Évolution des stratégies de prédation
L’évolution des stratégies de prédation est un processus impliquant des changements comportementaux et structuraux au cours des générations dans la recherche et la capture des proies chez une population de prédateurs.
Ces traits issus de l'évolution sont héritables génétiquement ou culturellement (par apprentissage par les parents) au cours des générations. L'induction d'un nouveau trait dans une population de prédateurs implique que ce trait soit transmissible et que l'obtention de celui-ci confère aux individus une meilleure chance de survie (et donc qu'il se répande dans la population par sélection naturelle).
La prédation est une interaction entre un être vivant (appelé prédateur), qui en tue un autre (appelé proie) afin de s'en nourrir et d'ainsi maintenir son homéostasie. L'évolution a donné lieu à une multitude d'adaptations et à l'élaboration de stratégies qui constituent aujourd'hui la diversité des stratégies de prédation.
Les prédateurs et les stratégies de prédation qu'ils emploient ont un impact direct sur les défenses anti-prédation développées par les proies ainsi que sur la dynamique des populations de proies. On mesure couramment ces impacts sur les populations de proies à l'aide des modèles mathématiques de Lotka-Volterra ou de Nicholson-Bailey[1].
Le concept de stratégie de prédation

La quantité de nourriture qu’un prédateur peut s’approprier, digérer et convertir en énergie est limitée, soit par l’abondance des proies, par leur accessibilité ou encore par le temps nécessaire à leur capture et leur assimilation. Or, ces ressources limitées en nourriture et en énergie, l’organisme doit pouvoir les allouer à sa croissance, à son entretien, à la lutte contre ses ennemis et à sa reproduction.
Une stratégie de prédation doit donc pouvoir répondre aux questions suivantes :
- Quelles proies chercher ?
- Où et quand se nourrir ?
- Comment capturer de façon efficace la proie ? (Compromis entre effort de capture et valeur nutritionnelle de la proie).
Les prédateurs auront donc tendance à optimiser leurs stratégies afin de tirer de maximum de profit de la proie chassée. La sélection naturelle favorise donc, parmi les différentes stratégies possibles, celle qui confère à la population de prédateurs la meilleure fitness (valeur sélective) possible[2].
Cette solution, appelée stratégie optimale, dépendra des contraintes qui pèsent sur le prédateur telle que l’accessibilité de la proie, sa capturabilité ou encore les défenses anti-prédation mises en place par la proie (stratégies de fuite pour mieux échapper au prédateur, se rendre non comestible par la sécrétion de venins par exemple, capacité à se camoufler dans son environnement…).
Pour illustrer cette notion, on peut prendre l’exemple des crabes verts (Carcinus maenas), qui sont notamment des prédateurs des moules (Mytiloida). Une étude d’Elner et Hugues (1978) a montré que pour le crabe, le profit augmente avec la taille des moules puis diminue car les coquilles de grandes tailles sont plus longues à briser (et donc plus coûteuses en énergie). Cette étude a aussi montré que les crabes préfèrent consommer des moules de taille intermédiaire qui leur apporteront un maximum d’énergie pour un minimum d’effort[3].
Origine de la prédation
L’origine de la prédation est mal connue, elle remonte vraisemblablement à 2,7 milliards d’années par la prédation des procaryotes qui a amené l’évolution vers les cellules eucaryotes. L’apparition des prédateurs mobiles sur les bactéries et les protistes remonterait à 2 milliards d’années. À partir du Cambrien (il y a 550 millions d’années) il y a eu une transition entre des écosystèmes microbiens simples à des écosystèmes ayant un réseau trophique complexe (producteurs primaires, consommateurs)[4].
Réseaux trophiques
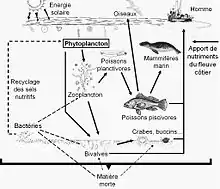
Les réseaux trophiques montrent à quel point les êtres vivants dépendent les uns des autres comme le sont le prédateur et sa proie. Au sein du réseau trophique, les prédateurs sont les consommateurs secondaires. Ce sont des animaux carnivores (zoophages) ayant pour proie les consommateurs primaires. De plus, dans certaines chaînes alimentaires, les super-prédateurs se situent au sommet de la chaîne alimentaire donc ils ne sont la proie d'aucun autre prédateur. Plus il y a d’inter-connectivité plus il y a une complexification du réseau trophique via le flux d’énergie à travers les différents niveaux trophiques[5].
Différentes stratégies de prédation
Il existe différents régimes alimentaires chez les prédateurs. En effet, certaines espèces de prédateurs peuvent être polyphages, c’est-à-dire ayant un régime très varié.
