Bactérie magnétotactique
Les bactéries magnétotactiques (BMT) sont des micro-organismes, découverts en 1975 par R. P. Blakemore, capables de se déplacer le long de lignes d'un champ magnétique[1]. Elles contiennent des organites, les magnétosomes, qui sont à l'origine de leurs propriétés magnétiques. De par leurs possibles applications médicales et biotechnologiques, elles intéressent aujourd'hui les scientifiques.
Historique
Les bactéries magnétotactiques ont été découvertes en 1975 par R. P. Blakemore, alors étudiant en thèse à l'Université du Massachusetts. Il étudiait des populations de bactéries retrouvées dans les sédiments lacustres et marins. Il fut étonné par un mouvement uniforme de certaines bactéries, et démontra grâce à un aimant que ce mouvement était dû au champ magnétique[2].
La première espèce isolée en culture pure est Magnetospirillum magnetotacticum, souche MS-1, aux États-Unis, par Blakemore et al.. Trois autres espèces ont par la suite été isolées : Magnetospirillum magneticum, souche AMB-1 et Desulfovibrio magneticus RS-1 au Japon, et Magnetospirillum gryphiswaldense, souche MSR-1 en Allemagne[3].
La recherche sur ces bactéries est rendue difficile par le faible nombre de souches que l'on a découvertes et les difficultés que l'on a à les isoler en culture. Cependant, des découvertes récentes en matière de techniques pour la culture de ces bactéries ont permis aux chercheurs une avancée dans la recherche relative à ces organismes[4].
Description biologique
Caractéristiques biologiques
Les BMT sont des procaryotes, de type Gram-négatif, caractérisées par leur capacité à produire des magnétosomes[5]. Elles sont anaérobies, voire microaérophiles selon les espèces[4]. De ce fait, on les retrouve principalement dans les sédiments lacustres ou marins, enfouies à l'interface oxique-anoxique (frontière entre le milieu riche en oxygène et celui dépourvu d'oxygène)[5],[6].
Toutes les BMT découvertes à ce jour sont motiles, grâce à la présence de flagelles[5]. Elles peuvent ainsi se déplacer par aérotaxie, c'est-à-dire en réaction à la concentration d'oxygène du milieu, afin de se diriger vers des lieux favorables à leur développement. Elles se déplacent également par magnétotaxie, c'est-à-dire qu'elles se dirigent selon les lignes du champ magnétique terrestre pour atteindre des milieux favorables à leur croissance. Ainsi, toutes les BMT qui se situent dans un même lieu vont se diriger dans la même direction selon les lignes du champ magnétique terrestre avec une vitesse de l'ordre de 40 à 1 000 μm·s-1[7].
Le séquençage du génome des BMT laisse penser qu'elles appartiendraient pour la plupart aux protéobactéries. Cependant, les différentes souches découvertes à ce jour ne sont pas issues d'un même ancêtre commun : les BMT représentent un taxon polyphylétique. Elles présentent donc de nombreuses morphologies : coccus, spirille, bacille, vibrio, voire multicellulaire[5].
Elles se distinguent également les unes des autres par leur synthèse de magnétosomes, de compositions différentes : certaines synthétisent des magnétosomes formés de magnétite (Fe3O4)[8], et appartiennent pour la plupart au groupe Alpha des protéobactéries[9], d'autres synthétisent des magnétosomes de type greigite (Fe3S4)[10] et sont pour la plupart apparentées aux Delta protéobactéries[11]. Cependant, certaines souches observées sont capables de synthétiser les deux formes de magnétosomes à la fois[12]. De même, la souche cultivée RS-1 est rattachée aux Delta protéobactéries bien qu'elle synthétise des magnétosomes de magnétite[13].
Enfin, plusieurs formes de magnétosomes ont été observées : pseudo-cubique[14], pseudo-parallélépipédique[14], ou encore allongé en forme de balle de fusil[15]. Mais une même souche ne peut produire qu'une seule forme de magnétosome[16].
Biominéralisation des magnétosomes
Les bactéries magnétotactiques minéralisent les magnétosomes à partir d'oxyde de fer, ce qui donne de la magnétite ou à partir de sulfure de fer et forme de la greigite. Ce processus de minéralisation est certainement contrôlé chimiquement, car lorsqu'un milieu contient beaucoup de sulfure d'hydrogène, les bactéries produisant de la magnétite continuent de fabriquer cette dernière et non de la greigite. Les cristaux de magnétite ou de greigite ont typiquement une plus grande longueur de 35 à 120 nm[4].
