Gymnophiona
Les Gymnophiona sont un ordre d'amphibiens vermiformes. Ils sont appelés en français apodes, cécilies ou gymnophiones. Le taxon Apoda est également utilisé pour désigner le groupe-couronne des espèces actuelles[1]. Il ne doit pas être confondu avec le genre Apoda qui désigne des lépidoptères nocturnes de la famille des Limacodidae.

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Amphibia |
| Sous-classe | Lissamphibia |
Familles de rang inférieur
Les gymnophiones ont été décrits par Johannes Peter Müller en 1832 et regroupent des amphibiens terrestres et aquatiques, auxquels des pattes atrophiées donnent l'aspect de vers de terre.
Description
Aspect général
Les gymnophiones sont des lissamphibiens fouisseurs, dépourvus de membres et de ceinture. Suivant les espèces, leur taille varie entre 7 cm et 1,4 m[2]. En général, leur taille varie entre 20 cm et 50 cm (Taylor, 1968). Ils sont généralement carnivores.
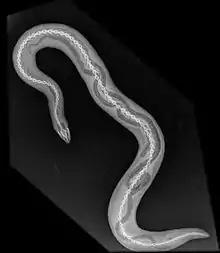
Leur corps est cylindrique et allongé. Il est marqué sur toute son étendue d’une succession d’anneaux et sillons transversaux. Le tronc est terminé par une courte queue de même diamètre que le reste du corps. Chaque anneau correspond à une vertèbre, dont le nombre peut atteindre 300.
Chez plusieurs genres tels que Typhlonectes il n’y a pas d’anneaux mais des plis. Plusieurs familles possèdent une véritable queue correspondant à plusieurs vertèbres situées en arrière de l’orifice cloacal. Chez d'autres, il n’y a pas de queue car pas de vertèbres postérieures au cloaque. Chez d’autres enfin, un petit appendice, l'écu (shield de Taylor, 1968) est postérieur au cloaque. L’ouverture cloacale peut avoir une forme différente selon le sexe mais il est souvent difficile de déterminer extérieurement le sexe.
La peau
La peau est épaisse généralement grisâtre et peut être recouverte d’écailles chez les espèces appartenant aux familles considérées comme les moins évoluées (Rhinatrematidae, Ichthyophiidae). Ces écailles ressemblent à celles des poissons ostéichthyens et recouvrent tout le corps. Chez d’autres espèces, les écailles couvrent seulement la partie postérieure du corps. Dans ce cas les écailles les plus antérieures sont plus petites que les autres et dispersées dans la peau. C’est le cas de Geotrypetes seraphini, par exemple. Aucune écaille n’a été observée dans la peau des formes considérées comme les plus évoluées (Typhlonectidae) même si quelques écailles dégénérées ont été décrites (Wake et Nygren, 1987). Leur peau est visqueuse, aux sécrétions toxiques.
Le squelette
Le crâne est composé de plus d’os que chez les autres Lissamphibiens. La mâchoire inférieure est constituée de deux moitiés réunies par du tissu conjonctif. Une structure rétro-articulaire est observée en arrière de chaque mâchoire inférieure. Sur cette structure anatomique s’insère une puissante musculature impliquée dans la mastication et la fermeture de la bouche après la capture des proies.
Les vertèbres sont de type primitif et permettent seulement un déplacement horizontal (Lawson 1963, 1966a, Taylor 1977, Renous et Gasc 1986a, b). Ce sont des vertèbres dites amphicèles, rappelant celles des poissons osseux. Le centre de la vertèbre comporte un centrum, cicatrice du passage de la chorde dorsale embryonnaire. Les trois premières vertèbres sont plus grandes que les autres et elles sont très mobiles. Elles constituent le collier que l’on peut comparer au cou des vertébrés amniotes (Lescure and Renous 1992).
Les Gymnophiones ne possèdent ni membres ni ceintures. Mais chez le primitif Ichthyophis glutinosus, des traces de ceintures régressées ont été observées, et pendant le développement embryonnaire, des blastèmes de membres se développent mais dégénèrent à la métamorphose (Renous et al., 1997). La perte des membres est donc secondaire.
