Pectinidae
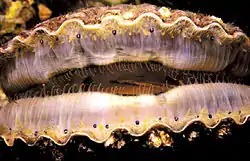
Los pectínidos (Pectinidae), conocidos por su nombre común como vieiras, son una familia de moluscos bivalvos, emparentados de cerca con las almejas y las ostras. La denominación taxonómica pectínido deriva del latín pecten, que significa «peina», «peineta». Los pectínidos tienen, en efecto, esa forma convexa típica de las peinas e, igualmente, diferencia de anchura entre la parte superior y la inferior. Ambas resultan ornamentales.
| Vieira | ||
|---|---|---|
| Rango temporal: Triásico a Reciente | ||
 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Mollusca | |
| Clase: | Bivalvia | |
| Orden: | Ostreoida | |
| Suborden: | Pectinina | |
| Superfamilia: | Pectinoidea | |
| Familia: |
Pectinidae Wilkes, 1810 | |
| Géneros | ||
| ||
El nombre de vieira se aplica principalmente a cualquiera de las numerosas especies de almejas de agua salada o moluscos bivalvos marinos de la familia taxonómica Pectinidae, si bien también se aplica a veces a especies de otras familias estrechamente relacionadas dentro de la superfamilia Pectinoidea, que también incluye a las ostras espinosas.
Las vieiras son una familia cosmopolita de bivalvos que se encuentran en todos los océanos del mundo, pero nunca en agua dulce. Son uno de los pocos grupos de bivalvos que son principalmente de "vida libre", en tanto muchas especies son capaces de nadar rápidamente distancias cortas e incluso de migrar cierta distancia por el fondo del océano. Una pequeña minoría de especies de vieiras vive adherida a sustratos rocosos cuando es adulta, mientras que otras se adhieren a objetos estacionarios o enraizados, como las praderas marinas, en algún momento de su vida mediante un filamento que segregan llamado biso. La mayoría de las especies, sin embargo, viven recostadas en sustratos arenosos y, cuando perciben la presencia de un depredador, como una estrella de mar, pueden intentar escapar nadando rápida pero erráticamente por el agua usando la propulsión a chorro que crean al chocar repetidamente sus conchas. Las vieiras tienen un sistema nervioso bastante desarrollado y, a diferencia de la mayoría de los bivalvos, todas las vieiras tienen un anillo de numerosos ojos simples situados alrededor del borde de sus mantos.
Muchas especies de vieiras son muy apreciadas como fuente de alimento, y algunas se crían para la acuicultura. La palabra "vieira" también se aplica a la carne de estos bivalvos, el músculo aductor, que se vende como marisco. Las conchas de las vieiras, de colores brillantes, simétricas y en forma de abanico, con su ornamentación radiada y a menudo estriada, son muy apreciadas por los coleccionistas de conchas y se han utilizado desde la antigüedad como motivos en el arte, la arquitectura y el diseño.
Debido a su amplia distribución, las conchas de vieira son habituales en las playas y a menudo tienen colores brillantes, lo que las convierte en un objeto popular para coleccionar entre veraneantes y personas que exploran las playas.[1] Las conchas también ocupan un lugar importante en la cultura popular, incluso en su simbolismo.
Nombres comunes

Se las suele llamar con la palabra gallega «vieira» porque en Galicia son abundantes. En castellano se llaman «veneras». Ambas palabras derivan de Venus, la diosa del amor. Botticelli y otros pintores la representan surgiendo de una «vieira». Como todos los bivalvos, la «vieira» o «venera» simboliza bien a la vulva.
Viven en aguas profundas de la mayor parte de los mares, sobre bancos de arena limpia y firme cerca de la costa, hasta 100 metros de profundidad. Son hermafroditas, y maduran primero las gónadas masculinas.
En Argentina, Colombia, España, Uruguay y Venezuela se denominan vieiras.
En Perú se conocen como conchas de abanico.
En México se les conoce como Pata de Mula.
En Chile se les llama ostiones y en sus aguas habitan dos especies: Argopecten purpuratus (ostión del norte), muy apreciado en la acuicultura del país, y Zigochlamys patagonica (ostión de sur o patagónico).[2]
Biología
Distribución y hábitat
Las vieiras habitan en todos los océanos del mundo, pero el mayor número de especies vive en la región del Indo-Pacífico. La mayoría de las especies viven en aguas relativamente poco profundas, desde la línea de bajamar hasta los 100 m, mientras que otras prefieren aguas mucho más profundas. Aunque algunas especies viven exclusivamente en ambientes muy reducidos, la mayoría son oportunistas y pueden vivir en una gran variedad de condiciones. Las vieiras pueden vivir dentro, sobre o debajo de rocas, coral, escombros, praderas marinas, kelp, arena o barro. La mayoría de las vieiras comienzan su vida como juveniles adheridos por medio del biso, capacidad que algunos conservan durante toda su vida, mientras que otros se convierten en adultos autónomos de vidas libres.[3]
Anatomía y fisiología
La valva es ondulada, y la inferior es más grande que la superior que es casi plana. Las ondulaciones radian desde el vértice de cada valva, efectuando un dibujo similar a un abanico. Sus bordes son afilados y ondulados.
Las vieiras poseen conchas con surcos, canales en los cuales se puede apreciar el registro del crecimiento del animal. Al igual que el resto de los moluscos bivalvos, sus valvas se abren y cierran gracias al músculo abductor. Son organismos nadadores, generalmente se encuentran reposando en el fondo sobre su valva inferior, la que suele tener una coloración más pálida que la superior.