La minéralisation de la magnétite dépend de nombreux facteurs, les plus importants étant la concentration en dioxygène et la présence d'oxydes d'azote[4]. Certaines BMT ont besoin d'être en milieu micro-aérobie pour pouvoir synthétiser de la magnétite. La production de magnétite de la bactérie est alors maximale lorsque la concentration en oxygène est aux alentours de 1 %, alors qu’une concentration supérieure à 5 % inhibe la production de magnétite[17]. L'expérience montre que l'oxygène servant à la synthèse de l'oxyde de fer est celui des molécules d'eau et non de l'oxygène moléculaire, ce qui pourrait expliquer que certaines BMT peuvent synthétiser de la magnétite en milieu anaérobie[18]. En revanche, la minéralisation de la greigite semble se faire uniquement en l’absence d’oxygène[3].
De toute évidence, la polyphylie des BMT laisse supposer différents moyens biochimiques de synthèse de minéraux magnétiques. Cependant, un transfert horizontal de gènes au cours de l’évolution peut également être envisagé pour expliquer cette polyphylie[4].
Les cristaux magnétiques fabriqués par les bactéries magnétotactiques sont d'une grande qualité, car ils sont d'une haute pureté chimique, et possèdent une bonne homogénéité[19]. La fabrication en laboratoire de tels cristaux reviendrait cher, c'est pourquoi les scientifiques cherchent à cultiver ces bactéries en laboratoire, mais cela demeure malaisé.
Magnétosomes
Les magnétosomes sont composés d'un cristal magnétique contenu dans une bicouche lipidique. Ces magnétosomes forment des chaînes dans le cytoplasme de la bactérie. Les cristaux magnétiques sont soit de la magnétite (oxyde de fer(II,III) FeO·Fe2O3) soit de la greigite (oxyde de soufre(II, III) FeS·Fe2S3), la taille de ces cristaux étant de l'ordre de 35 à 120 nm[4]. La formation des magnétosomes requiert trois étapes ordonnées, mais on ne connait pas l'ordre dans lequel la bactérie procède à ces étapes. Néanmoins, le processus de fabrication des magnétosomes reste très rapide, il est de l'ordre d'une dizaine de minutes[3].
L'absorption du fer par la bactérie
À l'état naturel ionique, il existe deux sortes d'ions fer, les cations fer(II) Fe2+ et les ions fer(III) Fe3+. Ces deux ions sont présents dans le milieu de croissance des bactéries magnétotactiques. Les ions Fer(II) étant très solubles dans l'eau, les BMT les absorbent à l'aide d'enzymes et protéines spécifiques, mais selon des procédés semblables aux autres bactéries[20]. Les ions Fer(III), eux, étant insolubles à pH neutre, les BMT doivent les chélater afin de les solubiliser. À l'instar de nombreux autres procaryotes, elles ont ainsi recours à des sidérophores[21],[22],[3]. Les sidérophores forment un complexe avec le fer(III), qui est alors solubilisé et transporté dans le cytoplasme de la cellule[4].
La formation de la membrane du magnétosome
La membrane du magnétosome est composée principalement d'une bicouche lipidique d'environ 3 à 4 nm d'épaisseur. Cette bicouche est constituée de phospholipides, d'acides gras et de protéines membranaires[23]. Les magnétosomes se forment au sein du cytoplasme des cellules, ce qui explique que la composition de la membrane magnétosomale soit très semblable à celle de la membrane plasmique des cellules. Néanmoins, on ne sait toujours pas si la membrane des magnétosomes est formée avant l'absorption du fer ou si elle est formée autour de cristaux de fer déjà présents dans la cellule[4].
Le transport du fer dans le magnétosome
La croissance des cristaux de fer au sein de la bactérie se fait lorsque les cristaux sont déjà présents dans le magnétosome, ce qui implique que le fer doit être transporté à l'intérieur des magnétosomes. Des protéines ont été retrouvées à la fois dans le cytoplasme des cellules et dans les magnétosomes, comme la protéine MagA présente notamment au sein de la souche AMB-1 de Magnetospirillum magneticum[23]. Ces protéines ont certainement un rôle dans le transport du fer de la cellule vers les magnétosomes.
