Évolution du mimétisme chez les insectes
Les insectes (embranchement des Arthropodes) possèdent trois paires de pattes et ont un corps divisé en tagmes (la tête, le thorax et l’abdomen). Leur apparition est estimée à plus de 400 millions d’années (Dévonien inférieur). Le nombre d'espèces connues est estimé à plus de 1,5 million, mais il en existerait 10 fois plus : les insectes sont donc les animaux les plus nombreux sur Terre. Au cours du temps, certains insectes ont développé des mécanismes de défense comme le mimétisme, afin d’en tirer un avantage adaptatif. Les individus mimétiques, en raison de leur apparence ou de leur comportement, peuvent se confondre avec des individus d'espèces différentes ou encore avec leur biotope. Il existe trois intervenants dans le mimétisme :
- le mime, qui peut prendre l'apparence ou imiter le comportement d’un individu d’une autre espèce, ou encore du milieu dans lequel il se trouve ;
- le modèle, qui peut correspondre à un individu toxique ou qui n'est pas palatable (désagréable à la consommation), ou encore à un élément abiotique ;
- le dupe, qui est en général un prédateur, et dont sa survie dépend de sa capacité à détecter le mime du modèle.
Pour les articles homonymes, voir Mimétisme (homonymie).
L’évolution du mimétisme chez les insectes peut s’explique par l’apparition de mutations positivement sélectionnées qui se transmettent dans la population via plusieurs mécanismes. Plusieurs espèces ont pu développer différents mimétismes qui leur sont propres.
Historique

Le naturaliste anglais Henry Walter Bates[1] fournit une première confirmation à la théorie de l'évolution de Darwin, en publiant un article sur la théorie du mimétisme, « The imitation by a species of other life forms or inanimate objects », à la suite de son voyage en Amazonie de 1848 à 1859. Il donne une explication au phénomène du mimétisme en montrant qu'une espèce non toxique de papillon amazonien, de la sous-famille des Dismorphiinae, a évolué en imitant des couleurs semblables à celles d'une tout autre espèce, de la sous-famille des Heliconiinae, négligée par les oiseaux à cause de sa toxicité. Ce phénomène a été nommé mimétisme Batésien. Le mécanisme proposé par Bates fut pendant longtemps un des exemples les plus frappants de sélection naturelle. Mais Bates n'a pas su répondre à la question " pourquoi deux espèces toxiques se co-miment ". Il a fallu attendre le biologiste allemand Johann Friedrich Theodor Müller[2], lui aussi partisan de Darwin. Il publia un article en 1879 dans lequel il définit pour la première fois le mimétisme entre deux espèces toxiques. Pour cela, il étudia des groupes de papillons toxiques sud-américains qui présentaient une ressemblance parfaite. Ce mimétisme est communément appelé mimétisme Müllerien.
Les différentes formes de mimétisme chez les insectes
Le mimétisme Batésien
Généralement, les espèces plus ou moins toxiques émettent un signal d'avertissement clairement perceptible par les prédateurs. Le plus souvent, il s'agit d'une coloration, mais le signal peut aussi être chimique ou sonore. Cette stratégie adaptative est appelée l'aposématisme. Dans le cas du mimétisme Batésien, des espèces non toxiques adoptent l'apparence (couleurs, motifs...) d'une espèce toxique. Ainsi, l'organisme mimétique acquiert une protection contre la prédation. D'autre part, le mime peut aussi copier le comportement du modèle pour que la supercherie soit optimale. Par exemple, la guêpe Gelis agilis - le mime - ressemble fortement à une espèce de fourmi noire Lasius niger - le modèle. De plus, des expériences ont montré qu’elle pouvait aussi émettre des signaux chimiques (hormones) comme les fourmis qui lui servent de modèle[3].