La disposición interna de los órganos y sistemas dentro de la familia de las vieiras varía muy poco, por lo que lo que sigue puede aplicarse a la anatomía de cualquier especie de vieira..[3]
Orientación
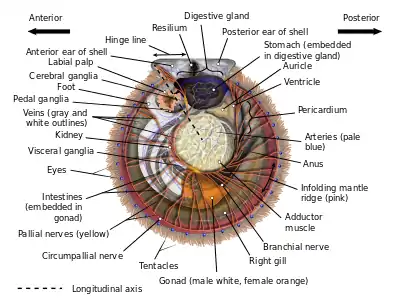
La concha de una vieira consta de dos lados or valvas, una izquierda y otra derecha, divididas por un plano de simetría.[4] La mayoría de las especies de vieiras se apoyan en su valva derecha y, por consiguiente, esta valva suele ser más profunda y redondeada que la izquierda (es decir, la superior), que en muchas especies es de hecho cóncava. En tanto la charnela (la articulación o "bisagra") de las dos valvas está orientada hacia arriba, un lado corresponde a la parte morfológica anterior o frontal del animal, el otro es la posterior o trasera, la charnela corresponde a la región dorsal o trasera/superior, y la parte inferior corresponde a la ventral o (por así decirlo) inferior/vientre.[5] Sin embargo, puesto que muchas conchas de vieira son más o menos simétricas bilateralmente ("equivalvadas"), así como simétricas al frente y atrás ("equiláteras"), determinar hacia qué lado está "orientado" un determinado animal requiere información detallada sobre sus valvas.
Valvas

La concha de una vieira típica consta de dos valvas de forma similar con una charnela en línea recta a lo largo de la parte superior, desprovista de dientes, y con un par de aletas planas u "orejas" (a veces llamadas "aurículas", aunque este es también el término para dos cámaras de su corazón) a cada lado de su punto medio, un rasgo que es único y evidente en todas las vieiras adultas.[3] Estas orejas pueden ser de tamaño y forma similares, si bien la anterior puede ser algo mayor (la posterior nunca es más grande que la anterior, una característica importante para distinguir cuál valva es). Como ocurre en casi todos los bivalvos, una serie de líneas y/o anillos de crecimiento se origina en el centro de la charnela, en un punto llamado "pico" rodeado por una zona generalmente elevada llamada "umbo". Estos anillos de crecimiento aumentan de tamaño de manera descendente hasta llegar al borde ventral curvo de la concha. Las conchas de la mayoría de las vieiras tienen una forma estilizada que facilita el movimiento durante la natación en algún momento de su ciclo vital, a la vez que proporcionan protección contra los depredadores. Las vieiras con valvas acanaladas tienen la ventaja de la fuerza arquitectónica que le proporcionan estas crestas llamadas "costillas", si bien éstas son algo costosas en términos de peso y masa. Una característica única de la familia de las vieiras es la presencia, en algún momento del ciclo de vida del animal, de una característica distintiva y taxonómicamente importante de la concha, una estructura en forma de peine llamada "ctenolium," situada en el borde anterior de la valva derecha, junto a la muesca bisal de la valva. Aunque muchas vieiras pierden esta característica al convertirse en adultos que nadan libremente, todas las vieiras tienen un ctenolium en algún momento de su vida, y ningún otro bivalvo tiene una característica análoga en su concha. El ctenolium sólo se encuentra en las vieiras modernas; los dos supuestos ancestros de las vieiras modernas, los Entoliidae y los Aviculopectinidae, no lo poseían.[6]
Sistema muscular
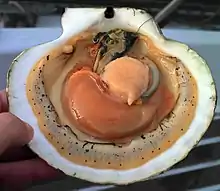
Al igual que las ostras verdaderas (familia Ostreidae),las vieiras tienen un único músculo aductor central, por lo que el interior de sus conchas presenta una característica cicatriz central que marca el punto de unión de este músculo. El músculo aductor de las vieiras es más grande y está más desarrollado que el de las ostras, debido a que las vieiras son nadadoras activas; se sabe que algunas especies de vieiras se desplazan en masa de una zona a otra. En las vieiras, la forma de la concha tiende a ser muy regular, y se suele utilizar como forma arquetípica de una concha marina.[5]
Ojos

Las vieiras tienen un gran número (hasta 200) de ojos pequeños (de aproximadamente 1 mm) dispuestos a lo largo del borde de sus mantos. Estos ojos representan una innovación particular entre los moluscos, ya que dependen de un espejo cóncavo y parabólico de cristales de guanina para enfocar y retroreflejar la luz en lugar de una lente como se encuentra en muchos otros tipos de ojos.[7] Además, sus ojos poseen una retina de doble capa: la exterior responde con mayor intensidad a la luz y la interior a la oscuridad abrupta.[8] Aunque estos ojos no son capaces de resolver formas con gran fidelidad, la sensibilidad combinada de ambas retinas a la luz que entra en el ojo y a la luz retroreflectada por el espejo otorga a las vieiras una definición de contraste excepcional, así como la capacidad de detectar patrones cambiantes de luz y movimiento.[9][10] Las vieiras confían principalmente en sus ojos como un sistema de detección de amenazas de "alerta temprana", escudriñando a su alrededor en busca de movimientos y sombras que podrían indicar la presencia de depredadores. Además, algunas vieiras modifican su comportamiento de natación o alimentación en función de la turbidez o claridad del agua, detectando el movimiento de material particulado en la columna de agua.[11]
Sistema digestivo
Las vieiras son filtradoras y se alimentan de plancton. A diferencia de muchos otros bivalvos, carecen de sifones. El agua se mueve sobre una estructura filtrante, donde las partículas de alimento quedan atrapadas en la mucosidad. A continuación, los cilios de la estructura mueven el alimento hacia la boca. Luego, el alimento se digiere en la glándula digestiva, un órgano al que a veces se denomina erróneamente "hígado", pero que envuelve parte del esófago, el intestino y todo el estómago. Los excrementos pasan por el intestino (cuya terminación, como la de muchos moluscos, entra y sale del corazón del animal) y salen por el ano.[5]: p.20 Como son organismos filtradores, ante fenómenos de afloraciones algales masivas (marea roja) es recomendable evitar su consumo.
