Contraintes phylogénétiques et adaptation
La notion de contrainte phylogénétique renvoie aux concepts de phylogénie[1] en s'appuyant sur les concepts[2] de sélection naturelle, d’adaptation et d’évolution.
Une adaptation, produit de la sélection naturelle, peut donc être influencée par des contraintes phylogénétiques, c’est-à-dire par l’histoire évolutive de l’individu. L’adaptation se construit donc sur histoire préexistante.
On appelle donc contraintes phylogénétiques tout mécanisme qui par son action empêche l'adaptation optimale. Ce concept a été fortement développé en écologie dans les années 80 et 90 sous l'influence des publications[3] de Stephen Jay Gould.
Contraintes phylogénétiques
Définition
On peut définir une contrainte phylogénétique sur l'évolution d'un caractère d'un individu, comme tout mécanisme lié à l'histoire évolutive et à la phylogénie de cet individu qui empêche l'intervention de la sélection naturelle et l'adaptation envisageable de ce caractère.
Le concept de contrainte phylogénétique est souvent utilisé pour expliquer pourquoi un trait ne s’est pas développé de façon optimale, ou pourquoi il n’a pas été perdu au cours de l’évolution.
Historique
En 1979, Stephen Jay Gould & Richard C. Lewontin publient le célèbre article The spandrels of San Marco and the Panglossian paradigm a critique of the adaptationist programme[3]. Ils critiquent le fait que dans l’approche adaptationniste de nombreuses études, chaque trait est pris indépendamment des autres, or les organismes doivent être analysés en tant qu’ensemble intégré. C'est à partir de cet article provocateur que le concept de contraintes phylogénétiques va évoluer et être considéré dans les processus d'adaptation.
De nombreuses publications ont suivi celle de Gould dans les années 80 et 90.
Dans une publication en 1985, John Maynard Smith définit les contraintes développementales comme étant des « biais dans la production ou la variabilité des différents phénotypes, génétiquement possibles, induit par les mécanismes se produisant lors du développement provoqués par la structure, la composition, ou la dynamique du système de développement »[réf. nécessaire].
Pour Derrickson & Ricklefs[4] (1988), il s'agirait de tout changement dans l'interaction entre un phénotype et son environnement, qui puisse être relié aux mécanismes qui ont conduit à la diversification d'un des taxons auquel cet organisme appartient. Dans leur publication ils font la différence entre des effets phylogénétiques et des contraintes phylogénétiques. Un effet phylogénétique est seulement l’expression de la tendance des espèces à être similaires parce qu’ils possèdent une histoire commune. Alors qu’une contrainte phylogénétique est l’effet de cette histoire commune sur la diversification des changements au sein d’un clade.
Mary C. McKitrick[5] (1993) reprend cette dernière définition en se concentrant sur les conséquences plutôt que sur les causes de ces contraintes : une contrainte phylogénétique est un «quelconque résultat ou composante de l'histoire phylogénétique d'une lignée qui prévient et anticipe le cours de l'évolution de cette lignée». Cette définition rejoint celle donnée pour le terme exaptation par Gould et Vrba en 1982 dans Exaptation-a missing term in the science of form[6].
En 1993, Ligon[7] émet l’hypothèse qu’une adaptation survenue à un certain moment de l’évolution d’un groupe taxonomique peut être la cause d’une contrainte phylogénétique sur un taxon actuel, appartenant à ce groupe.
D'après D. Brent Burt[8] (2001), on peut mettre en évidence une contrainte phylogénétique si on détecte un avantage sélectif au changement d'un caractère puis si on détecte des mécanismes empêchant ce caractère d’évoluer, à contrario de ce qui devrait se passer si l’adaptation avait lieu.
L’importance donnée aux contraintes phylogénétiques sur l’évolution des espèces n’a cessé de s’accroître depuis l’article de Gould. Il est devenu clair que les espèces possèdent des traits loin d’être parfaitement adapté à leur besoin individuel, ce qui pourrait être expliqué par des contraintes phylogénétiques.