Les mimétismes "bon" et "imparfait"
Dans la nature, il y a des organismes très ressemblants à leur modèle, c’est ce que l’on appelle un "bon" mimétisme. Un "bon" mimétisme peut être illustré avec le cas des syrphes qui miment les hyménoptères. Par exemple l’espèce Volucella bombylans plumata a une fidélité mimétique forte avec son modèle Bombus terrestris[4]. Mais il existe aussi un mimétisme dit "imparfait". Ce cas est aussi rencontré chez les espèces de syrphes qui adoptent l’apparence des hyménoptères comme les guêpes, les abeilles ou encore les bourdons. En effet, certaines études ont montré que le mimétisme avait une relation avec la taille. Les scientifiques ont pu mettre en évidence que dans une population de syrphes, la taille des individus est variable, et que les petits individus avaient une fidélité mimétique moins forte que celle des grands individus. Plusieurs hypothèses ont été avancées pour le maintien des formes imparfaites, notamment celle de la sélection détendue. Celle-ci suppose que les individus de petite taille n’ont pas besoin d’investir beaucoup de leur énergie dans un « bon » mimétisme. En effet, du fait de leur taille, ils ne sont pas les cibles principales des prédateurs[5].
Course aux armements
Le mimétisme Batésien peut être perçu comme une forme de parasitisme de l’individu mime envers son modèle. En effet, le mime copie le modèle, augmentant ainsi ses chances de survie, et donc sa probabilité de transmettre ses gènes à sa descendance. La population de mimes va donc croître, et alors augmenter les chances du prédateur de manger une espèce à l’aspect nocif, mais qui en réalité est comestible. Deux conséquences peuvent alors survenir :
- soit le prédateur va perdre sa capacité à apprendre, donc continuer à chasser les proies nocives et leurs mimes : ce sont alors les modèles qui vont subir à nouveau une pression de prédation ;
- soit le prédateur va acquérir la capacité à distinguer le mime du modèle : ce sont ici les mimes qui vont subir à nouveau une pression de prédation.
Dans le premier cas, le modèle va devoir s’adapter en acquérant de nouvelles défenses, que le mime finira par « copier ». Dans le deuxième cas, la présence du modèle est indispensable pour que cet avantage soit sélectionné. Le mime peut contrer ce désavantage. Pour cela, la sélection naturelle sélectionne au sein de la population, les individus les plus ressemblants aux modèles augmentant ainsi leur fidélité mimétique. À la suite de cela, le prédateur ne sera donc plus capable de déterminer avec certitude le mime du modèle. Une nouvelle fois, le modèle devra acquérir de nouvelles défenses pour augmenter ses chances de survie. Il en résulte donc une co-évolution entre le mime et le modèle.
Le mimétisme Müllerien

Le mimétisme Müllérien[6],[7],[8] est une forme de mimétisme se référant à deux espèces toxiques qui s’imitent l’une et l’autre avec le même motif d’avertissement. Ainsi, tous les mimes partagent les avantages du pattern puisque le prédateur reconnaît toutes les espèces toxiques après quelques mauvaises expériences. Chaque espèce toxique doit “sacrifier” un certain nombre d’individus pour l’éducation des prédateurs afin d’être reconnue comme toxique. C’est le cas des papillons diurnes toxiques Ithomidae et Héliconidae qui en se ressemblant, améliorent la reconnaissance de leur code couleur par les prédateurs.
Dans la pratique, il est difficile de distinguer un mimétisme Batésien d'un mimétisme Müllérien. En effet, la palatabilité d'un insecte pour son prédateur est difficile à évaluer. De plus, si les deux mimes n'ont pas une toxicité strictement similaire, le mime le moins toxique pourrait bénéficier davantage du mimétisme que le mime le plus toxique. Il peut alors être difficile à évaluer si le mimétisme est un bénéfice pour les deux mimes ou uniquement pour l'espèce la moins toxique.
Le mimétisme Mertensien
Le mimétisme Mertensien, découvert par Wolfgang Wickler en 1968 et nommé en référence à Robert Mertens, est un cas moins connu de mimétisme. Il permet à une espèce mortelle d'adopter l'apparence d'une espèce moins dangereuse. Ainsi, si l'espèce mortelle est un prédateur, elle acquiert un avantage dans l'accessibilité de ses proies. D'autre part, ce mimétisme peut avoir un fonctionnement similaire à celui du mimétisme Batésien. En effet, en adoptant un motif d'avertissement d'une espèce moins dangereuse, une espèce mortelle peut bénéficier de l'apprentissage d'évitement des prédateurs envers les espèces moins toxiques. Ce mimétisme n'a pas encore été observé chez les insectes.