Sistema nervioso
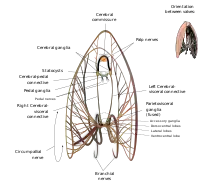
Como todos los bivalvos, las vieiras carecen de cerebro propiamente. En su lugar, su sistema nervioso está controlado por tres ganglios en pares, situados en varios puntos de su anatomía: los ganglios cerebrales o cerebropleurales, los ganglios pedales y los ganglios viscerales o parietoviscerales. Todos son de color amarillento. Los ganglios viscerales son, de lejos, los más grandes y extensos de los tres, y se presentan como una masa casi fusionada cerca del centro del animal – proporcionalmente, son los conjuntos de ganglios más grandes e intrincados de cualquier bivalvo moderno. De ellos irradian todos los nervios que conectan los ganglios viscerales con el anillo nervioso circumpalial que hace un bucle alrededor del manto y conecta con todos los tentáculos y ojos de la vieira. Este anillo nervioso está tan bien desarrollado que, en algunas especies, puede considerarse legítimamente como un ganglio adicional.[5]: p.46 Los ganglios viscerales son también el origen de los nervios branquiales que controlan las branquias de la vieira. Los ganglios cerebrales son el siguiente conjunto más grande de ganglios, y se encuentran separados a una distancia significativa, dorsales a los ganglios viscerales. Están unidos a los ganglios viscerales por largos conectores cerebro-viscerales, y entre sí por medio de una comisura cerebral que se extiende dorsalmente en un arco alrededor del esófago. Los ganglios cerebrales controlan la boca de la vieira a través de los nervios palpebrales, y también se conectan a los estatocistos que ayudan al animal a percibir su posición en el entorno. Están conectados a los ganglios pedales por conectivos cerebrales-pedales cortos. Los ganglios pedales, si bien no están fusionados, están situados muy cerca el uno del otro, cerca de la línea media. Desde los ganglios pedales, la vieira extiende nervios pedales que controlan el movimiento y la sensibilidad de su pequeño pie muscular.[5]: pp. 43–47
Reproducción
La familia de las vieiras es inusual en el sentido de que algunos miembros de la familia son dioicos (es decir, los machos y las hembras están separados), mientras que otros son hermafroditas simultáneos (ambos sexos en un mismo individuo), y unos pocos son hermafroditas protoandroicos (machos en la juventud que después pasan a ser hembras). La hueva de las hembras es roja, y la de los machos blanca. Los espermatozoides y los óvulos se liberan libremente en el agua durante la época de apareamiento, y los óvulos fecundados se hunden en el fondo. Al cabo de varias semanas, las vieiras inmaduras eclosionan y las larvas, versiones transparentes en miniatura de los adultos llamadas "freza", van a la deriva en el plancton hasta que se posan de nuevo en el fondo para crecer, normalmente fijándose mediante hilos de biso. Algunas vieiras, como la vieira de la bahía de Long Island (Argopecten irradians), tienen vidas cortas, mientras que otras pueden vivir 20 años o más. La edad puede inferirse a menudo a partir de los anillos concéntricos de sus conchas.[5]
Locomoción, movilidad y comportamiento

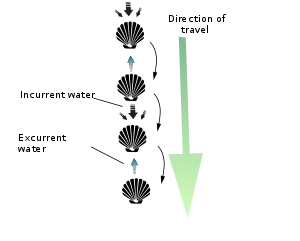
Las vieiras son en su mayoría de vida libre y activa, a diferencia de la gran mayoría de bivalvos, que son casi todos de movimiento lento y endofaunales. Se cree que todas las vieiras comienzan con un biso, que las adhiere a algún tipo de sustrato como la zostera marina cuando son muy jóvenes. La mayoría de las especies pierden el biso a medida que crecen. Otras especies de vieiras pueden encontrarse en el fondo del océano sujetas a objetos o sustratos duros por medio de tales hilos bisales (por ejemplo, Chlamys distorta o Hinnites multirigosus).[3] Los hilos bisales son fibras fuertes y sedosas que se extienden desde el pie muscular y que sirven para sujetarse a un soporte firme, como una roca. Algunas especies también pueden encontrarse en el fondo del océano, desplazándose con el uso de un pie extensible situado entre sus válvulas o enterrándose en la arena extendiendo y retrayendo sus pies.[5]
Sin embargo, la mayoría de las especies de la familia de las vieiras son nadadores activos de vida libre, que se impulsan a través del agua mediante el uso de los músculos aductores para abrir y cerrar rápidamente sus conchas. Pueden nadar con breves ráfagas de velocidad para escapar de depredadores (principalmente estrellas de mar). El cierre de las válvulas impulsa el agua con gran fuerza cerca de la charnela a través del velum, un pliegue en forma de cortina del manto que dirige la expulsión del agua alrededor de la charnela. Las vieiras nadan en la dirección de la apertura de la válvula, a menos que el velum dirija un cambio brusco de dirección.
