH está relacionado con anomalías en la función y regulación de DAT. La hipofunción dopaminérgica en la corteza frontal y ganglios basales es una característica neurobiológica observada en TDAH.[5] Psicoestimulantes, metilfenidato y anfetamina, que inhiben fuertemente el DAT, son eficaces en el tratamiento del TDAH. El metilfenidato (Ritalin) inhibe el DAT y el NET, lo que resulta en un aumento de la dopamina y norepinefrina extracelular que pueden fácilmente ligarse a las células postsinápticas. El metilfenidato actúa sobre el DAT como un inhibidor de la recaptación no selectivo.[1] El metilfenidato no es un inhibidor del SERT.[5]
Depresión
Se ha observado que la patología de la depresión implica la disfunción de los circuitos de neurotransmisores monoamina en el sistema nervioso central (SNC), particularmente de la serotonina y la norepinefrina. Los inhibidores selectivos de la recaptación de serotonina (ISRS) son los antidepresivos más utilizados e incluyen la fluoxetina (Prozac), citalopram (Celexa), y fluvoxamina. Estos fármacos inhiben la recaptación de la serotonina desde el espacio extracelular hacia el terminal sináptico al inhibir selectivamente el SERT. Se ha observado recientemente que la serotonina, la norepinefrina y la dopamina podrían estar involucrados en la depresión. Por lo tanto, fármacos tales como la venlafaxina y paroxetina están siendo utilizados como antidepresivos eficaces que inhiben selectivamente al SERT y al NET.[6] El antidepresivo tricíclico desipramina es un fármaco antidepresivo que es un inhibidor relativamente selectivo de la recaptación de NE. Estudios de la inhibición de NET correlacionan con la actividad antidepresiva.[7]
Esquizofrenia
La regulación de NET está vinculada a una transmisión de dopamina alterada y comportamientos similares a la esquizofrenia. La nisoxetina es un inhibidor de NET y revierte algunos comportamientos ligados a la esquizofrenia. Los NET regulan las actividades de la NE así como el equilibrio de la DA. Además, para el aclaramiento normal de DA es necesario un DAT funcional, lo que sugiere que la disfunción de DAT podría contribuir a la esquizofrenia.[1]
Psicoestimulantes
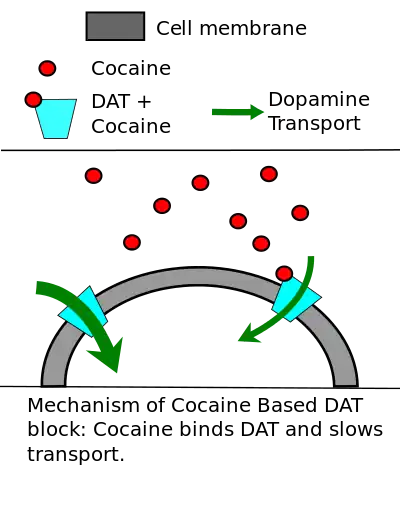
El DAT también es objetivo de varios "bloqueadores de DAT" incluyendo anfetaminas y la cocaína. Estos productos químicos inhiben la acción del DAT y, en menor medida, los otros transportadores de monoamina, pero sus efectos están mediados por mecanismos independientes.
Los transportadores de la monoamina son los objetivos de muchos agentes farmacológicos que afectan la función cerebral, incluyendo los psicoestimulantes cocaína y anfetaminas. La cocaína y anfetaminas emplean mecanismos diferentes que ambos resultan en un aumento en monoaminas extracelulares al disminuir la recaptación. Los psicoestimulantes afectan principalmente al DAT, aunque existe una cierta inhibición del SERT y NET. Se cree que los grandes aumentos en la dopamina sináptica resultando en el aumento de la estimulación de las neuronas objetivo crean las sensaciones reportadas por los usuarios de cocaína y anfetaminas.[2]
Cocaína
Los efectos estimulantes y eufóricos de la cocaína son creados cuando la cocaína inhibe la recaptación de la dopamina por DAT, lo que resulta en un aumento de la dopamina extracelular. La dopamina puede entonces unirse más fácilmente con las neuronas, lo que sobreestimula las células. La cocaína es un inhibidor competitivo no selectivo de los transportadores de monoamina, compartiendo un mecanismo similar al metilfenidato. La cocaína interactúa el DAT, SERT, y NET, aunque los efectos en el comportamiento y el refuerzo de la cocaína dependen de su inhibición del DAT y el aumento de la dopamina extracelular.[2]
Anfetaminas
Las anfetaminas funcionan como sustratos para los transportadores de monoaminas. La familia de las anfetaminas incluye una amplia variedad de subtipos incluyendo la dextroanfetamina, metanfetamina, metilendioximetanfetamina (éxtasis), y catinona.[4] Una vez transportadas al terminal sináptico de las neuronas, las anfetaminas son encerradas en vesículas sinápticas y actúan como bases débiles en las vesículas. Esto causa una redistribución de las monoaminas vesiculares desde la vesícula sináptica hacia el citoplasma del terminal sináptico.[2] Una elevación en la concentración de monoamina citoplasmática conduce a la reversión de los transportadores de monoaminas. Como resultado, existe una liberación masiva de neurotransmisores monoamina en el espacio extracelular. El exceso de neurotransmisores monoamina en la hendidura sináptica conduce a la sobreestimulación de las células.[4]
Historia de la investigación