D’autres peuvent être oligophage c'est-à-dire qu’elles ne consomment qu’une faible diversité d’aliments. D’autres enfin peuvent être mono-phage, c’est-à-dire qu’ils se nourrissent exclusivement d’une seule espèce de proie.
Le régime alimentaire peut varier selon le stade de développement. En effet, certains êtres vivants sont herbivores ou omnivores à la naissance et deviennent des prédateurs à l’âge adulte (amphibiens). A contrario, d’autres sont prédateurs aux stades juvéniles et deviennent au cours de leur croissance granivores ou herbivores (perdrix). Le régime alimentaire peut aussi varier selon les saisons et les disponibilités nutritives dans le milieu ainsi que selon l’activité du métabolisme.
Il existe différentes stratégies de prédation.
Chasse passive
Lors de la chasse passive, les proies viennent aux prédateurs sans qu’il n'aient besoin d’agir. Dans certains cas, les proies sont attirées vers les prédateurs grâce aux adaptations de prédation. Par la suite, il suffit aux prédateurs de piéger et consommer les proies. Il s'agit donc d'une technique de chasse peu coûteuse en énergie.
Exemple : Les moules (filtreurs), les plantes carnivores...
Chasse à l'affût
Les prédateurs attendent patiemment leurs proies et les capturent grâce à des « armes » bien adaptées.
Exemple : le cas des araignées[6].
Certains mammifères chassent aussi à l’affût comme le tigre qui est un très mauvais sprinter mais qui est équipé de canines très performantes d’environ 9 cm de long qui lui permettent de saisir ses proies au niveau du cou afin de l’achever.
Chasse mobile solitaire
.jpg.webp)
Les prédateurs poursuivent leurs proies individuellement. Ainsi le butin n'est pas partagé, mais ils doivent redoubler d'efforts pour parvenir à capturer leurs proies.
Exemple: le guépard pratique une chasse solitaire et use de sa rapidité pour capturer sa proie.
Chasse en groupe

Ce sont des prédateurs sociaux qui pourchassent leurs proies en groupe en se partageant les tâches et le fruit de la chasse.
Exemple : Les orques qui chassent en gousse ou encore les loups en meute.
Les adaptations morphologiques à la prédation
Au cours de l’évolution, différents types d’adaptations ont été sélectionnées afin d’optimiser la prédation.
Différents organes et techniques de prédation ont donc été développés afin de permettre à un type de prédateur ayant une alimentation spécifique d’augmenter son taux de prédation :
- Dents : Certains prédateurs sont équipés de dents très acérées qui leur permettent de capturer, déchiqueter… Exemple : grands nombres de rangées pour les requins carnivores comme le grand requin blanc.
Il existe d’autres organes utiles à la prédation comme le bec chez les oiseaux qui est adapté au type de proie qu'il chasse ou encore d'autres appendices comme les pinces des homards…
- Dards urticants : Ce sont des tentacules composées de cellules urticantes qui peuvent paralyser, empoisonner voir tuer. On les retrouve chez certains cnidaires, comme la « guêpe des mers » (Chironex fleckeri) il s'agit de la méduse la plus venimeuse.
- Venin : Il existe des prédateurs qui possèdent un venin qui leur permet d'atteindre et de capturer leur proie. Exemple: Un grand nombre de serpents produisent du venin (le plus venimeux est le mamba noir, Dendroaspis polylepis).
- Électricité : Les prédateurs peuvent être équipés d'un organe électrique qui leur permet d’envoyer de fortes décharges électriques afin d'immobiliser ou de tuer leur proie. Exemple: L’anguille électrique (Electrophorus electricus) qui peut envoyer des décharges allant de 100 à 700 V.
.jpeg.webp)
- Leurre et piège : Certains prédateurs utilisent des techniques de piégeage ou de leurre pour attirer les proies jusqu'à eux. Exemple: la baudroie (Lophius piscatorius) est équipée d’un organe bioluminescent qui lui permet d’attirer ses proies dans les profondeurs des océans en imitant les signaux lumineux produits lors de la parade sexuelle de ses proies ou en se faisant passer pour des bactéries bioluminescentes dont se nourrissent ses proies. C’est ainsi que ce prédateur piège ou « pêche » ses proies en les attirant directement jusqu'à sa gueule. Cette technique utilise peu d’énergie et est rentable.
- Mimétisme agressif : Un mimétisme agressif est un mimétisme qui consiste, pour un prédateur, à ressembler à une espèce inoffensive pour leurrer ses proies[7],[8].