En efecto, todos los aspectos relacionados con la forma característica de su concha -su simetría, estrechez, su superficie lisa y/o acanalada, su charnela pequeña y flexible, su potente músculo aductor y su borde continuo y uniformemente curvado- facilita tal actividad. A menudo lo hacen en ráfagas de varios segundos de duración antes de cerrar la concha por completo y de hundirse de nuevo en el fondo de su entorno. Las vieiras son capaces de desplazarse por una columna de agua, bien hacia delante/ventralmente (lo que se denomina "natación") aspirando agua a través del espacio entre sus valvas, una área llamada "boca" o "abertura", y expulsándola a través de unos pequeños orificios cerca de la línea de su charnela (su "bisagra") denominados aberturas exhalantes, o bien hacia atrás/dorsalmente (lo que se denomina "salto") expulsando el agua por de la misma manera en la que entró (es decir, ventralmente). Una vieira saltadora suele aterrizar en el fondo marino entre cada contracción de sus válvulas, mientras que una vieira nadadora se mantiene en la columna de agua durante la mayoría o todas sus contracciones y recorre una distancia mucho mayor (aunque rara vez a una altura superior a 1 m sobre el fondo marino y rara vez por distancias superiores a los 5 m).[3] Tanto los saltos como los movimientos natatorios consumen mucha energía, y la mayoría de las vieiras no son capaces de realizar más de cuatro o cinco seguidos antes de quedar completamente agotadas y necesitar varias horas de descanso. Si una vieira nadadora cae sobre su lado izquierdo, es capaz de darse la vuelta sobre su lado derecho mediante un movimiento similar al del palmeo de la concha llamado reflejo de enderezamiento. Se rumora que las llamadas vieiras cantoras emiten un suave sonido audible al agitar sus conchas bajo el agua (aunque es motivo de debate si esto realmente ocurre). Otras vieiras pueden extender su pie desde entre sus valvas y, contrayendo los músculos de este, pueden excavar en la arena.[12]
Las vieiras son muy sensibles a las sombras, las vibraciones, el movimiento del agua y los estímulos químicos.[13] Todas poseen una serie de 100 ojos azules, incrustados en el borde del manto de sus válvulas superiores e inferiores, que pueden distinguir entre la luz y la oscuridad. Sirven como mecanismo de defensa vital para evitar a los depredadores. Si bien son bastante débiles, su serie de ojos puede detectar movimientos circundantes y alertar ante la presencia de depredadores, más comúnmente estrellas de mar, cangrejos y caracoles.[5] Tanto la capacidad fisiológica como el nivel de ejercicio de las vieiras disminuyen con la edad debido al declive de la función celular y, especialmente, de la mitocondrial,[14] lo que aumenta su riesgo de captura y disminuye sus tasas de supervivencia. Los individuos de mayor edad muestran una menor densidad de volumen mitocondrial y capacidad aeróbica, así como una menor capacidad anaeróbica interpretada a partir de la cantidad de glucógeno almacenado en el tejido muscular.[14] Factores ambientales, como los cambios en los parámetros de estrés oxidativo, pueden inhibir el crecimiento y el desarrollo de las vieiras..[15]
Se ha demostrado que cambios estacionales en la temperatura y en la disponibilidad de alimento afectan a la capacidad metabólica muscular. Las propiedades de las mitocondrias del músculo aductor fásico de la especie Euvola ziczac varían significativamente durante su ciclo reproductivo anual. Las vieiras de verano en mayo tienen capacidades oxidativas máximas y de oxidación de sustratos más bajas que en cualquier otro momento del año. Este fenómeno se debe a los menores niveles de proteínas en los músculos aductores.[16]
Perlas

Las vieiras producen ocasionalmente perlas, aunque las perlas de vieira no muestran la acumulación de capas translúcidas o "nácar" que le dan su atractivo a las perlas de las ostras, y suelen carecer de lustre e iridiscencia. Pueden ser opacas, pequeñas y de color variable, pero hay excepciones que son apreciadas por sus cualidades estéticas.[17]
Mutualismo
Algunas vieiras, entre ellas Chlamys hastata, suelen llevar sobre sus conchas epibiontes como esponjas y percebes. La relación de la esponja con la vieira puede caracterizarse como una forma de mutualismo, en tanto la esponja proporciona protección al interferir con la adhesión de las patas tubulares de las estrellas de mar depredadoras,[18][19][20] camuflando a las vieiras Chlamys hastata de depredadores,[19] o formando una barrera física alrededor de las aberturas del biso para evitar que las estrellas de mar introduzcan sus membranas digestivas.[20] La incrustación de esponjas protege a las C. hastata del asentamiento de larvas de percebe, sirviendo de protección contra epibiontes que aumentan la susceptibilidad a depredadores. Por lo tanto, el asentamiento de larvas de percebe se produce con más frecuencia en conchas sin esponjas que en conchas con incrustaciones de esponjas.[18]
De hecho, la incrustación de percebes influye negativamente en la natación de C. hastata. Las vierias de C. hastata que nadan con incrustaciones de percebes requieren más energía y muestran una diferencia detectable en el gasto energético anaeróbico comparadas con aquellas que no tienen incrustación. En ausencia de incrustación de percebes, vieiras individuales nadan significativamente más tiempo, viajan más lejos y alcanzan una mayor elevación.[21]
Ciclo vital y crecimiento