Anomalie isotopique du fer des magnétosomes
Les magnétites des magnétosomes sont enrichies en 57Fe relativement aux isotopes pairs (54Fe, 56Fe et 58Fe)[24]. Des anomalies similaires avaient déjà été observées pour d'autres éléments comme le mercure ou le soufre, mais jamais pour le fer. Le mécanisme d'acquisition de cette anomalie n'est pas encore établi, mais il pourrait être lié aux propriétés électroniques de spin du fer. La recherche de cette anomalie pourra aider à l'identification des fossiles de bactéries magnétotactiques dans des échantillons terrestres voire extraterrestres.
Description magnétique
Les bactéries magnétotactiques sont des organismes micro-aérobies, elles ont donc besoin d’un milieu pauvre en dioxygène pour subsister. Pour trouver un milieu favorable à leur croissance, elles ont développé une capacité appelée magnétotaxie, c'est-à-dire qu’elles se servent des lignes de champ magnétique pour se déplacer.
Magnétotaxie
Comment fonctionne-t-elle ?
Les cristaux synthétisés par les bactéries mesurent entre 35 et 120 nm. Des cristaux de cette taille sont monodomaines, ce qui signifie que les moments magnétiques de chaque atome d’un cristal sont parallèles. (« On appelle domaine magnétique ou domaine de Weiss chaque région d’un seul tenant dans laquelle tous les moments magnétiques atomiques sont alignés parallèlement les uns aux autres »[25],). De plus, l’ensemble des cristaux adjacents au sein d’un magnétosome ont leurs moments magnétiques parallèles. Il en résulte un moment magnétique permanent pour le magnétosome, qui se comporte alors comme un aimant[3].
En raison de son moment magnétique permanent, le magnétosome subit un couple dans le champ magnétique terrestre. L'amplitude de ce couple dépasse les forces thermiques qui tendent à orienter les bactéries de manière aléatoire (par agitation). En conséquence, les bactéries magnétotactiques s’orientent selon les lignes du champ magnétique terrestre indépendamment du mouvement brownien. La magnétotaxie est donc l’alliance entre cette capacité passive à s’orienter (les bactéries, même mortes, restent orientées selon les lignes de champ), et à se déplacer grâce à leur flagelle[4].
En quoi leur permet-elle de s'orienter vers un lieu à bonne teneur en O2 ?
Les bactéries magnétotactiques se déplacent toujours le long des lignes de champ terrestre, car elles constituent les plus attractives pour ces bactéries. Dans l’hémisphère nord, elles ont tendance à suivre le sens positif du champ magnétique terrestre, on dit qu'elles nagent parallèlement à celui-ci (c'est-à-dire vers le nord), tandis que, dans l’hémisphère sud, elles ont plutôt tendance aller en sens inverse du champ magnétique, on dit qu'elles nagent antiparallèlement à celui-ci (c'est-à-dire qu’elles vont vers le sud). Certains types de bactéries magnétotactiques peuvent, dans certaines conditions encore méconnues, s'orienter vers le sud en étant au nord (ou inversement), voire changer de sens pendant le trajet. Elles ne peuvent cependant jamais se déplacer de façon non-parallèle aux lignes de champ terrestre.
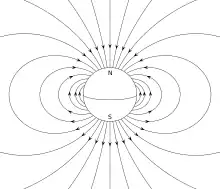
Or, le champ magnétique de la Terre étant incliné sous l’horizontale dans l’hémisphère nord, et au-dessus dans l’hémisphère sud, les bactéries qui suivent les lignes de champ en s'approchant de l'un des pôles ont tendance à "descendre". Elles migrent donc en s’enfonçant dans les sédiments le long des lignes de champ géomagnétique[4]. En regardant un schéma du champ magnétique terrestre (voir exemple ci-contre), on observe que les lignes de champ magnétique partent du noyau de la Terre, se dirigent vers le sud, puis partent vers le nord en formant des arcs de cercle avant de finalement revenir au noyau. De ce fait, lorsque les bactéries magnétotactiques au nord suivent les lignes de champ vers son pôle, elles ont tendance à s'enfoncer. Au sud, elles vont dans le sens contraire, et s'enfoncent par conséquent sous les sédiments.
Étant donné que la teneur en oxygène diminue avec la profondeur dans les milieux sédimentaires[26], les bactéries magnétotactiques trouvent dans la profondeur des couches sédimentaires un environnement propice à leur développement puisqu'elles sont pour la plupart micro-aérobies.