Le cerveau
Le cerveau des Gymnophiones adultes ressemble à celui des autres amphibiens mais présente à la fois des caractéristiques trouvées chez les vertébrés amniotes et des caractéristiques propres, telles que l’allongement et une flexion entre le diencéphale et le mésencéphale. Les nerfs olfactifs sont particulièrement développés et deux bulbes olfactifs accessoires associés aux organes voméronasaux sont observés en position antérieure.
Les organes des sens

Les yeux sont de petite taille, recouverts par la peau qui peut être plus ou moins épaisse et transparente à ce niveau. Chez Scolecomorphus, ils sont recouverts par les os crâniens. Le cristallin n’est pas totalement lamellaire car il comporte des cellules nucléées, la rétine comporte seulement des cônes. Les nerfs optiques sont réduits, la musculature oculomotrice est combinée aux tentacules. L’aire céphalique qui correspond aux muscles oculomoteurs est également réduite (Wake 1985, Himstedt et Manteuffel 1985, Brun et Exbrayat 2007). Chez les Gymnophiones, un pigment vert caractéristique des autres Amphibiens actuels n’est pas retrouvé (Rage 1985). Cette régression des yeux représente incontestablement une adaptation à la vie fouisseuse.
Une paire de tentacules a été décrite chez tous les Gymnophiones. Chacun d’entre eux est contenu dans un sac qui enveloppe également l’œil correspondant. Chaque tentacule est extrudé à l’extrémité du museau, entre la narine et l’œil à l’aide d’une musculature (Billo and Wake 1987), lubrifié par les glandes de Harder (glandes lacrymales). Chaque tentacule est relié à un organe voméronasal. Les tentacules sont en permanence animés d’un mouvement de va-et-vient par lequel ils captent les molécules qui sont analysées par les organes voméronasaux. Il s’agit d’une adaptation à la vie fouisseuse qui permet à l'animal de s’orienter dans le tunnel qu’il est en train de creuser alors que les narines et la bouche sont hermétiquement fermées.
Des organes de la ligne latérale, caractéristiques des vertébrés aquatiques, sont observés chez les larves aquatiques des Gymnophiones mais pas chez les larves intra-utérines. On en trouve également chez les adultes de certaines espèces aquatiques. Ces organes sont répartis de part et d’autre du corps (neuromastes) et sur le museau (organes ampoulaires).
Le tube digestif
Les dents des adultes sont caractéristiques des amphibiens avec un pédicelle sur lequel une partie, la couronne, est insérée par l’intermédiaire d’une couche de tissu conjonctif fibreux. Chez les larves intra-utérines, une dentition fœtale particulière permet à l’animal de se nourrir des sécrétions de la paroi utérine. Ces dents fœtales sont très différentes de celles des adultes par leur origine, leur structure et leur répartition.
L’œsophage, l’estomac et l’intestin ne présentent pas de caractères particuliers. Le foie est allongé, segmenté, situé à droite dans la cavité générale. Il est recouvert, même chez l’adulte, d’une couche périphérique de tissu hématopoïétique (Paillot et al. 1997a, b).
L’appareil respiratoire
Chez les adultes, l’appareil respiratoire est constitué d’une trachée qui peut comporter des alvéoles, d’une paire de poumons dont le développement est variable selon les espèces : les deux poumons peuvent être très longs, développés tous deux, ils peuvent être plus courts mais développés de manière identique, l’un peut être plus court ou même inexistant. Atretochoana ne possède aucun poumon et respire par la cavité buccale. Les larves possèdent une paire de branchies triradiées. Chez Typhlonectes, ces dernières sont transformées en organes vésiculeux qui contribueront à constituer une sorte de placenta au cours du développement embryonnaire.
Le cœur
Le cœur des Gymnophiones est compose d’un cône artériel, d’un ventricule comportant un grand pilier musculeux central, de deux oreillettes séparées par une cloison musculeuse pouvant être complète ou fenêtrée, et d’un sinus veineux.