- Camouflage : Le camouflage permet au prédateur de se confondre dans son environnement afin de ne pas se faire repérer par sa proie.
Les étapes de la prédation
Les stratégies de prédation peuvent se décliner en quatre grandes phases qui sont la détection, l’approche et l’attaque, la capture ainsi que la consommation. Le déclenchement de la prédation est dû à des signaux internes métaboliques communément appelés faim et implique une vigilance sensorielle du prédateur.
Détection
- Rencontre et détection : Dans la plupart des cas, la phase de recherche est une étape très importante pour les prédateurs mobiles, cette phase requiert leur vigilance totale surtout pour les proies les plus petites qui sont difficilement détectable. Afin que cette détection soit fructueuse, le prédateur doit posséder des sens développés tel qu’un excellent odorat, une vision de longue portée, une ouïe fine et un toucher sensible. Pour ce faire il est nécessaire de posséder des organes sensoriels tels que les fossettes thermosensibles chez les crotales qui lui permettent de détecter les radiations infrarouges émises par les proies potentielles qui sont proches de lui. Cette étape est réduite pour les chasseurs à l’affût car c’est la proie qui arrive au prédateur. Les prédateurs s’adaptent aux rythmes biologiques de leurs proies, notamment leurs rythmes circadiens.
- Identification : Dans cette étape, le prédateur reconnaît une autre espèce animale comme étant une de ses potentielles proies.
Approche/Attaque
Les proies les plus faibles (jeunes, âgées, malades ou blessées) sont généralement les plus touchées par la prédation, cette sélection peut être anticipée ou non anticipée.
On définit une sélection anticipée comme une sélection où le prédateur teste ces proies et sélectionne la plus faible (exemple : la hyène fait courir ses proies pour tester leurs capacités individuelles de fuite). Dans la sélection non anticipée, ils attaquent leur proie sans véritable test, en définitive la proie la plus faible sera celle qui sera attrapée la première.
Il y a deux types d’approches :
- par embuscade : elle permet de surprendre la proie et est souvent utilisée par les chasseurs à l’affût, souvent peu endurants ;
- par poursuite : elle permet d’optimiser la distance pour l’attaque et est spécifique pour les prédateurs mobiles solitaires.
Dans les deux cas les individus doivent minimiser les mouvements visibles à l’approche de la proie. Les hiboux, par exemple, ont une structure d’ailes spécifique pour rester silencieux. Par la suite, le prédateur peut déclencher l’attaque[9].
Capture ou Subjugation
La capture et la mise à mort de la proie nécessitent une morphologie particulière, comme des crocs, des griffes, des techniques spécifiques.
Consommation
C’est le moment où le prédateur consomme sa proie (fruit de ses efforts) tout en restant vigilant à l’approche d’une espèce qui a un penchant pour le racket (la hyène par exemple).
Réponses fonctionnelles de Holling

La réponse fonctionnelle d'un prédateur aux variations de densité de la proie se mesure par les variations du nombre moyen de proies ΔN consommées par individu et par jour. On distingue trois types de réponses fonctionnelles. (Holling, 1959) [10],[11].
Il existe plusieurs stratégies de prédation. Ces stratégies sont fonction de l’environnement et des perturbations qu’elle subit. On retrouve parmi elles la réponse fonctionnelle de type I. Cette stratégie est la plus simple, le taux de capture croit en fonction de la densité de proie. Ensuite on retrouve la stratégie de type II, qui prend en compte le temps de manipulation de la proie. Le taux de capture décroît donc avec l’augmentation de la densité de proies.
Il existe une dernière réponse fonctionnelle qui est en lien direct avec l’évolution des stratégies de prédation. En effet, le prédateur choisit préférentiellement les proies les plus abondantes. Son taux d’attaque croît avec l’augmentation du nombre de proies à faible densité de proies (densité dépendance positive) et son taux d’attaque diminue avec l’augmentation du nombre de proies à forte densité (densité dépendance négative). Le prédateur adapte donc sa stratégie de prédation en fonction de son environnement. Si son environnement possède peu d’une telle proie, il va apprendre à le reconnaître et à « fourrager » (action de foraging) en priorité une autre proie dans le cas où son environnement lui offre d’autres alternatives (bascule de proies, switching).
La réponse fonctionnelle de type I
Il s’agit d’une relation linéaire qui suit le modèle de Lotka-Volterra : le nombre de proies capturées augmente linéairement avec l’accroissement de ces dernières. Le taux de mortalité des proies dû à la prédation est donc constant. Il s’agit d’un modèle assez simplifié mais qui est plutôt représentatif pour certains organismes filtrants comme les daphnies ou encore pour certains chasseurs passifs comme l’araignée (en effet, le nombre d’insectes capturés sur la toile dépend directement de la densité des populations d’insectes proies).