La magnétotaxie constitue donc un moyen de locomotion efficace et privilégié pour ces organismes, car il leur assure la quasi-certitude de pouvoir trouver un environnement au taux de dioxygène leur permettant de se développer.
Aérotaxie
Les bactéries magnétotactiques se servent de la magnétotaxie afin de trouver un environnement où la concentration en oxygène est optimale pour leur croissance, c’est pour cela qu’on les retrouve en milieux sédimentaires.
La première théorie formulée quant au bénéfice de la magnétotaxie pour ces bactéries était la suivante : la magnétotaxie permettrait de manière directe aux bactéries magnétotactiques de trouver leur niche écologique pour se développer[2].
Cependant, la découverte de nombreuses bactéries magnétotactiques à la limite oxique/anoxique[4] laisse entendre que la magnétotaxie n’est pas le seul moyen qu'elles ont de trouver un environnement viable.
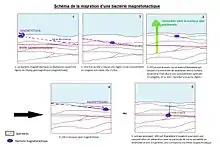
En effet, elles utilisent aussi l’aérotaxie, c'est-à-dire qu’elles migrent en fonction de la concentration en oxygène, et pas seulement par rapport aux lignes de champ magnétique. Par exemple, lorsqu’une bactérie magnétotactique utilise la magnétotaxie, elle s’enfonce sous les sédiments jusqu'à trouver un milieu avec la bonne teneur en oxygène et s’y fixe[7]. Le problème est que parfois les colonnes sédimentaires (voir schéma ci-contre), qui possèdent de forts gradients rédox, subissent une érosion ou d’autres phénomènes qui ont pour conséquence la remontée vers la surface des sédiments, et donc par exemple de bactéries magnétotactiques qui y vivent. Étant donné l’évolution de la concentration en oxygène en fonction de la profondeur, une bactérie magnétotactique qui est remontée vers la surface avec les sédiments ne se trouve plus dans un milieu de concentration en oxygène optimale.
La question est donc de savoir comment la bactérie va retrouver une région avec des conditions optimales de développement. Soit elle utilise à nouveau la magnétotaxie et trouve la concentration optimale en oxygène grâce aux lignes de champ, soit elle met de côté la magnétotaxie et utilise alors l’aérotaxie pour redescendre en profondeur dans la colonne sédimentaire où elle se trouve. Lorsqu'elle se sert de l’aérotaxie, puisqu’elle cherche un milieu micro-aérobie ou anaérobie strict, elle se fixe généralement à la limite oxique-anoxique, c’est pourquoi on retrouve un grand nombre de bactéries magnétotactiques à cet endroit.
Ainsi les bactéries magnétotactiques utilisent à la fois magnétotaxie et aérotaxie (on parle de magnéto-aérotaxie[4]) pour trouver un milieu de croissance optimal. La magnétotaxie augmente donc l’efficacité de l’aérotaxie, puisqu’une fois que la bactérie est dans une colonne sédimentaire, la recherche par aérotaxie ne se fait plus que suivant une seule dimension, contre trois dimensions pour les bactéries se servant uniquement de l’aérotaxie.
Applications
Aujourd'hui plusieurs applications des bactéries magnétotactiques sont envisageables, de par notamment leurs propriétés magnétiques, et ce dans différents domaines.
Médicale
En médecine, il serait tout d’abord possible d'utiliser les bactéries magnétotactiques comme agents de diagnostics.
La surface lipidique des bactéries magnétotactiques permet la fixation de bactériophages (ou phages) spécifiques. De plus, grâce aux magnétosomes et à leurs propriétés magnétiques, il est possible de guider les bactéries qui se déplacent grâce à leur flagelle. Ainsi, en produisant un champ magnétique et en le faisant varier au cours du temps (grâce par exemple à un ordinateur), il est possible de contrôler le déplacement des BMT, et ainsi de quadriller rapidement une zone définie. Au contact des bactéries cibles, les bactéries magnétotactiques vont s’y coller au moyen des phages. Il sera alors possible de détecter les bactéries cibles, en utilisant les cristaux magnétiques de la bactérie comme produits de contraste. L’avantage de cette détection est d’être plus précise et sensible que celles utilisées jusqu’à présent. De plus elle peut être appliquée aux êtres humains du fait de la non-toxicité de ces bactéries. Cette méthode ne serait cependant pas spécifique au ciblage des bactéries, tout dépendant de l’élément fixé aux BMT. Par exemple il serait possible d’utiliser cette technique pour repérer des cellules cancéreuses chez un individu de manière très précise.