Le système immunitaire
La structure du thymus est classique. Chez Typhlonectes, des corpuscules de Hassal, structures caractéristiques des amniotes sont observés mais pas chez d’autres genres (Ichthyophis par exemple) (Bleyzac et al. 2005). La rate ressemble à celle des autres amphibiens. Comme indiqué plus haut, le foie est entouré d’une couche de tissu hématopoïétique.
Le système excréteur
Il est constitué d’une paire de reins allongés et d’une vessie. Chez les espèces aquatiques, les glomérules sont de grande taille par rapport aux espèces terrestres. Chez les formes considérées comme primitives, une paire de pronéphros persiste chez les adultes, ce qui a fait parfois considérer les reins des Gymnophiones comme le modèle ancestral (Brauer 1902). Mais chez d’autres espèces, chaque pronéphros régresse à la métamorphose.
Comme chez les autres amphibiens, la vessie est une expansion de la paroi du cloaque sans connexion directe avec les canaux de Wolff (uretères).
Les organes endocriniens
L’épiphyse a été étudiée chez les adultes et au cours du développement embryonnaire de Typhlonectes compressicauda, chez qui le complexe pinéal est une structure en forme de petit pois provenant du diencéphale. Aucun organe parapinéal n'est observé chez les Gymnphiones (ainsi que chez les amniotes), contrairement aux autres amphibiens (anoures et urodèles).
La glande thyroïde ne présente aucune particularité notable, pas plus que les glandes parathyroïdes.
Les glandes interrénales (équivalents des surrénales) sont situées dans la partie antérieure des reins. Elles contiennent des cellules adrénocorticales et des cellules adrénales soumises à des variations de leur cycle sécrétoire au cours des saisons.
L'adénohypophyse des Gymnophiones est aplatie et contient plusieurs types cellulaires, comme chez les autres vertébrés : cellules lactotropes sécrétant de la prolactine, cellules gonaditropes sécrétant de l'hormone lutéinisante (LH) et de l'hormone folliculo-stimulante (FSH), cellules somatotropes avec de l'hormone de croissance (GH), corticotropes avec de l’hormone corticotrope (ACTH) et thyréotrope avec de la thyréostimuline (TSH). Tous ces types cellulaires sont soumis à des variations cycliques au cours des cycles de reproduction et des cycles saisonniers
Les appareils génitaux et la reproduction
L’appareil génital mâle a été étudié chez de nombreuses espèces (Wake 1968, 1970, Exbrayat and Estabel 2006, Akbarsha et al. 2006, Smita et al. 2006, Scheltinga and Jamieson 2006, Measey et al. 2008). Il comporte une paire de testicules multilobés associés à des corps adipeux segmentés, une paire de canaux de Müller et un cloaque érectile permettant la fécondation interne. La segmentation des testicules est plus régulière chez les genres les plus ancestraux (Ichthyophis, Rhinatrema) que chez les autres (Typhlonectes).
Chaque lobe testiculaire est composé de plusieurs lobules dans lesquels se déroule la spermatogenèse. Les cellules germinales sont regroupées en séries isogéniques, c'est-à-dire provenant de la même spermatogonie, qui prennent l’aspect d’une structure sphérique se déplaçant de la périphérie vers le centre entre des cellules de Sertoli géantes, l’ensemble donnant aux testicules de Gynmophiones un aspect unique chez les vertébrés.
Tous les Gymnophiones mâles possèdent une paire de canaux de Müller qui ne dégénèrent pas au cours du développement embryonnaire, comme c’est la règle générale chez les vertébrés mâles. Ils sont au contraire transformés en véritables glandes dont le cycle de sécrétion est étroitement lié au cycle de reproduction (Wake 1981, Exbrayat 1985, Akbarsha et al. 2006). Ces organes sont les équivalents de la prostate.
Tous les Gymnophiones pratiquent la fécondation interne. Pour cela, la partie postérieure du cloaque mâle est érectile et se retourne en doigt de gant au moment de la copulation, donnant un phallodeum de forme et de taille diverses selon les espèces, permettant l’intromission dans le cloaque de la femelle (Tonutti 1931, Exbrayat 1991, 1997).