La réponse fonctionnelle de type II
Le nombre de proies consommées augmente d’abord rapidement lorsque l’effectif de la proie augmente mais atteint ensuite un plateau. Il faut prendre en compte dans cette relation le fait que le prédateur puisse arriver à satiété ainsi que le temps passé par le prédateur à chercher ses proies, à les manipuler, à les manger et à les digérer, ce qui affecte le taux de consommation des proies. Il s’agit de la relation la plus typique, elle est assez représentative pour un certain nombre d’animaux comme le lion ou même l’Homme.
Ces prédateurs sont spécialistes d'un groupe restreint de proies.
La réponse fonctionnelle de type III
Lorsque l’effectif d’une proie est faible, les prédateurs qui suivent une réponse fonctionnelle de type III en consomment très peu. Pour de tels prédateurs, la consommation la plus élevée de la proie se manifeste lorsque l’effectif des proies est intermédiaire.
Lorsque leur effectif est faible, les proies subissent donc un relâchement de la pression de prédation. Par exemple, de nombreux prédateurs répondent aux émissions de kairomones (substances chimiques émises par les proies) et accroissent leur activité lors de fortes émissions par les proies.
Il s'agit donc de prédateurs généralistes ou opportunistes, capables de diversifier les proies qu'ils chassent.
Évolution du trait stratégie de prédation
L'évolution du trait "stratégie de prédation se caractérise sous plusieurs forme. Nous pouvons illustrer l’évolution des stratégies de prédation par l’évolution de la prédation en fonction de la taille des proies. Le prédateur va préférentiellement prédater une proie d’une taille intermédiaire afin de maximiser l’apport énergétique apporté par la proie et minimiser les pertes d’énergies. Nous avons donc bien une évolution comportementale de la stratégie de prédation avec l’apparition de la capacité du prédateur à reconnaître la taille des proies et choisir de manière active la plus adéquate. L’évolution des stratégies de prédation peut également avoir comme support la génétique. En effet, certains traits favorisant la prédation sont sélectionnés (comme l’optimisation du taux d’attaque) et vont engendrer un effet pléiotrope.
Il est nécessaire de considérer la sélection au niveau de l’individu et la sélection au niveau de groupe dans l’évolution des stratégies de prédation. Afin de connaître l’effet de la sélection au niveau du groupe sur la stratégie de prédation, on intègre dans le modèle de Lotka-Volterra la densité-dépendance négative dans la croissance des proies.
- dN/dt = rN - N*N/K – aNP
- dP/dt = BNP – bP
Où N = densité de la proie
- P = densité du prédateur
- a = taux d’attaque du prédateur
- aN = réponse fonctionnelle du prédateur
Ainsi à l’équilibre nous avons : (dP/dt=dN/dt=0)
- N* = b/B et P* = r/a (1-N/K)
Dans le cas où la proie s’adapte à la prédation en augmentant ses capacités démographiques, et qu’il y a sélection au niveau individuel par le prédateur pour une augmentation de r et K chez la proie : la croissance plus rapide de la proie est entièrement bénéfique pour le prédateur, qui ne fournira plus assez d’effort pour la capture de sa proie car il est en quantité suffisante. Ceci est représenté dans les valeurs à l’équilibre de la proie et du prédateur où l’augmentation de r et N permet sa forte densité à l’équilibre. Donc la sélection par la prédation au niveau du groupe ne peut pas favoriser l’augmentation de r et K chez la proie. Ainsi les stratégies de prédations qui seront sélectionnées sont celles qui favoriseront la croissance de la proie c’est-à-dire les stratégies au niveau individuel chez le prédateur.
Cependant, la sélection individuelle et de groupe converge vers une réduction du taux de prédation a, ou à une augmentation du taux de mortalité b du prédateur (stratégies anti-prédation).
Il faut néanmoins noter que la dynamique des populations est infiniment plus rapide que la dynamique évolutive.
Course aux armements
Il est impossible de parler de l'évolution des stratégies de prédation sans introduire la notion de coévolution entre les couples prédateurs-proies.
