Expansion planétaire de l'Homme moderne
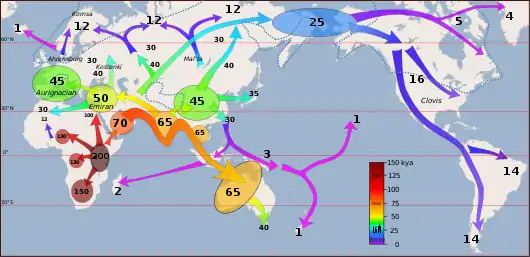
L'expansion planétaire de l'Homme moderne décrit la diffusion de l'Homme moderne à travers toute la planète, après sa sortie d'Afrique il y a environ 55 000 ans. Il n'a fallu que quelques milliers d'années à l'Homme moderne pour atteindre l'Australie vers l'Est et l'Europe vers le Nord, et un peu plus de temps pour atteindre l'Amérique puis les iles du Pacifique. Cette description s'appuie sur la théorie de l'origine africaine de l'Homme moderne.
Définition
Les vestiges fossiles découverts à ce jour donnent à Homo sapiens une origine africaine. Les données génétiques régulièrement publiées depuis 1987 indiquent que l'Homme moderne a émergé en Afrique. La distinction terminologique entre l'espèce Homo sapiens, identifiée par ses fossiles à partir de 300 000 ans, et l'Homme moderne, concept correspondant à l'humanité actuelle et défini à ce jour par la seule génétique, est régulièrement mise en avant par Jean-Jacques Hublin dans ses interventions, qui lui permettent notamment de souligner que l'Homme ne devient progressivement moderne, aussi bien anatomiquement que cognitivement, qu'il y a moins de 100 000 ans[1].
Des groupes d'Homo sapiens sont vraisemblablement sortis d'Afrique à plusieurs reprises entre 220 000 et 60 000 ans avant le présent (AP)[2]. On en trouve des vestiges fossiles en Israël datés de 185 000 ans[3], en Indonésie et en Chine datés respectivement de 68 000 et 110 000 ans, et peut-être même en Grèce âgés de 210 000 ans[4],[5]. Toutefois, selon les données génétiques, l'humanité non africaine actuelle serait entièrement issue de la dernière sortie d'Afrique, intervenue il y a seulement 55 000 ans[6], et qui aurait génétiquement effacé les sorties précédentes. Le présent article traite de l'expansion planétaire consécutive à cette ultime sortie d'Afrique, que l'on peut lire dans le génome des populations actuelles, et non des expansions antérieures, qui n'auraient laissé que des traces fossiles.
L'extension de l'Homme moderne à toute la planète se serait faite par micro-déplacements[7], de l'ordre d'une vingtaine de kilomètres par génération, lesquels, en l'espace de quelques centaines de générations, suffiraient à expliquer le peuplement de l'ensemble de l'Eurasie à partir de l'Afrique, sans qu'il y ait de migration au sens strict, à savoir un mouvement conscient et volontaire d'un ensemble de personnes d'un point à un autre dans un but donné[8].
À sa sortie d'Afrique, puis au cours de son expansion en Europe et en Asie, l'Homme moderne a rencontré d'autres espèces humaines, qui s'étaient développées localement pendant plusieurs centaines de milliers d'années : l'Homme de Néandertal, l'Homme de Denisova, l'Homme de Florès, l'Homme de Callao, et peut-être les derniers représentants d'Homo erectus en Asie du Sud-Est. Plusieurs épisodes d'hybridation se sont produits, dont les traces se lisent dans le génome de l'humanité actuelle. Cependant, l'héritage génétique de ces espèces dans notre génome demeure marginal, et les confrontations entre groupes ont vraisemblablement été nombreuses. Les espèces humaines locales ont finalement toutes disparu, probablement sous la pression exercée par l'Homme moderne[9],[1].
Génétique des populations
Deux parties du génome humain d'accès plus simple ont longtemps été seules exploitées pour déchiffrer l'histoire de l'Homme moderne. L'une est l'ADN mitochondrial et l'autre le chromosome Y. Ce sont les deux seules parties du génome qui ne se recombinent pas au moment de la fécondation. L'ADN mitochondrial et le chromosome Y sont transmis respectivement le long des lignées féminine et masculine.
ADN mitochondrial

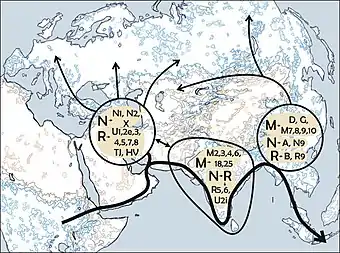
Les populations eurasiatiques appartiennent pour la plupart aux macro-haplogroupes M et N, qui descendent de l'haplogroupe est-africain L3.
De l'Arabie à l'Inde, la fréquence de l'haplogroupe M augmente vers l'est : dans l'Inde de l'est, l'haplogroupe M surpasse en nombre l'haplogroupe N par un ratio de 3 à 1. Cependant, plus on va vers l'Asie de l'Est, plus l'haplogroupe N réapparait en tant que lignage dominant. L'haplogroupe M est prédominant en Asie du Sud-Est mais parmi les Aborigènes d'Australie, l'haplogroupe N réapparait comme le lignage le plus commun[10].