Muchas vieiras son hermafroditas (tienen órganos femeninos y masculinos simultáneamente), alterando su sexo a lo largo de su vida, mientras que otras existen como especies dioicas, teniendo un sexo definido. En este caso, los machos se distinguen por tener testículos blancos con huevas y las hembras por tener ovarios anaranjados con huevas. Al alcanzar los dos años de edad suelen volverse sexualmente activos, pero no contribuyen de forma significativa a la producción de huevos hasta los cuatro años. El proceso de reproducción tiene lugar de forma externa mediante el desove, en el que los huevos y el esperma se liberan en el agua. El desove suele producirse a finales del verano y principios del otoño; también puede tener lugar en primavera en el ancón Atlántico Medio en la costa de los Estados Unidos.[22] Las hembras de las vieiras son muy fecundas, capaces de producir cientos de millones de huevos al año.[22]
Una vez fecundado el huevo, éste pasa a ser planctónico, es decir, un conjunto de microorganismos que flotan abundantemente en el agua dulce o salada. Las larvas permanecen en la columna de agua durante las siguientes cuatro a siete semanas antes de disiparse en el fondo del océano, donde se adhieren a objetos mediante hilos de biso. El biso se pierde eventualmente con la edad adulta, lo que convierte a casi todas las especies de vieiras en nadadoras libres. El crecimiento rápido se produce en los primeros años, con un aumento del 50-80 % en la altura de la concha y la cuadruplicación del tamaño en peso de la carne, y alcanzan el tamaño comercial a los cuatro o cinco años de edad aproximadamente.[22] Se sabe que la vida de algunas vieiras se prolonga más de 20 años.[23]
Etimología
El nombre de la familia Pectinidae, que se basa en el nombre del género tipo, Pecten, viene del latín pecten que significa peine, en referencia a una estructura en forma de peine de la concha que está situada junto a la muesca del biso.[24]
Filogenia
La historia fósil de las vieiras es rica en especies y especímenes. Los primeros registros conocidos de vieiras verdaderas (las que tienen un ctenolio) se encuentran en el período Triásico, hace más de 200 millones de años.[25] Las especies más antiguas se dividían en dos grupos, uno con un exterior casi liso: Pleuronectis von Schlotheim, 1820, mientras que el otro tenía costillas radiales o costilletas y aurículas: Praechlamys Allasinaz, 1972.[26] Los registros fósiles también indican que la abundancia de especies dentro de los Pectinidae ha variado mucho a lo largo del tiempo; los Pectinidae eran la familia de bivalvos más diversa en la era mesozoica, pero el grupo desapareció casi por completo a finales del Cretácico. Los supervivientes se especiaron rápidamente durante el periodo Terciario. Se han introducido casi 7.000 nombres de especies y subespecies tanto para los Pectinidae fósiles como para los recientes.[27]

El cladograma se basa en filogenía molecular usando marcadores de genes mitocondriales (12S, 16S) y nucleares (18S, 28S, and H3) hechos por Yaron Malkowsky y Annette Klussmann-Kolb en 2012.[28]
| Pteriomorphia |
| ||||||||||||||||||||||||||||||||||||||||||



Estructura taxonómica
Las vieiras pertenecen a la familia Pectinidae, moluscos bivalvos marinos de la superfamilia Pectinoidea. Otras familias dentro de esta misma superfamilia comparten una forma general de concha algo similar, y algunas especies dentro de algunas de las familias relacionadas también reciben comúnmente el nombre de "vieiras" (por ejemplo, Propeamussiidae, llamadas en inglés las vieiras del lodo).
La familia Pectinidae es la más diversificada de los pectinoideos en los océanos actuales. Pectinidae es una de las mayores familias de bivalvos marinos y contiene más de 300 especies existentes en 60 géneros. Su origen se remonta al Período Triásico Medio, hace aproximadamente 240 millones de años.[29] En términos de diversidad, ha sido una familia próspera hasta la actualidad.[30]
La evolución desde su origen ha dado lugar a un grupo exitoso y diverso: los pectínidos están presentes en todos los mares del mundo, encontrándose en ambientes que van desde la zona intermareal hasta las profundidades del hadal. Los pectínidos desempeñan un papel muy importante en muchas comunidades bentónicas y presentan una amplia gama de formas, tamaños, escultura y cultura de conchas.[31]
Raines y Poppe[33]enumeraron casi 900 nombres de especies de vieiras, pero la mayoría de ellos se consideran cuestionables o no válidos. Raines y Poppe mencionaron más de 50 géneros y unas 250 especies y subespecies. Aunque las especies suelen estar bien circunscritas, su atribución a subfamilias y géneros es a veces equívoca, y la información sobre la filogenia y las relaciones de las especies es mínima, entre otras cosas porque la mayoría de los trabajos se han basado únicamente en la morfología de los adultos.[34]
Los primeras y más completas descripciones taxonómicas de esta familia se basaron en caracteres morfológicos macroscópicos de las conchas adultas y representan esquemas de clasificación ampliamente divergentes.[35][36] Se logró cierto nivel de estabilidad taxonómica cuando los estudios de Waller en 1986, 1991 y 1993 concluyeron las relaciones evolutivas entre los taxones de pectínidos basándose en hipotéticas sinapomorfías morfológicas, algo que los anteriores sistemas de clasificación de Pectinidae no lograron. Creó tres subfamilias de Pectinidae: Camptonectinidae, Chlamydinae y Pectininae.[37] [38][39]