Les différents niveaux de contraintes
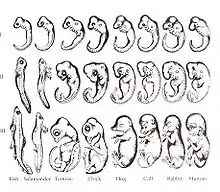
Il existe différents types de contraintes phylogénétiques :
- Développementale : d'après John Maynard Smith[9] (1985) il s'agit d'un « biais dans la variabilité ou la production des différents phénotypes, génétiquement possibles, induit par les mécanismes se produisant lors du développement provoqués par la structure, la composition, ou la dynamique du système de développement ». Il y a donc une grande influence des contraintes développementales sur les possibles variations des traits de l’individu. Par exemple : il existe une similitude dans les stades embryonnaires précoces chez les Vertébrés, ce qui décrit une parenté évolutive. Ils sont issus d’un même ancêtre commun et partagent des traits développementaux similaires. Plus tard, il y aura une différenciation due à l’adaptation des différents clades à chacune de leur niches.
- Mécanique : Par exemple chez l'homme, l’ensemble épaule–bras résulte d’une contrainte mécanique qui sert à faire tenir le bras par rapport à l'épaule mais surtout par rapport à l’ensemble du corps, il ne s’agit donc pas de l’adaptation d'un seul trait mais de l’intégration de ce trait par rapport à l’ensemble du corps.
- Morphologique : La locomotion chez les kangourous (Macropus rufus) par bond, sera une contrainte favorisée par rapport à la bipédie classique. Cette contrainte est à différentier d’une maladaptation. Cependant, la bipédie n’est pas impossible comme le montre le déplacement chez les dendrolagues (Dendrolagus matschiei) qui possèdent le même ancêtre commun que le kangourou.
- Génétique : Une contrainte génétique est un mécanisme empêchant la sélection des phénotypes délétères, elle est due au fait que pour un allèle donné, la fitness des individus hétérozygotes est supérieure à celle des deux autres phénotypes. Par exemple : Mac Gregor et Horer[10] (1980) équilibre létal du triton (Triturus cristatus). Pour le chromosome 1, il existe deux chromosomes d'aspect différent 1A et 1B. Les trois génotypes possibles (1A/1A), (1B/1A) et (1B/1B) devraient apparaître dans la population, pourtant seulement le génotype hétérozygote apparaît, les deux homozygotes ayant un effet létal. La fitness de l’hétérozygote est dans cet exemple infiniment supérieure à celle des homozygotes, ce qui confirme bien qu'il s'agit d'une contrainte génétique.
- Physiologique : Pour une espèce comme la mésange charbonnière (Parus major), qui, comme la plupart des animaux, ne peut s’approprier que les ressources alimentaires dans un périmètre restreint correspondant à sa morphologie et son état physiologique. En prenant en compte les facteurs environnementaux (prédation) et la disponibilité en proie, le rapport entre l'alimentation de son prédateur et la taille de sa proie constitue une contrainte physiologique pour cette espèce.
Adaptation
Définitions

L’adaptation peut être désignée à la fois comme un trait phénotypique fixé ou stabilisé dans une population par la sélection naturelle, ou comme le processus amenant à des ajustements des traits de l’organisme à l’environnement sous l’effet de la sélection naturelle. Elle offre un gain pour la fitness de l’individu (c’est-à-dire pour la survie et/ou la reproduction).
Une adaptation résulte d’une modification du génome par une mutation due au hasard. La dispersion de cette adaptation dans la population va dépendre de sa valeur adaptative par rapport à la forme d’origine mais s’il reste une composante stochastique.
L’hypothèse adaptative reposerait sur la sélection de l’adaptation « parfaite ». Or le spandrelisme des contraintes biologiques de Stephen Jay Gould et Richard C. Lewontin soulève le problème d’une vision beaucoup trop idéaliste de la sélection naturelle. Ils s’opposent directement à la vision Panglossienne du programme adaptatif : ce ne sont pas toujours des adaptations parfaites. Pour ce faire ils utilisent une métaphore pour expliquer ce problème : les spandrels de la cathédrale San Marco[3]. Ces éléments architecturaux qui paraissent parfaitement adaptés et leurs ornementations pensés pour décorer la cathédrale ne résultent finalement que de contraintes architecturales et mécaniques. Ils sont présents pour faire tenir le dôme, le dôme impose l’utilisation des spandrels. Ils résultent de contraintes, leur ornementation est secondaire. Finalement, Gould et Lewontin concluent que tout ce qui est sélectionné n’est pas forcément une adaptatif.
Les adaptations peuvent alors résulter d’une contrainte phylogénétique et leur intérêt de sélection est finalement secondaire.