L'auto-mimétisme
.jpg.webp)
Dans le cas de l’auto-mimétisme[9], les insectes utilisent certaines parties de leur corps pour imiter d’autres parties de leur propre corps ou du corps d'autres animaux, souvent les yeux.
Par exemple, de nombreux papillons nocturnes tel que Caligo idomeneus ont des taches simulant un œil (appelées ocelles), ayant pour but d’effrayer le prédateur et/ou de créer un effet de surprise pour donner à la proie le temps de fuir. Le phénomène d’auto-mimétisme est analogue au mécanisme du mimétisme Batésien parfait. Dans les deux cas, l'avantage mimétique dépend de la proportion des mimes non comestibles, de la durée de la mémoire des prédateurs et de l'abondance des mimes par rapport aux prédateurs. Cependant, les bases de l'évolution de ces deux phénomènes sont différentes.
Le camouflage ou cryptisme : une forme de mimèse
Dans la nature, les cas de mimétismes sont très fréquents. Il faut néanmoins faire une différence entre le mimétisme et la mimèse qui regroupe les cas de cryptisme ou de camouflage. La mimèse permet à des espèces de se fondre dans leur environnement. En effet, l’individu mimétique adopte l’apparence de son biotope et reste immobile pour passer inaperçu aux yeux de ses prédateurs. L’homochromie fait partie du camouflage. En effet, l’individu adopte les couleurs de son environnement proche.

On parle d’homochromie si la teinte prise par l’animal est uniforme et correspond bien à l’environnement avoisinant. En plus de la couleur, certains animaux vont beaucoup plus loin. En effet, en plus de prendre la même coloration que son biotope, certains organismes sont aussi capables de prendre la forme de l’objet sur lequel il se tient le plus souvent. On parle alors d’homotypie. On peut ainsi trouver de nombreux insectes qui prennent l’apparence d’écorce ou de feuille. C'est par exemple le cas du phasme. Cet insecte prend généralement la couleur et la forme d’une brindille pour se confondre avec son environnement. Cela confère à l’individu une protection contre la prédation, c’est une mimèse défensive.
On peut aussi trouver des cas de mimèse offensive, qui permet à l’individu mimétique de chasser ses proies sans être repérés. On retrouve ce cas de mimèse chez les mantes-fleurs. Mais on peut aussi trouver une variante de l’homochromie simple, c’est l’homochromie variable. Cela permet aux animaux de prendre les couleurs de leur environnement à tous moment. Mais, pour l’instant, il n’y a aucun exemple dans la littérature de ce type de mimèse pour les insectes.
Les mécanismes d'apparition et de maintenance d'une forme mimétique
Les mutations
Les mutations sont à l'origine de processus d'évolution au sein d'une population. Lorsqu'une mutation affecte une cellule germinale d'un individu et que celle-ci n'est pas réparée, elle peut se transmettre aux générations suivantes. C'est un événement stochastique rare, spontané et indépendant du besoin de l'espèce touchée, qui se propage au cours du temps, ou disparaît via le phénomène de sélection naturelle. Les mutations peuvent conférer un avantage sélectif en améliorant la fitness d'un organisme, lui permettant d'étendre sa répartition au sein d'une population.
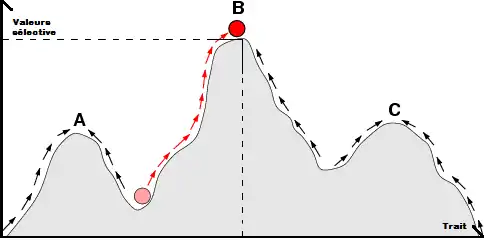
Les mutations peuvent générer l’apparition de nouveaux allèles au sein d’une population. Ceux-ci peuvent être dominants ou récessifs. La dominance des allèles[10] est souvent le résultat de leur expression ou de leur activité. Étant donné que les nouveaux allèles apparaissent de façon hétérozygote, la sélection de la dominance est importante pour la propagation de ces nouveaux allèles. En effet, d'après la théorie du tamis de Haldane (Haldane’s sieve), la sélection est plus efficace quand le nouvel allèle est dominant.
Pour qu’une mutation soit sélectionnée, il faut que le nouveau trait présente une valeur sélective supérieure à celle du trait de l’individu résident. La mutation doit donc apporter un avantage adaptatif.