El marco de su filogenia muestra que los estados de hábitos de vida repetidos derivan de la convergencia y el paralelismo evolutivos.[40][41] Estudios han determinado que la familia Pectinidae es monofilética, desarrollándose a partir de un único ancestro común. Los ancestros directos de Pectinidae fueron bivalvos parecidos a las vieiras de la familia Entoliidae.[42] Los Entoliidae tenían aurículas y muescas bisales sólo en la juventud, pero no tenían un ctenolio, un arreglo en forma de peine a lo largo de los márgenes de la muesca bisal en los Pectinidae. El ctenolio es el rasgo que define a la familia moderna de Pectinidae y es una característica que ha evolucionado dentro del linaje.[43]
En un artículo de 2008, Puslednik y cols. identificaron una considerable convergencia de la morfología de la concha en un subconjunto de especies de Pectinidae planeadoras, lo que sugiere que la evolución morfológica iterativa puede ser más frecuente en la familia de lo que se creía.[44]
Se han realizado varios esfuerzos para emprender estudios filogenéticos. Solo tres han evaluado más de 10 especies[45][46][47] y apenas uno ha incluido múltiples grupos externos.[46] Casi todos los análisis moleculares previos de los Pectinidae han utilizado únicamente datos mitocondriales. Las filogenias basadas únicamente en datos de secuencias mitocondriales no siempre proporcionan una estimación precisa del árbol de especies. Pueden surgir factores complicados debido a la presencia de polimorfismos genéticos en las especies ancestrales y a la coalescencia de linajes resultante.[48][49]
En las filogenias moleculares de los Bivalvia, tanto los Spondylidae como los Propeamussiidae se han resuelto como hermanos de los Pectinidae.[46][50]
Lista de subfamilias and géneros
Familia Pectinidae
- Subfamilia Camptonectinae Habe, 1977[51]
- Delectopecten Stewart, 1920
- Ciclopecten Seguenza, 1877
- Lyropecten Conrad, 1862
- Pseudohinnites Dijkstra, 1989
- Subfamilia Hemipectinae Habe, 1977 (en disputa, a menudo en Chlamydinae: Chlamydini)
- Hemipecten Adams & Reeve, 1849
- Subfamilia Palliolinae Korbkov in Eberzin, 1960
- Tribu Palliolini Waller, 1993
- Palliolum Monterosato, 1884
- Lissochlamys Sacco, 1897
- Placopecten Verrill, 1897
- Pseudamussium Mörch, 1853
- Mesopeplum Iredale, 1929
- Tribu Palliolini Waller, 1993
- Subfamilia Pectininae
- Tribu Amusiini Ridewood, 1903
- Amusium Röding, 1798
- Dentamussium Dijkstra, 1990
- Euvola Dall, 1898
- Leopecten Masuda, 1971
- Ylistrum Mynhardt & Alejandrino, 2014
- Tribu Decatopectinini Waller, 1986
- Anguipecten Dall, Bartsch & Rehder, 1938
- Annachlamys Iredale, 1939
- Bractechlamys Iredale, 1939
- Decatopecten Rüppell en G. B. Sowerby II, 1839
- Excellichlamys Iredale, 1939
- Flexopecten Sacco, 1897
- Glorichlamys Dijkstra, 1991
- Gloripallium Iredale, 1939
- Juxtamusium Iredale, 1939
- Mirapecten Dall, Bartsch & Rehder, 1938
- Tribu Pectinini Wilkes, 1810
- Annachlamys Iredale, 1939
- †Gigantopecten Rovereto, 1899
- Minnivola Iredale, 1939
- †Oopecten Sacco, 1897
- †Oppenheimopecten Teppner, 1922
- Pecten Müller, 1776 (incluye a las vieiras reales o gigantes, Pecten maximus; Pecten jacobaeus; vieira japonesa, Pecten yessoensis; vieira neozelandesa, Pecten novaezealandiae; y la vieira Ravenel, Pecten raveneli)
- Serratovola Habe, 1951
- Tribu Amusiini Ridewood, 1903

- Subfamilia Chlamydinae von Teppner, 1922
- Tribu Clamydini von Teppner, 1922
- Chlamys Röding, 1798
- Complichlamys Iredale, 1939
- Coralichlamys Iredale, 1939
- Equichlamys Iredale, 1929
- Hinnites Deference, 1821
- Laevichlamys Waller, 1993
- Manupecten Monterosato, 1872
- Nodipecten Dall, 1898
- Notochlamys Cotton, 1930
- Pascahinnites Dijkstra & Raines, 1999
- Pedum Bruguière, 1791
- Psychrochlamys Jonkers, 2003
- Scaeochlamys Iredale, 1929
- Semipallium Jousseaume in Lamy, 1928
- Swiftopecten Hertlein, 1936
- Veprichlamys Iredale, 1929
- Tribu Austrochlamydini Jonkers, 2003
- Austrochlamys Jonkers, 2003
- Tribu Adamussiini Habe, 1977
- Adamussium Thiele, 1934
- Tribu Fortipectinini Masuda, 1963
- Mizuhopecten Masuda, 1963
- Patinopecten Dall, 1898
- Tribu Crassadomini Waller, 1993
- Crassadoma Bernard, 1986
- Caribachlamys Waller, 1993
- Tribu Mimachlamydini Waller, 1993
- Mimachlamys Iredale, 1929
- Spathochlamys Waller, 1993
- Talochlamys Iredale, 1935 incluye a Talochlamys pusio (Linnaeus, 1758) == Chlamys distorta (da Costa, 1778)
- Tribu Aequipectinini F. Nordsieck, 1969
- Aequipecten Fischer, 1886 (incluye Aequipecten muscosus)
- Argopecten Monterosato, 1889 (incluye la vieira de bahía, Argopecten irradians, la vieira calico del Atlántico, Argopecten gibbus and y la vieira calico del Pacífico, Argopecten ventricosus)
- Cryptopecten Dall, Bartsch & Rehder, 1938
- Haumea Dall, Bartsch & Rehder, 1938
- Leptopecten Verrill, 1897
- Leptopecten latiauratus Conrad, 1837
- Volachlamys Iredale, 1939
- Tribu Clamydini von Teppner, 1922
- Subfamilia incertae sedis
- Hyalopecten Verrill, 1897

Usos
|
Pecten tigris Lamarck, 1819, especímenes de museo, Naturalis |
Las vieiras son bivalvos de gran importancia comercial, debido al sabroso sabor de su carne, que puede ser consumida cruda o cocida (en empanadas, a la parmesana, etc.). |
Simbolismo
 Símbolo oficial del Camino de Santiago |
La concha de la vieira común (Pecten jacobaeus) o venera recibió de Lineo esa denominación taxonómica en 1758, en obsequio al símbolo del peregrino en el Camino de Santiago (España). Curiosamente la denominación taxonómica Pecten jacobaeus puesta por Lineo se corresponde con la venera endémica del Mediterráneo y no con la venera de mucho mayor tamaño propia de las costas gallegas, que recibe el nombre de Pecten maximus. Al ser típica de las costas de Galicia, los peregrinos la llevaban a sus lugares de origen como prueba de que habían llegado hasta el final del camino. |
Notas
- Robinson y Robinson, 2000.