L’appareil génital femelle a également été étudié chez plusieurs espèces (Wake 1968, 1970, Exbrayat and Estabel 2006, Exbrayat 2006c, Raquet et al. 2006). Les ovaires sont allongés et associés à des corps adipeux segmentés. Après l’ovulation, le follicule, vidé de son ovocyte, se transforme en corps jaune qui évoluera en véritable glande endocrinienne vascularisée, sécrétrice de progestérone, qui sera maintenue en place pendant tout le temps où l’œuf restera dans les oviductes. Chez les espèces vivipares, les corps jaunes persisteront pendant toute la durée de la gestation et seront le siège de variations structurales.
Les œufs sont contenus dans les oviductes. Après l’ovulation, les ovocytes sont conduits à la partie antérieure de l’oviducte et c’est à ce niveau que la fécondation est observée. Chez les espèces ovipares, les œufs et embryons sont entourés d’une gangue muqueuse sécrétée par les glandes tubaires. Lors de la ponte, les œufs embryonnés seront reliés les uns aux autres par un cordon muqueux.
Chez les espèces vivipares, la partie postérieure des oviductes est transformée en utérus. La paroi utérine sécrète des substances glyco-lipo-protéiques qui serviront de nourriture aux embryons, qui pourront également arracher les cellules épithéliales de la paroi utérine. Les embryons pratiquent également l’adelphophagie, c’est-à-dire qu’ils dévorent les embryons morts ou peut-être moins développés. Chez Typhlonectes compressicauda, une placentation qui implique les branchies et la paroi utérine est observée à la fin du développement (Hraoui-Bloquet, 1995 ; Exbrayat and Hraoui-Bloquet, 2006).
Le développement et la métamorphose
Le développement embryonnaire des Céciliens a été peu étudié. Les œufs sont télolécithes, c'est-à-dire qu'ils contiennent une importante quantité de vitellus située au pôle végétatif. Le développement concerne un blastidisque situé au pôle animal. L’embryon se développe autour de sa masse de vitellus. Chez les espèces vivipares, au moment de l’éclosion intra-utérine, il est équipé d’une denture fœtale qui lui permet de racler la paroi utérine. Cette denture disparaîtra avant la naissance. La métamorphose peut être lente, chez les espèces ovipares dont les larves sont aquatiques (10 mois chez Ichthyophis), ou rapide, chez les espèces vivipares (estimée à 2 mois chez Typhlonectes). Elle est caractérisée notamment par la disparition des branchies, sauf chez Typhlonectes, par la régression des bourgeons des membres, par la disparition du pronéphros chez certaines espèces, par la mise en place de la structure définitive de la peau. Pendant la croissance des petits, la mère produit tous les trois jours une nouvelle peau grasse que les petits dévorent tous ensemble en quelques minutes[3]. Les modes de reproduction sont variables. La fécondation est toujours interne. De nombreuses espèces sont ovipares (Ichthyophis), parfois au développement direct[pas clair]. Une sorte de placentation impliquant les branchies des larves intra-utérines et la paroi de l’oviducte est observée chez Typhlonectes compressicauda (Taylor, 1968, Hraoui-Bloquet et Exbrayat, 1995, Wake, 1977). Les apodes forment un petit groupe très spécialisé, éloigné du type amphibien classique. Ces amphibiens ne paraissent pas subir de métamorphose, car elle ne se signale que par la perte des branchies externes et s’effectue très tôt dans l’œuf.
Ces espèces hibernent tous les ans au même endroit à 1 ou 2 cm près.[réf. nécessaire] Certaines convoitent les œufs des autres pour se nourrir[pas clair].
Distribution et habitat

Ce sont des animaux terrestres qui vivent dans l'humus, la boue des marécages en forêt tropicale, voire en eau libre.