Notion de supergène
Un supergène est un ensemble de gènes voisins sur un même chromosome qui possèdent un lien génétique étroit. Les supergènes peuvent résulter d'une inversion d'une partie d'un chromosome qui immobilise un ensemble de plusieurs gènes adjacents. Ces épisodes d’inversion empêchent la recombinaison chromosomique lors de la formation des cellules sexuelles par absence d'homologie. Le supergène est donc transmis tel quel aux descendants. Des études ont mis en évidence l'existence de supergènes contrôlant le pattern mimétique chez les insectes. Le cas du papillon Heliconius numata, mime Müllerien du papillon appartenant au genre Melinaea illustre ce phénomène. En effet, un bloc de 30 gènes composant un supergène (P25) responsable du mimétisme a été découvert[11]. L’inversion de ce bloc de gènes a été mise en évidence, interdisant le mélange génétique. Il existe au moins trois types de supergènes chez Heliconius numata où seul l'ordre des gènes dans le bloc diffèrent, entraînant plusieurs variants phénotypiques. Chacun des variants de Heliconius numata ressemble à une espèce différente de Melinaea. Sans cette inversion, chaque gène serait indépendant et soumis à la recombinaison, entraînant ainsi une grande gamme de phénotypes sans lien avec Melinaea.
Il existe d’autres exemples pour l’apparition du mimétisme qui diffèrent de celui d’Heliconius numata. C’est ce qu’ont observé Haruhiko Fujiwara et ses collègues[12] en travaillant sur Papilio polytes (Mormon commun). Ce papillon, non toxique est un mime Batésien de l'espèce toxique Pachliopta aristolochiae. Le gène doublesex (dsx) est un facteur de transcription régulant la détermination du sexe chez les protostomiens et deutérostomiens. L'apparition d'une inversion dans le gène dsx a été observée chez des femelles de Papilio polytes. Après cette mutation, l'allèle H (allèle mimétique de dsx, noté dsx(H)), agirait sur l'expression des gènes responsables du pattern des ailes. Il est remarquable de noter que dans ce cas, un seul gène est capable d’agir comme un supergène et de contrôler la totalité du pattern des ailes. Ce papillon présente un polymorphisme et un dimorphisme, seules les femelles sont mimétiques. Le phénotype des femelles non mutantes est semblable aux mâles, et est déterminé par d'autres gènes que dsx. Les femelles porteuses de cette mutation ont un phénotype ressemblant à l'espèce modèle , et il n’existe pas de formes intermédiaires. Dsx(H) est dominant ; le génotype des mutantes peut alors être hétérozygote (Hh) ou homozygote (HH), (h l’allèle ancestral récessif)[13].
La sélection naturelle
Charles Darwin révolutionna le domaine de la science avec son ouvrage « L'Origine des espèces » publié en 1859, où il présente sa théorie de l'évolution. Aujourd'hui, l'évolution se définit comme un ensemble de changements qui surviennent au cours du temps, à la suite de mutations. Ce processus est subi par les espèces, et permet l’apparition de nouvelles formes de vie. Cela permet d'expliquer en partie la diversité du monde vivant. Pour Darwin, le moteur principal de sa théorie de l'évolution est la sélection naturelle. Ce mécanisme fournit un avantage adaptatif aux individus les mieux adaptés à un environnement donné, augmentant ainsi leur fitness, c’est-à-dire leur assurant une plus grande chance de transmettre leurs gènes à la génération suivante. En effet, les mutations qui entraînent l’apparition de nouveaux caractères doivent être avantageuses pour être sélectionnées. Il arrive parfois que les mutations entraînent l’apparition d’allèles délétères, entraînant des individus malades ou peu adaptés à leur environnement. Dans ce cas-là, la sélection naturelle va écarter ces individus des circuits de reproduction. La théorie de l'évolution permet ainsi d'expliquer et de comprendre comment l'environnement peut influer sur l'évolution des espèces, mais aussi sur celle des populations.