- «Sitio WORMS».
- Shumway y Parsons, 2011.
- Milsom y Rigby, 2009.
- Drew, 1906.
- Hautmann, Michael (2010). «The first scallop». Paläontologische Zeitschrift 84 (2): 317-322. S2CID 84457522. doi:10.1007/s12542-009-0041-5.
- Speiser, Daniel I.; Johnsen, Sönke (29 de diciembre de 2008). «Comparative Morphology of the Concave Mirror Eyes of Scallops (Pectinoidea)». American Malacological Bulletin 26 (1–2): 27-33. ISSN 0740-2783. S2CID 11584708. doi:10.4003/006.026.0204.
- Speiser, D. I.; Loew, E. R.; Johnsen, S. (2011). «Spectral sensitivity of the concave mirror eyes of scallops: potential influences of habitat, self-screening and longitudinal chromatic aberration». Journal of Experimental Biology 214 (3): 422-431. PMID 21228201. doi:10.1242/jeb.048108. Parámetro desconocido
|doi-access=ignorado (ayuda) - «Eyes detect changing movement patterns: queen scallop». asknature.org.
- Land, MF; Fernald, RD (1992). «The evolution of eyes». Annual Review of Neuroscience 15: 1-29. PMID 1575438. doi:10.1146/annurev.ne.15.030192.000245. Archivado desde el original el 9 de enero de 2018. Consultado el 30 de junio de 2021.
- Speiser, Daniel I.; Johnsen, Sönke (1 de julio de 2008). «Scallops visually respond to the size and speed of virtual particles». Journal of Experimental Biology 211 (13): 2066-2070. ISSN 0022-0949. PMID 18552295. doi:10.1242/jeb.017038. Parámetro desconocido
|doi-access=ignorado (ayuda) - Gosling, 2015.
- Land, M.F. (1966). «Activity in the optic nerve of Pecten maximus in response to changes in light intensity, and to pattern and movements in optical environment». Journal of Experimental Biology 45 (1): 83-99.
- Philipp, E.E.R.; Schmidt, M.; Gsottbauer, C.; Sänger, A. M.; Abele, D. (2008). «Size- and age- dependent changes in adductor muscle swimming physiology of the scallop Aequipecten opercularis». Journal of Experimental Biology 211 (15): 2492-2501. PMID 18626084. doi:10.1242/jeb.015966. Parámetro desconocido
|doi-access=ignorado (ayuda) - Guerra, C.; Zenteno-Savín, T.; Maeda-Martínez, A. N.; Abele, D.; Philipp, E. E. R. (2013). «The effect of predator exposure and reproduction on oxidative stress parameters in the Catarina scallop Argopecten ventricosus». Comparative Biochemistry and Physiology A 165 (1): 89-96. PMID 23416890. doi:10.1016/j.cbpa.2013.02.006.
- Boadas, M.A.; Nusetti, O.; Mundarain, F. (1997). «Seasonal variation in the properties of muscle mitochondria from the tropical scallop Euvola (Pecten) ziczac». Marine Biology 128 (2): 247-255. S2CID 84538863. doi:10.1007/s002270050089.
- Matlins, 2001.
- Bloom, S. (1975). «The motile escape response of a sessile prey: a sponge-scallop mutualism». Journal of Experimental Biology and Ecology 17 (3): 311-321. doi:10.1016/0022-0981(75)90006-4.
- Pitcher, C.R.; Butler, A.J. (1987). «Predation by asteroids, escape response, and morphometrics of scallops with epizoic sponges». Journal of Experimental Marine Biology and Ecology 112 (3): 233-249. doi:10.1016/0022-0981(87)90071-2.
- Forester, A.J. (1979). «The association between the sponge Halichondria panicea (Pallas) and scallop Chlamys varia (L.): a commensal protective mutualism». Journal of Experimental Marine Biology and Ecology 36 (1): 1-10. doi:10.1016/0022-0981(79)90096-0.
- Donovan, D.; Bingham, B.; Farren, H.; Gallardo, R.; Vigilant, V. (2002). «Effects of sponge encrustation on the swimming behaviour energetics and morphometry of the scallop Chlamys hastata». Journal of the Marine Biological Association of the United Kingdom 82 (3): 469-476. doi:10.1017/s0025315402005738.
- Hart, D.R.; Chute, A.S. (2004). «Essential Fish Habitat Source Document: Sea Scallop, Placopecten magellanicus, Life History and Habitat Characteristics». NOAA Tech Memo NMFS NE-189.
- «Scallop Aquaculture». College of Marine Science, University of South Florida.
- Rice, 2012.
- Hautmann, Michael (2010). «The first scallop». Paläontologische Zeitschrift 84 (2): 317-322. S2CID 84457522. doi:10.1007/s12542-009-0041-5.
- Waller, T. R. (1993). «The evolution of Chlamys (Mollusca: Bivalvia: Pectinidae) in the tropical western Atlantic and eastern Pacific». American Malacological Bulletin 10 (2): 195-249.
- Harper et al., 2000.
- Malkowsky, Yaron; Klussmann-Kolb, Annette (May 2012). «Phylogeny and spatio-temporal distribution of European Pectinidae (Mollusca: Bivalvia)». Systematics and Biodiversity 10 (2): 233-242. S2CID 84085349. doi:10.1080/14772000.2012.676572.