Les espèces de cet ordre se rencontrent en Amérique du Sud, en Amérique centrale, en Afrique de l'Ouest, en Afrique centrale, en Afrique de l'Est, en Asie du Sud et en Asie du Sud-Est. Elles sont absentes de l'Australie, de Madagascar et des Antilles.
Nomenclature et systématique
Selon Amphibian Species of the World (16 septembre 2014)[4] :
- Caeciliidae Rafinesque, 1814
- Chikilidae Kamei, San Mauro, Gower, Van Bocxlaer, Sherratt, Thomas, Babu, Bossuyt, Wilkinson & Biju, 2012
- Dermophiidae Taylor, 1969
- Herpelidae Laurent, 1984
- Ichthyophiidae Taylor, 1968
- Indotyphlidae Lescure, Renous & Gasc, 1986
- Rhinatrematidae Nussbaum, 1977
- Scolecomorphidae Taylor, 1969
- Siphonopidae Bonaparte, 1850
- Typhlonectidae Taylor, 1968
et les espèces fossiles basales :
- † Eocaecilia micropodia Jenkins & Walsh, 1993
- † Rubricacaecilia monbaroni Evans & Sigogneau-Russell, 2001
- Caecilia pulchraserrana (Caeciliidae)

 Herpele squalostoma (Herpelidae, femelle et petits)
Herpele squalostoma (Herpelidae, femelle et petits)
 Epicrionops sp. (Rhinatrematidae)
Epicrionops sp. (Rhinatrematidae)
 Reconstitution d'un Eocaecilia fossile
Reconstitution d'un Eocaecilia fossile
Par le passé, les gymnophiones ont été séparées en trois groupes selon leur niveau d'« évolution » : les cécilies primitives pour les familles des Rhinatrematidae et des Ichthyophiidae, les cécilies moyennement évoluées pour le genre Uraeotyphlus (Ichthyophiidae parfois dans les Uraeotyphlidae) et les cécilies très évoluées pour la famille des Caeciliidae et les anciennes familles des Scolecomorphidae et des Typhlonectidae[5].
Phylogénie externe
Phylogénie des ordres d'amphibiens modernes d'après Marjanovic & Laurin (2007)[6] :
| Lissamphibia |
| ||||||||||||||||||
Phylogénie interne
Phylogénie des gymnophiones d'après Wilkinson et al. (2011)[7] et San Mauro et al. (2014)[8],[9] :
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Notes et références
- http://fossilworks.org/bridge.pl?a=taxonInfo&taxon_no=252779
- Cécilie
- « Vivre de sang froid »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?), Documentaire de Scott Alexander, Épisode 2 sur 5, PLANETE TV, 17:30 Lundi 8 novembre 2010.
- Amphibian Species of the World, consulté le 16 septembre 2014
- Les Cécilies, Amphibiens mystérieux, 2005. Musée d'histoire naturelle Fribourg, Suisse (Swiss Caecilian Research Berne). Textes et illustrations de l'exposition temporaire du 10 septembre 2005 au 15 janvier 2006 Lire le document pdf.
- Marjanović, D. & Laurin, M. (2007) Fossils, Molecules, Divergence Times, and the Origin of Lissamphibians . Systematic Biology 56, 369-388.
- (en) M. Wilkinson, San Mauro, D., Sherratt, E. et Gower, D.J., « A nine-family classification of caecilians (Amphibia: Gymnophiona) », Zootaxa, vol. 2874, , p. 41–64 (lire en ligne)
- (en) Pyron, R.A. et Wiens, J.J., « A large-scale phylogeny of Amphibia including over 2800 species, and a revised classification of extant frogs, salamanders, and caecilians », Molecular Phylogenetics and Evolution, vol. 73, no 2, , p. 177–189 (DOI 10.1016/j.ympev.2011.06.012)
- (en) D. San Mauro, Gower, D. J., Müller, H., Loader, S. P., Zardoya, R., Nussbaum, R. A. et Wilkinson, M., « Life-history evolution and mitogenomic phylogeny of caecilian amphibians », Molecular Phylogenetics and Evolution, vol. 61, , p. 543–583 (PMID 24480323, DOI 10.1016/j.ympev.2014.01.009)
Bibliographie
- Müller, 1832 : Beiträge zur Anatomie und Naturgeschichte der Amphibien. I. Ueber die natürliche Eintheilung der Amphibien. Zeitschrift für Physiologie, vol. 4, p. 190-275.