La sélection naturelle concerne des caractères qui ont un impact sur la survie ou le succès reproductif. Pour qu’elle soit fonctionnelle, le caractère doit :
- être variable, par exemple, la couleur des ailes chez les papillons varie en fonction de la zone de répartition géographique et de la présence du modèle ;
- présenter un avantage sélectif, c'est-à-dire qu'il augmente la probabilité de transmettre ses gènes à la génération suivante et donc sa fitness ;
- être héritable, donc qu'il soit transmissible à la génération suivante.
La sélection naturelle est le mécanisme de maintien d'un caractère dans une population pour de nombreuses espèces notamment dans le cas du mimétisme. Elle va favoriser les individus présentant un avantage adaptatif comme une fidélité mimétique forte, augmentant ainsi leur chance de survie et donc leur succès reproducteur. Un autre mécanisme possible est la sélection sexuelle qui semble également jouer sur le maintien d’une forme de mimétisme dans une population. Un isolement reproductif (union homogame) a été observé chez des insectes comportant le même motif d’ailes. Le pattern sert alors de repère dans le choix de partenaires. La propagation de gènes codant un même pattern est donc facilitée[14].
La sélection par prédation
Les prédateurs naissent naïfs puis apprennent au cours de leur vie à éviter les proies toxiques. Ce comportement d’apprentissage d’évitement nécessite une phase où le prédateur teste un nombre de proies avant d’apprendre à éviter celles qui lui sont désagréables. Pour que l'apprentissage soit efficace, l'ingestion de la proie ne doit pas être létale. Dans le cas du mimétisme batésien, le mime profite du comportement d'évitement du prédateur face à l'espèce modèle qui est non palatable (fréquence dépendance négative). Le fonctionnement du mimétisme batésien repose sur une fréquence du mime moindre par rapport au modèle.
- Le maintien du mimétisme batésien au sein d’une population dépend en grande partie du mécanisme de fréquence-dépendance. En effet, un mime va adopter l’apparence de l’espèce modèle en reproduisant son message d’alerte. Pour que cette stratégie perdure au cours du temps, il faut que les mimes soient moins fréquents dans un environnement donné que les modèles. Dans le cas contraire, les prédateurs auraient plus de chance de rencontrer un mime et d’associer son pattern à une espèce comestible, ce qui produirait un réel désavantage pour les deux populations. La sélection d’une forme par la sélection naturelle va donc dépendre de la proportion de modèles dans la population totale.
« Le mimétisme est la conséquence de la sélection imposée par la prédation, par le partage des coûts dans l'enseignement des prédateurs naïfs » Müller[2]. Des co-mimes müllériens tirent leur protection par la force du nombre qui inclut un partage des coûts face aux prédateurs. Le processus fonctionne après le "sacrifice" commun d’individus co-mimes (densité-dépendance positive). Dans la nature, le pattern de couleur d’une espèce toxique est en général connu des prédateurs, la pression de prédation exercée sur cette espèce est donc minimale. Or, si cette même espèce toxique change son patron de couleur, les prédateurs ne seront plus en mesure de le reconnaître et devront donc subir une pression de prédation plus forte. Ainsi, l’apparition de forme rare ou hybride au sein d’une espèce toxique est désavantagée, il en résultera une sélection stabilisante. Ainsi, la sélection par prédation sera anti-apostatique, c’est-à-dire contre la rareté[15].
- Le maintien du mimétisme müllérien au sein d’une population dépend en grande partie du mécanisme de densité-dépendance. Un prédateur attaque un nombre fixe de proies avant d'apprendre à l'éviter. Plus il y a de phénotypes semblables, plus le prédateur apprend vite. Donc le cumul des densités des populations de co-mimes va favoriser un apprentissage d’évitement rapide des prédateurs naïfs. Dans un premier temps, les deux espèces co-mimes partagent un coût commun d’individus sacrifiés pour l’apprentissage du prédateur ; ils vont donc coopérer. Puis, une fois l’apprentissage effectué, les deux co-mimes bénéficieront d’une survie et d’une fitness plus grande. Cette densité-dépendance positive favorise donc les phénotypes communs et contre-sélectionne les phénotypes rares. Ce mécanisme est aussi connu sous le nom d’effet Allee.