La course aux armements se base sur le concept de coévolution en écologie. En effet, l’évolution, comme décrite par Charles Darwin, implique une sélection au sein du vivant, des individus les mieux adaptés aux conditions biotiques et abiotiques. Ces individus les plus optimaux (pour un prédateur les critères seront par exemple la rapidité et l’efficacité à attraper la proie) ont une plus grande fitness et sont donc avantagés, à long terme la proportion de ces individus augmente dans la population. Au sein du couple proie-prédateur, chacun subit la sélection naturelle et des stratégies d’optimisation de la prédation (pour les prédateurs) ou de défenses face aux prédateurs (pour les proies) sont sélectionnées. À chaque apparition d’un nouveau trait chez un des partenaires du couple proie-prédateur, l’autre partenaire doit « surenchérir » par une nouvelle stratégie (s’adapter par sélection naturelle) impliquant l’apparition d’un nouveau trait qui contrera à son tour la stratégie de son partenaire (contre-adaptation). Cette surenchère constante d’un des partenaires peut aussi être appelée course à l’armement. Cette expression illustre bien le concept de coévolution montrant qu’à chaque nouveauté du partenaire, l’autre doit contrecarrer cette dernière. Ce mécanisme est un mécanisme continu et évolutionniste dans lequel le couple proie-prédateur reste inchangé[12].
Cette coévolution peut bien entendu avoir une fin. Cette fin arrive sous deux conditions, soit un des partenaires du couple arrive à extinction, soit un des partenaires atteint un optimum qui ne pourra pas être déstabilisé par une nouvelle adaptation. Certaines coévolutions peuvent ne pas avoir de fin stable et cette surenchère peut continuer inlassablement. Par exemple, le lièvre et le lynx subissent la coévolution et les variations en abondances de ces deux espèces sont régulières selon un cycle ayant une périodicité de 9 à 11ans[13].
Cette coévolution implique donc l’adaptation des prédateurs comme décrit auparavant mais aussi des proies. L’adaptation des proies s’illustre par des défenses anti-prédation.
Héritabilité
L’héritabilité est la capacité de transmission d’un trait à un autre individu de son espèce. L’héritabilité des stratégies de prédation intervient lors de la transmission des capacités liées à la prédation d’un individu à sa descendance. Le trait peut être hérité génétiquement ou acquis par apprentissage.
Voir aussi
Articles connexes
Notes et références
- P. A. Abrams, The evolution of predator-prey interactions: theory and evidence, Annual Review of Ecology and Systematics, 2000, p. 2-4.
- C. Tirard, R. Barbault, L. Abbadie, N. Loeuille, Mini manuel d'écologie, cours+QCM/QROC, Dunod, 2012, p. 33-37
- R. W. Elner, Roger N. Hughes Energy Maximization in the Diet of the Shore Crab, Carcinus maenas Journal of Animal Ecology, vol. 47, no 1, février 1978, p. 103-116 (British Ecological Society
- S. Bengtson, Origins and early evolution of predation. In M. Kowalewski, P. H. Kelley, The fossil record of predation in The Paleontological Society Papers no 8, The Paleontological Society, 2002, p. 289–317
- P. A. Abrams, Implications of flexible foraging for interspecific interactions: lessons from simple models, Functional Ecology, 24(1), 2010, p. 7-17.
- L. M. Forster, Vision and prey-catching strategies in jumping spiders, Am Sci 70, 1982, p. 165–175
- J. Lloyd, Occurrence of Aggressive Mimicry in Fireflies The Florida Entomologist, vol. 67, no 3, septembre 1984, p. 368-376
- R. R. Jackson, Aggressive mimicry, prey-specific predatory behaviour and predator-recognition in the predator-prey interactions of Portia fimbriata and Euryattus sp., jumping spiders from Queensland, 1990
- E. Curio, The ethology of predation, Springer Science & Business Media, vol. 7, 2012.
- C.S. Holling, Some Characteristics of Simple Types of Predation and Parasitism, The Canadian Entomologist no 91, 1959, p. 385-398. doi:10.4039/Ent91385-7.
- C. S. Holling, The components of predation as revealed by a study of small mammal predation of the European Pine Sawfly, The Canadian Entomologist, vol. 91, 1959, p. 293-320.
- R. Dawkins, J. R. Krebs, Proc. R. Soc. Lond. B, 1979, Arms Races between and within Species, p. 489-511 ; DOI: 10.1098/rspb.1979.0081.
- N. C. Stenseth, W. Falck, O. N. Bjørnstad, C. J. Krebs, Population regulation in snowshoe hare and Canadian lynx: Asymmetric food web configurations between hare and lynx, Proceedings of the National Academy of Sciences of the United States of America, 1997;94(10), p. 5147-5152
 Portail de la biologie
Portail de la biologie  Portail origine et évolution du vivant
Portail origine et évolution du vivant