Les bactéries magnétotactiques pourraient aussi être utilisées comme transporteurs dans le but d’améliorer l’efficacité de certains traitements, notamment des tumeurs. En effet il est difficile d’administrer très précisément un traitement dans toutes les parties du corps humain. En fixant à des bactéries magnétotactiques des microbilles contenant les agents thérapeutiques à utiliser, et en acheminant les bactéries magnétotactiques au moyen d'un champ magnétique permettant leur contrôle, il serait possible de délivrer le traitement de manière très ciblée. Cette utilisation des BMT permettrait de diminuer les doses à utiliser et la dispersion d’agents toxiques dans les autres tissus sains.
Comme agents de séparation
Les bactéries magnétotactiques peuvent être utilisées comme convoyeurs, et ce dans différentes situations, permettant la mise en place de techniques de séparation efficaces. Ceci est possible grâce à trois caractéristiques de ces BMT : leur ratio surface/volume, la possibilité de fixer à leur surface différents agents bioactifs, et la possibilité de les guider grâce à un champ magnétique. Ainsi par le biais d’un mécanisme identique à celui utilisé en médecine pour cibler des bactéries, il serait possible de fixer sur une substance précise des BMT grâce aux agents bioactifs ajoutés préalablement à leur surface, puis de les extraire grâce à nouveau au déplacement des bactéries par application d’un champ magnétique. Dans ce cadre, les bactéries magnétotactiques pourraient être utilisées par exemple pour la dépollution d’eaux, ou encore pour séparer différentes substances en présence.
Exobiologie
Les bactéries magnétotactiques sont avant tout célèbres pour être l'un des arguments en faveur de l'existence de vie en dehors de la Terre, sur Mars précisément. En effet, la NASA avait relevé la présence dans la météorite martienne ALH84001 de carbonates présentant une zonation chimique particulière[27], car ces derniers étaient semblables aux cristaux synthétisés par les bactéries magnétotactiques. L'hypothèse de présence de vie sur Mars s'est donc nourrie des similitudes en forme et en composition entre les cristaux produits par les bactéries magnétotactiques et les cristaux de carbonate prélevés sur la météorite.
Notes et références
- (en) « Ecology, Diversity, and Evolution of Magnetotactic Bacteria », sur Microbiology and Molecular Biology Reviews,
- (en) R.P. BLAKEMORE, « Magnetotactic bacteria », Science, no 190, , p. 377-379 (lire en ligne)
- PRADEL et al., « Les bactéries magnétotactiques et la biogénèse des magnétosomes »,
- D. A. BAZYLINSKI and R. B. FRANKEL, « Magnetosome formation in prokaryotes », Nature reviews,
- BAZYLINSKI, D. A. Structure and function of the bacterial magnetosome, ASM News, 1995, no 61, p. 337–343
- SPRING S. & BAZYLINSKI D. A., In : The Prokaryotes, New York : Springer, 2000
- (en) Dirk SCHÜLER, Magnetorecepetion and magnetosomes in Bacteria, Springer, , 319 p.
- FRANKEL R. B., BLAKEMORE R. P. & WOLFE R. S. Magnetite in freshwater magnetotactic bacteria, Science, 1979, no 203, p. 1355–1356
- BAZYLINSKI D. A., FRANKEL R. B. & JANNASCH H. W., Anaerobic production of magnetite by a marine magnetotactic bacterium, Nature, 1988, no 334, p. 518–519
- HEYWOOD B. R. et al., Controlled biosynthesis of greigite (Fe3S4) in magnetotactic bacteria, Naturwiss., 1990, no 77, p. 536–538
- DELONG E. F. et al., Multiple evolutionary origins of magnetotaxis in bacteria, Science, 1993, no 259, p. 803–806
- BAZYLINSKI D. A. et al., Fe3O4 and Fe3S4 in a bacterium, Nature, 1993, no 366, p. 218
- KAWAGUCHI R. et al., Phylogenetic analysis of a novel sulfatereducing magnetic bacterium, RS-1, demonstrates its membership of the δ-Proteobacteria, FEMS Microbiol. Lett., 1995, no 126, p. 277–282
- BLAKEMORE R. P., Magnetotactic bacteria, Annu. Rev. Microbiol., 1982, no 36, p. 217–238
- MANN S., SPARKS N. H. C. & BLAKEMORE R. P., Ultrastructure and characterization of anisotropic inclusions in magnetotactic bacteria, Proc. R. Soc. Lond. B Biol. Sci., 1987, no 231, p. 469–476
- BAZYLINSKI D. A. et al., Electron microscopic studies of magnetosomes in magnetotactic bacteria, Microsc. Res. Tech., 1994, no 27, p. 389-401
- BLAKEMORE R. P. et al., Microaerobic conditions are required for magnetite formation within Aquaspirillum magnetotacticum, Geomicrobiol. J., 1985, no 4, p. 53–71
- MANDERNACK. W. et al., Oxygen and iron isotope studies of magnetite produced by magnetotactic bacteria, Science, 1999, no 285, p. 1892–1896
- Jean PELMONT, Biodégradations et métabolismes : les bactéries pour les technologies de l'environnement, EDP Sciences, , 798 p.