- Hautmann, Michael (2010). «The first scallop». Paläontologische Zeitschrift 84 (2): 317-322. S2CID 84457522. doi:10.1007/s12542-009-0041-5.
- Waller, Thomas R. (2006). «New phylogenies of the Pectinidae (Mollusca: Bivalvia): Reconciling morphological and molecular approaches». En Shumway, Sandra E., ed. Scallops: biology, ecology and aquaculture II (Amsterdam: Elsevier). pp. 1-44.
- Brand, A.R. (2006). «Scallop ecology: distributions and behavior». Scallops: Biology, Ecology and Aquaculture. Developments in Aquaculture and Fisheries Science 35. pp. 651-744. ISBN 9780444504821. doi:10.1016/S0167-9309(06)80039-6.
- Raines, Poppe y Groh, 2006.
- Raines, B. K. & Poppe, G. T. (2006): The Family Pectinidae.[32]
- Barucca, M., Olmo, E., Schiaparelli, S. & Canapa, A. (2004): Molecular phylogeny of the family Pectinidae (Mollusca: Bivalvia)
- Waller, Thomas R. (1972). «The functional significance of some shell micro-structures in the Pectinacea». Paleontology (International Geological Congress). pp. 48-56.
- Shumway, Sandra E.; Parsons, G. Jay, eds. (7 de junio de 2016). Scallops: Biology, Ecology, Aquaculture, and Fisheries. Elsevier Science. p. 5. ISBN 978-0-444-62719-3.
- Waller, Thomas R. (1986). «A new genus and species of scallop (Bivalvia: Pectinidae) from off Somalia, and the definition of a new tribe Decatopectinin i». Nautilus 100 (2): 39-46. doi:10.5962/bhl.part.26491. Parámetro desconocido
|doi-access=ignorado (ayuda) - Shumway y Waller, 1991.
- Waller, Thomas R. (1993). «The evolution of "Chlamys" (Mollusca: Bivalvia: Pectinidae) in the tropical western Atlantic and eastern Pacific». American Malacological Bulletin 10 (2): 195-249.
- Alejandrino, A.; Puslednik, L.; Serb, J. M. (2011). «Convergent and parallel evolution in life habit of the scallops». BMC Evolutionary Biology 11 (1): 164. PMC 3129317. PMID 21672233. doi:10.1186/1471-2148-11-164.
- Waller, Thomas R. (2007). «The evolutionary and biogeographic origins of the endemic Pectinidae (Mollusca: Bivalvia) of the Galápagos Islands». Journal of Paleontology 81 (5): 929-950. S2CID 86121432. doi:10.1666/pleo05-145.1.
- Dijkstra, H.H.; Maestrati, P. (2012). «Pectinoidea (Mollusca, Bivalvia, Propeamussiidae, Cyclochlamydidae n. fam., Entoliidae and Pectinidae) from the Vanuatu Archipelago». Zoosystema 34 (2): 389-408. S2CID 85935928. doi:10.5252/z2012n2a12.
- Waller, Thomas R. (1984). «The ctenolium of scallop shells: functional morphology and evolution of a key family-level character in the Pectinacea (Mollusca: Bivalvia)». Malacologia 25 (1): 203-219.
- Puslednik, L.; Serb, J.M. (2008). «Molecular phylogenetics of the Pectinidae (Mollusca: Bivalvia) and the effect of outgroupselection and increased taxon sampling on tree topology». Molecular Phylogenetics and Evolution 48 (3): 1178-1188. PMID 18579415. doi:10.1016/j.ympev.2008.05.006.
- Barucca, M.; Olmo, E.; Schiaparelli, S.; Capana, A. (2004). «Molecular phylogeny of the family Pectinidae (Mollusca: Bivalvia) based on mitochondrial 16S and 12S rRNA genes». Molecular Phylogenetics and Evolution 31 (1): 89-95. PMID 15019610. doi:10.1016/j.ympev.2003.07.003.
- Matsumoto, M.; Hayami, I. (2000). «Phylogenetic analysis of the family Pectinidae (Bivalvia) based on mitochondrial cytochrome C oxidase subunit». Journal of Molluscan Studies 66 (4): 477-488. doi:10.1093/mollus/66.4.477. Parámetro desconocido
|doi-access=ignorado (ayuda) - Saavedra, C.; Peña, J.B (2006). «Phylogenetics of American scallops (Bivalvia: Pectinidae) based on partial 16S and 12S ribosomal RNA gene sequences». Marine Biology 150 (1): 111-119. S2CID 84205954. doi:10.1007/s00227-006-0335-z.
- Pamilo, P.; Nei, M. (1988). «Relationships between gene trees and species trees». Molecular Biology and Evolution 5 (5): 568-583. PMID 3193878. doi:10.1093/oxfordjournals.molbev.a040517.
- Wu, C.I. (1991). «Inferences of species phylogeny in relation to segregation of ancient polymorphisms». Genetics 127 (2): 429-435. PMC 1204370. PMID 2004713.
- Waller, Thomas R., 1998. Origin of the Molluscan Class Bivalvia and a Phylogeny of Major Groups. Pp. 1–45. In: P.A. Johnston & J.W. Haggart (eds), Bivalves: An Eon of Evolution. Calgary: University of Calgary Press xiv + 461 pp.
- Habe, 1977.
Enlaces externos
| Control de autoridades |
|
|---|
 Datos: Q1311395
Datos: Q1311395 Multimedia: Pectinidae / Q1311395
Multimedia: Pectinidae / Q1311395 Especies: Pectinidae
Especies: Pectinidae