El campo de la investigación de transportadores de monoaminas comenzó aproximadamente hace cinco décadas con la investigación sobre los NETs de Julius Axelrod. Axelrod finalmente recibió su Premio Nobel de esta investigación, que llevó al descubrimiento de los DATs y SERT, así como las consecuencias asociadas con las interacciones de los antidepresivos y psicoestimulantes con las proteínas MATs. Desde los estudios iniciales de Axelrod, la comprensión de las propiedades farmacológicas y funcionales de las proteínas MAT ha sido esencial en el descubrimiento de tratamientos terapéuticos de muchos trastornos mentales.[2] Durante la década de 1990 varias técnicas de clonación utilizando MATs han dilucidado la estructura genética de estas proteínas. En 1991, Susan Amara y sus colegas determinaron la secuencia de aminoácidos de los NETs, descubriendo sus similitudes de codificación relativamente altas a la del transportador de GABA.[2]
La investigación actual está en curso para entender cómo funcionan y cómo están regulados los MAT al ver los recién descubiertos dominios estructurales y funcionales de estas proteínas.[2][8] Durante la última década, la disponibilidad de alteración dirigida de genes transportadores de monoamina en modelos animales así como en los enfoques de formación de imágenes in vivo han mostrado progreso en los estudios asociados con trastornos psiquiátricos y de movimiento.[2] La investigación actual está tratando de aclarar el grado en que las cascadas de quinasas, proteínas que interactúan con los transportadores y la fosforilación contribuyen a la regulación de los MAT.[1]
Agentes MAT dobles y triples (IRSN y IRT)
Abajo hay ejemplos de fármacos que actúan directamente mediante la inhibición de dos o más MAT simultáneamente. Inhibidores de la recaptación de serotonina y noradrenalina IRSN actúan bloqueando ambos SERT y NET. Los inhibidores de la recaptación triple (IRT) actúan bloqueando los DAT, NET, y SERT simultáneamente. La mayoría de los antidepresivos modernos funcionan en el principio del bloqueo de transportadores de la recaptación. Los ISRS tales como la fluoxetina (Prozac) y IRSN como la venlafaxina son los principales tipos de fármacos administrados como primer tratamiento de la depresión y ansiedad.
- PRC200 es un ejemplo de un IRSN.
- JNJ-7925476 es un ejemplo de un TRI.
- Híbridos nocaína-modafinilo tal como 16e[9][10]
- Blough (2002) mostró que los MAT exhiben un "dominio remoto fenilo vinculante"[11]
Referencias
- Ramamoorthy, Sammanda, Toni Shippenberg, and Lankupalle Jayanthi. "Regulation of monoamine transporters: Role of transporter phosphorylation." Pharmacology & Therapeutics 129 (2010): 220-238. Print.
- Torres, Gonzalo E.; Raul R. Gainetdinov, Marc G. Caron (enero de 2003). «Plasma Membrane Monoamine Transporters: Structure, Regulation and Function». Nature Reviews Neuroscience 4 (1): 13-25. PMID 12511858. doi:10.1038/nrn1008.
- Gainetdinov, Raul, and Marc Caron. "Monoamine Transporters: From Genes to Behavior." The Annual Review of Pharmacology and Toxicology 43 (2003): 261-264. Print.
- H.H. Sitte; M. Freissmuth (2007). «17: Monoamine transporters in the brain: Structure and Function». En Abel Lajta, ed. Handbook of Neurochemistry and Molecular Neurobiology: Neural Membranes and Transport (3rd edición). Springer Reference. ISBN 978-0-387-30347-5.
- Fone, Kevin; David J Nutt (febrero de 2005). «Stimulants: use and abuse in the treatment of attention deficit hyperactivity disorder». Current Opinion in Pharmacology 5 (1): 87-93. PMID 15661631. doi:10.1016/j.coph.2004.10.001.
- Nemeroff, Charles B.; Michael J. Owens (octubre de 2002). «Treatment of mood disorders». Nature neuroscience: 1068-1070. doi:10.1038/nn943.
- Maarten E. A. Reith; Nian-Hang Chen (1997). Maarten E. A. Reith, ed. Neurotransmitter Transporters. Humana Press Inc. ISBN 0-89603-372-4.
- Hahn, MK; RD Blakely (marzo de 2002). «Monoamine Transporter Gene Structure and Polymorphisms in Relation to Psychiatric and Other Complex Disorders». The Pharmacogenomics Journal 2 (4): 217-235. PMID 12196911. doi:10.1038/sj.tpj.6500106.
- Zhou J, He R, Johnson KM, Ye Y, Kozikowski AP. Piperidine-based nocaine/modafinil hybrid ligands as highly potent monoamine transporter inhibitors: efficient drug discovery by rational lead hybridization. Journal of Medicinal Chemistry. 2004 Nov 18;47(24):5821-4. doi 10.1021/jm040117o PMID 15537337
- He R, Kurome T, Giberson KM, Johnson KM, Kozikowski AP (2005). «Further structure-activity relationship studies of piperidine-based monoamine transporter inhibitors: effects of piperidine ring stereochemistry on potency. Identification of norepinephrine transporter selective ligands and broad-spectrum transporter inhibitors». J. Med. Chem. 48 (25): 7970-9. PMID 16335921. doi:10.1021/jm050694s.
| Control de autoridades |
|
|---|
 Datos: Q5692421
Datos: Q5692421 Multimedia: Monoamine transporters / Q5692421
Multimedia: Monoamine transporters / Q5692421