- GUERINOT M. L., Microbial iron transport, Annu. Rev. Microbiol., 1994, no 48, p. 743–772
- NAKAMURA C. et al., Characterization of iron uptake in the magnetic bacterium Aquaspirillum sp. AMB-1, Appl. Biochem. Biotechnol., 1993, no 39–40, p. 169–176
- CALUGAY R. J. et al., Siderophore production by the magnetic bacterium Magnetospirillum magneticum AMB-1, FEMS Microbiol. Lett., 2003, no 218, p. 371–375
- GORBY Y. A., BEVERIDGE T. J. & BLAKEMORE R. P., Characterization of the bacterial magnetosome membrane, J. Bacteriol., 1988, no 170, p. 834–841
- (en) Matthieu Amor, Vincent Busigny, Pascale Louvat, Alexandre Gélabert, Pierre Cartigny, Mickaël Durand-Dubief, Georges Ona-Nguema, Edouard Alphandéry, Imène Chebbi et François Guyot, « Mass-dependent and -independent signature of Fe isotopes in magnetotactic bacteria », Science, American Association for the Advancement of Science, vol. 352, no 6286, , p. 705-708 (DOI https://dx.doi.org/10.1126/science.aad7632).
- Roland GALLAY, « Propriétés magnétiques : domaine de Weiss »
- Jacques BEAUCHAMP, « Cours de sédimentologie : les milieux de sédimentation »
- Philippe LABROT, « Le mystère ALH84001 »
Voir aussi
Articles connexes
Liens externes
- Rafal E. DUNIN-BORKOWSKI et al., « Magnetic Microstructure of Magnetotactic Bacteria by Electron Holography », AAAS, Science, (consulté le )
- Y. R. CHEMLA, H. L. GROSSMAN, T. S. LEE, J. CLARKE, M. ADAMKIEWICZ, and B. B. BUCHANAN, « A new study of bacterial motion : superconducting quantum interference device microscopy of magnetotactic bacteria », Biophysical Journal, volume 76,
- D.A. BAZYLINSKI and R.B. FRANKEL, « Magnetosome formation in prokaryotes », Nature Reviews, volume 2,
- PRADEL et al., « Les bactéries magnétotactiques et la biogénèse des magnétosomes »,
- Roland GALLAY, « Propriétés magnétiques : Domaine de Weiss » (consulté le )
- Jacques BEAUCHAMP, « Cours de sédimentologie : les milieux de sédimentation »
- N. MENGUY, « BioMinéralisations de nano-oxydes de fer par les bactéries magnétotactiques »
- « Recherche »
- (en) « Production, Modification and Bio-Applications of Magnetic Nanoparticles Gestated by Magnetotactic Bacteria », Nano Research,
- Jean PELMONT, Biodégradations et métabolismes : les bactéries pour les technologies de l'environnement, EDP Sciences, , 798 p.
- (en) Dirk SCHÜLER, Magnetorecepetion and magnetosomes in Bacteria, Springer, , 319 p.
- (en) Edmund BAUERLEIN, Biomineralization : from biology to biotechnology and medical applications, Wiley, , 337 p. (ISBN 3-527-31065-7, présentation en ligne)
- (en) Joel S. MILLER, Magnetism : molecules to materials, vol. 4, Wiley-VCH, , 337 p. (ISBN 3-527-30429-0, lire en ligne)
 Portail de la microbiologie
Portail de la microbiologie