Polymorphisme spatial
Échelle géographique
À l’échelle du globe, une espèce mimétique présente une variation de morphes selon les aires de répartition [2]. Ainsi, on peut retrouver des mimes avec une large gamme phénotypique, répartis dans différents anneaux de mimétisme (nom donné aux communautés impliquées dans un système de mimétisme batésien et mullérien)[16]. On retrouve ce cas avec les espèces Heliconius erato et Heliconius melpomene qui présentent une co-évolution de leurs patterns en fonction des localités.
Échelle locale
Il n’est pas rare de trouver dans une même zone plusieurs schémas d’avertissement entrainant ainsi la formation d’une mosaïque spatiale. Cette observation pose un paradoxe. En effet, un mimétisme est d’autant plus efficace vis-à-vis du prédateur si les signaux ne diffèrent pas et si les formes sont communes (sélection anti-apostatique)[2]. Différentes explications sont proposées :
- les différents anneaux de mimétisme peuvent contenir des membres qui ne sont pas totalement chevauchants dans l’espace et dans le temps. Par exemple, on retrouve des patterns différents entre les mimétismes de papillons occupant la canopée et ceux de sous-bois ;
- le mimétisme peut également subir des contraintes morphologiques, qui imposent une limite dans la gamme de phénotypes possibles à imiter. C'est-à-dire que les individus possèdent des formes trop distinctes entre eux. Ainsi, l’apparition de nouveaux phénotypes intermédiaires serait donc contre-sélectionnée.
Notes et références
- Bates, Henry Walter. “Complete Dictionary of Scientific Biography”. 2008. Encyclopedia.com
- Thomas N.Sherratt et al. (2008). « The evolution of Müllerian mimicry ». Naturwissenschaften. ;95(8):681-95. doi: 10.1007/s00114-008-0403-y
- Miriama Malcicka et al. (2015). « Multi-trait mimicry of ants by a parasitoid wasp ». sci rep. 27;5:8043. doi: 10.1038/srep08043
- Edmunds M et al. (2014). « Evidence for Batesian mimicry in a polymorphic hoverfly ». evolution. 68(3):827-39. doi: 10.1111/evo.12308
- Wilson JS et al. (2013). “Ecological and evolutionary processes drive the origin and maintenance of imperfect mimicry”. PLoS One. 8(4):e61610. doi: 10.1371/journal.pone.0061610
- Malcolm Emmunds. (1999). « Why are the good and poor mimics ? » Biological Journal of the Linnean Society. Pages 459–466
- James Mallet. (2000).« Causes and consequences of a lack of coevolution in Mullerian mimicry ». Evolutionary Ecology 12. 13: 777±806, 1999
- Alexandra C. V. Balogh et al. (2005).« Müllerian mimicry : an examination of Fisher’s theory of gradual evolutionary change », in Proceeding of the Royal Society B. 7;272(1578):2269-75
- Lincoln P. Brower . (1970) .« Theoretical Investigations of Automimicry I. Single Trial Learning” Processdings of the National Academy of Sciences. ;66(4):1059-66
- Le Poul Y et al. (2014). « Evolution of dominance mechanisms at a butterfly mimicry supergene ». Nat commun. 5:5644. doi: 10.1038/ncomms6644
- Kunte K et al. (2014). « Doublesex is a mimicry surpergene ». Nature. ;507(7491):229-32. doi: 10.1038/nature13112
- Mallet J. (2015). « New genomes clarify mimicry evolution ». Nat Genet. 47(4):306-7. doi: 10.1038/ng.3260
- Kunte K et al. (2014). « Doublesex is a mimicry surpergene ». Nature. ;507(7491):229-32. doi: 10.1038/nature13112
- Parchem RJ et al. (2007) « Patterns on the insect wing » Curr Opin Genet Dev.17(4):300-8
- Alexandra C. V. Balogh. (2008).« Learning and the mimicry spectrum: from quasi-Bates to super-Müller », Animal Behaviour. Pages 1591–1599
- (en) Riddhi Deshmukh, Saurav Baral, A. Gandhimathi, Muktai Kuwalekar & Krushnamegh Kunte, « Mimicry in butterflies: co-option and a bag of magnificent developmental genetic tricks », Wiley Interdiscip Rev Dev Biol, vol. 7, no 1, (DOI 10.1002/wdev.291, lire en ligne).
 Portail de l’entomologie
Portail de l’entomologie