Ils postulent que dans cette vision idéaliste on ignore d’autres phénomènes comme la dérive génétique, les relations d’allométrie (relations entre la taille d’un organe et l’ensemble du corps) ou l’intégration d’un Bauplan (plan d'organisation) contraint par l’origine phylogénétique de l’espèce, en effet, toute espèce « actuelle » est l’héritière de structures qui adaptaient ses ancêtres à d’autres environnements.
Les deux principaux points mis en avant dans leur critique sont les suivants :
- On doit prendre compte tous les traits de l’organisme et non les prendre indépendamment des autres (bannir le réductionnisme) : il ne faut pas oublier que la sélection s’adresse à l’organisme dans sa globalité, et non focalisée sur un seul trait;
- La sélection naturelle n’est pas la seule hypothèse pour optimiser l’adaptation : par exemple, par simple dérive génétique, un allèle rare peut être perdu, or cet allèle était peut-être adapté au milieu ; une mutation dite défavorable peut être fixée.
Confusions à éviter
Une convergence évolutive concerne un trait qui présente une même forme d’adaptation au sein de deux espèces différentes soumises aux mêmes contraintes environnementales. Il s’agit deux évolutions indépendantes qui ne proviennent pas d’un ancêtre commun.
Exemple : Le cas des mammifères marins et des poissons, leur aspect extérieur est semblable alors que ce sont deux groupes très différents. Il en est de même pour les yeux complexes des vertébrés et des céphalopodes, ces deux taxons ont évolué vers des systèmes optiques similaires, pourtant leur ancêtre commun n’en possédait pas.
Pour un même génotype, on retrouve un phénotype différent selon l’habitat. C'est un compromis associé à un organisme sachant d’adapter avec une certaine variabilité : possession d’une gamme de solution selon les conditions.
La plasticité phénotypique tend à maximiser la fitness dans un environnement rencontré. Elle est donc réversible contrairement à une adaptation (rarement réversible).
- Exaptation
Stephen Jay Gould et Elisabeth Vrba (en)[6] dans un article de 1981 parlent d'exaptation. C'est-à-dire, une adaptation actuelle n'a pas forcément la même fonction que l'adaptation initiale. Finalement c'est l'évolution de la fonction initiale d'une adaptation vers une autre fonction. Des caractéristiques plus complexes peuvent survenir après la fixation d'une adaptation a priori simple.
Ils prennent l'exemple des oiseaux, les plumes peuvent être vues comme une adaptation pour la thermorégulation et plus tard vu comme une exaptation pour attraper des insectes. Puis le développement des plumes plus larges et leur arrangement sur l'aile sont devenus une nouvelle adaptation pour attraper des insectes.
Innovation clé
Les succès évolutifs d’un groupe taxonomique peuvent être expliqués par la notion d'« Innovation clé »[11], c’est-à-dire l’apparition d’un nouveau trait phénotypique qui permet la colonisation d’une nouvelle niche écologique. De plus, ce phénomène décrit la capacité de diversification rapide, souvent non réversible, en réponse à de nouvelles contraintes environnementales, de certains taxons.
La diversité et le nombre important d’espèces dans certains groupes phylogénétiques, par rapport aux groupes frères résultent du concept de Key Innovation.
Il existe plusieurs exemples afin d’illustrer ce concept. L’un d’entre eux est la viviparité chez certaines espèces de vipères[12]. En effet, une analyse phylogénétique montre que des espèces vivipares de vipères ne sont pas plus répandues dans les climats plus froids mais ont tendance à évoluer dans ces milieux-là. Il a été démontré que les femelles de cette espèce peuvent maintenir une température d’incubation plus haute et plus stable à travers une thermorégulation comportementale (exposition au soleil) donc la viviparité est une adaptation au froid. Ainsi cette key innovation permet une diversification des vipères vivipares et une augmentation de leur fitness.
De la même manière, l’écholocation chez les chauves-souris (microchiroptères uniquement) montre qu’à petite échelle, il y a une modulation de ce comportement en fonction d’un nouvel environnement et de leur alimentation. En prenant en compte l’existence d’un ancêtre diurne et frugivore, les données phylogénétiques ont permis d’établir que l’écholocation est une adaptation qui permettrait un élargissement de leur niche réalisée à une niche moins ou peu exploitée (capture d’oiseaux migrateurs nocturnes en vol par exemple) et donc la colonisation d’une nouvelle niche. Ainsi, les deux key innovation décrites sont l’adaptation à l’orientation spatiale et le fourragement.