- Delsol, Flatin & Exbrayat 1995 : Le tube digestif des Amphibiens adultes. Traité de Zoologie, tome XIV, fasc. I A. Grassé & Delsol (eds), Masson, Paris, p. 497-508.
- Dünker, Wake & Olson, 2000 : Embryonic and larval development in the Caecilian Ichthyophis kohtaoensis. Journal of morphology, vol. 243, p. 3-34 (texte intégral).
- Exbrayat, 2000 : Les Gymnophiones, ces curieux amphibians. Société Nouvelle des éditions Boubée, Paris.
- Exbrayat, 2006 : Reproductive biology and phylogeny of Gymnophiona (Caecilians). Vol 5 of series, Jamieson GMB (ed). Science Publishers, Enfield Jersey Plymouth.
- Himstedt, 1996 : Die Blindwühlen. Westarp-Wissenschaft Magdeburg.
- Nussbaum & Wilkinson, 1989 : On the classification and phylogeny of Caecilians (Amphibia: Gymnophiona), a critical review. Herpetological Monographs, vol. 3, p. 1-42.
- Rage, 1985 : Origine et phylogénie des Amphibiens. Bulletin de la Société Herpétologique de France, vol. 34, p. 1-19.
- Sakai, Billo & Kriz, 1986 : The structural organization of the kidney of Typhlonectes compressicaudus (Amphibia, Gymnophiona). Anatomy and Embryology, vol. 174, p. 243-252.
- Sammouri, Renous, Exbrayat & Lescure, 1990 : Développement embryonnaire de Typhlonectes compressicaudus (Amphibia Gymnophiona). Annales des Sciences Naturelles, Zoologie, sér. 13, vol. 11, p. 135-163.
- Sarasin & Sarasin, 1887-1890 : Ergebnisse Naturwissenschaftlicher Forschungen auf Ceylon. Zur Entwicklungsgeschichte und Anatomie der Ceylonischen Blindwuhle Ichthyophis glutinosus. C.W. Kreidel’s Verlag, Wiesbaden.
- Seshachar, 1936 : The spermatogenesis of Ichthyophis glutinosus (Linn.) I. The spermatogonia and their division. Zeitschrift für Zellforschung und mikroskopische Anatomie, vol. 24, p. 662-706.
- Taylor, 1968 : The Caecilians of the world. A taxonomic review. University Kansas Press, Lawrence, Kansas, USA.
- Tonutti, 1931 : Beitrag zur Kenntnis der Gymnophionen. XV Das genital System. Morphologisches Jahrbuch, vol. 70, p. 101-130.
- Wake, 1968 : Evolutionary morphology of the Caecilian urogenital system. Part I : the gonads and fat bodies. Journal of Morphology, vol. 126, p. 291-332.
- Wake, 1970 : Evolutionary morphology of the caecilian urogenital system. Part II : the kidneys and urogenital ducts. Acta Anatomica, vol. 75, p. 321-358.
Liens externes
- (en) Référence Amphibian Species of the World : Gymnophiona Müller, 1832 (consulté le )
- (en) Référence Animal Diversity Web : Gymnophiona (consulté le )
- (en) Référence Catalogue of Life : Gymnophiona (consulté le )
- (en) Référence NCBI : Gymnophiona (taxons inclus) (consulté le )
- (en) Référence Tree of Life Web Project : Gymnophiona (consulté le )
- (en) Référence Paleobiology Database : Gymnophiona Rafinesque 1814 (consulté le )
- (en) Référence uBio : Gymnophiona Müller, 1831 (consulté le )
- (en) Référence UICN : taxon Gymnophiona (consulté le )
- (en) Référence Wild Herps : photographies de Gymnophiona (consulté le )
 Portail de l’herpétologie
Portail de l’herpétologie