Radiation évolutive
La radiation évolutive est un phénomène où on l'on observe une augmentation d’un nombre d'espèces , dans un court laps de temps. Cette évolution rapide expliquerait l'existence de ces clades qui possèdent une grande diversité d'espèces. Ce phénomène est souvent observé après une extinction d'espèce ou plus simplement un changement environnemental favorisant la multiplication de certaines espèces en dépit d'autres.
On parle ensuite de radiation adaptative si ces nouvelles espèces colonisent de nouvelles niches non disponibles à l'origine.
Méthode comparative[13]
L'étude comparative est utilisée afin de vérifier l’origine d’un nouveau trait, s’il est issu d’une contrainte phylogénétique ou d’une adaptation due à la sélection naturelle. On essaye de mettre en évidence une corrélation entre la structure du trait et l’environnement dans plusieurs lignées en tenant compte de leur ancêtre commun. La phylogénie rend les résultats non indépendants. On compare la plupart du temps deux traits phénotypiques aux travers de plusieurs clades, ou un même trait phénotypique avec un environnement variable.
Cette méthode comporte certaines difficultés, il faut qu’il y ait suffisamment d’hypothèses phylogénétiques incluant assez de groupes non liés avec la même fonction (afin de pouvoir les comparer entre elles) et pouvoir faire des hypothèses de comparabilité du trait pour les différents groupes (sur le degré d’analogie du trait lui-même dans l’environnement où l’organisme vit).
Comme dit précédemment Burt[8], propose une méthode pour mettre en évidence une contrainte phylogénétique en détectant un avantage sélectif s'il y a un changement au niveau du trait, puis en détectant des mécanismes empêchant ce trait d’évoluer.
Bibliographie
- François Ramade, Dictionnaire encyclopédique de l'Écologie et des Sciences de l'Environnement, Dunod, 2002 (ISBN 2100066706)
- Lecointe et le Guyader, Classification phylogénétique du vivant, Belin, 2009 (3e ed.)
- Ridley, Évolution Biologique, De Boeck, 1997 (2e ed.)
Notes et références
- Classification phylogénétique du vivant, Lecointe et le Guyader, 2009 (3e ed) Belin (Paris)
- M. Ridley, Évolution Biologique, 2e ed. De Boeck, 1997.
- S. J. Gould, R. C. Lewontin, The spandrels of San Marco and the Panglossian paradigm: a critique of the adaptationist programme, Proc. R. Soc. Lond, 1979. 205:581-98
- E. M. Derrickson, R. E. Ricklefs, Taxon-dependent diversification of life-history traits and the perception of phylogenetic constraints, Funct. Ecol., 1988, 2:417-23
- Phylogenetic Constraint in Evolutionary Theory: Has It Any Explanatory Power?, Mary C. McKitrick, Annual Review of Ecology and Systematics, vol. 24 (1993), pp. 307-330
- S. J. Gould & E. S. Vrba, Exaptation-a missing term in the science of form, Paleobiology, 1982, p. 4-15.
- J. D. Ligon, The role of phylogenetic history in the evolution of contemporary avian mating and parental care systems, in Current ornithology, Springer US, 1993, p. 1-46.
- D. B. Burt, Evolutionary stasis, constraint and other terminology describing evolutionary patterns, Biological Journal of the Linnean Society, 72(4), 2001, p. 509-517.
- J. M. Smith, R. Burian, S. Kauffman, P. Alberch, J. Campbell, B. Goodwin, L. Wolpert, Developmental constraints and evolution: a perspective from the Mountain Lake conference on development and evolution, Quarterly Review of Biology, 1985, p. 265-287.
- H. A. Horner, H. C. Macgregor, Normal development in newts (Triturus) and its arrest as a consequence of an unusual chromosomal situation, Journal of herpetology, 1985, p. 261-270.
- M. R. Berenbaum, C. Favret, M. A. Schuler, On defining key innovations in an adaptive radiation: Cytochrome P450s and papilionidae, American Naturalist, 1996, p. 139-155.
- D. Parmley, J. A. Holman, Earliest fossil record of a pigmy rattlesnake (Viperidae: Sistrurus Garman), Journal of Herpetology, 41(1), 2007, p. 141-144.
- J. Felsenstein, Phylogenies and the comparative method, American Naturalist, 1985, p. 1-15.
Articles connexes
Lien externe
 Portail de la philosophie
Portail de la philosophie