ADN-Y
L'haplogroupe F est apparu il y a environ 45 000 ans, en Afrique du Nord (dans ce cas il montrerait une seconde vague de migration hors d'Afrique) ou en Asie du Sud. Plus de 90 % des hommes nés ailleurs qu'en Afrique descendent en ligne masculine du premier porteur de l'haplogroupe F.
ADN autosomal
L'ADN autosomal est la partie non sexuée du génome, qui est transmise par les deux parents et se recombine à chaque génération. Elle représente l'essentiel du génome (22 paires de chromosomes sur 23). En juin 2009, une analyse des données sur les génomes SNP de l'International HapMap Project (Phase II) et du CEPH Human Genome Diversity Panel samples a été publiée[11]. Ces modèles furent prélevés sur 1 138 individus qui n'étaient pas liés[11]. Avant cette analyse, les généticiens de population s'attendaient à trouver des différences importantes entre les groupes ethniques, avec des allèles partagés entre ces groupes mais différents ou n'existant pas dans les autres groupes[12]. Au lieu de cela, l'étude de 53 populations à partir des données des HapMap et CEPH a révélé que les groupes de population étudiés comprenaient seulement trois groupes génétiques : les Africains, les Eurasiens (qui incluent les natifs d'Europe et du Moyen-Orient, jusqu'à l'ouest de l'actuel Pakistan), et les Asiatiques de l'Est, qui incluent les natifs d'Asie orientale, d'Asie du Sud, des Amériques et d'Océanie[12]. L'étude a montré que la plupart des différences des groupes ethniques pouvaient être attribuées à la dérive génétique, et que les populations africaines modernes ont une plus grande diversité génétique que les deux autres groupes génétiques[12],[13]. Ce dernier point s'explique aisément quand on considère que la diversité génétique des Eurasiens n'a démarré qu'après la sortie d'Afrique, alors que celle des Africains a pu s'accumuler sur une période plus longue.
Sortie d'Afrique
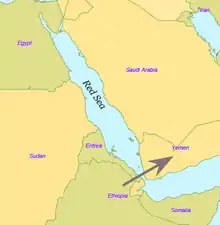
L'Homme moderne sort d'Afrique il y a environ 55 000 ans[6],[14]. L'ADN mitochondrial semble indiquer qu'un groupe de porteurs de l'haplogroupe L3 aurait migré d'Afrique orientale vers la péninsule arabique[15].
Certains chercheurs pensent que seules quelques personnes ont quitté l'Afrique dans le cadre d'une unique migration et qu'elles ont peuplé le reste du monde[réf. nécessaire]. Seul un petit groupe de près de 150 personnes aurait franchi la mer Rouge. C'est pourquoi, de tous les lignages présents en Afrique, seules les filles d'un seul lignage, L3, sont présentes hors d'Afrique. S'il y avait eu plusieurs migrations, on trouverait plus d'un lignage africain hors d'Afrique. Les filles du L3, les lignages M et N, sont peu fréquentes en Afrique subsaharienne (l'haplogroupe M1 est très ancien et diversifié en Afrique du Nord et en Afrique du Nord-Est) et semblent y être arrivées récemment. Une explication possible est que ces mutations se sont produites en Afrique de l'Est peu avant l'exode et, par effet fondateur, sont devenues les haplogroupes dominants après la sortie d'Afrique. Les mutations ont aussi pu se produire peu après la sortie d'Afrique.
D'autres chercheurs ont proposé un modèle de dispersion double selon lequel il y aurait eu deux sorties d'Afrique, dont l'une par la mer Rouge, qui se serait dirigée vers l'Inde en traversant les régions côtières (la route de la Côte), et qui serait représentée par l'Haplogroupe M. La seconde impliquerait un autre groupe, porteur de l'haplogroupe N, qui aurait suivi le Nil à partir de l'Afrique de l'Est, se dirigeant vers le nord et gagnant le Levant à travers le Sinaï. Puis, ce groupe se serait séparé dans plusieurs directions, certains allant en Europe et d'autres se dirigeant vers l'est, en Asie[16]. Cette hypothèse tente d'expliquer pourquoi l'haplogroupe N est prédominant en Europe et pourquoi l'haplogroupe M y est absent. Cependant, une population de pionniers européens qui, initialement, possédait à la fois les haplogroupes M et N pourrait avoir perdu l'haplogroupe M en raison d'une dérive génétique accélérée par un éventuel goulet d'étranglement de population.
Les preuves d'une migration vers l'est par la côte d'Arabie ont pu être en partie détruites par la montée du niveau de la mer pendant l'Holocène[17],[18].
Une calotte crânienne fossile d'Homo sapiens, notée Manot 1, découverte en 2008 dans la grotte de Manot, en Galilée occidentale (Israël), publiée en 2015 avec une datation de 54 700 ans +/- 5 500 ans avant le présent, est le plus ancien fossile de morphologie moderne trouvé à ce jour au Levant. Sa datation semble indiquer un probable contact des hommes modernes avec les populations néandertaliennes contemporaines du Levant, représentées par les fossiles néandertaliens de la grotte de Kébara, datés d'environ 60 000 ans, et de la grotte d'Amud, datés d'environ 55 000 ans. Selon les études génétiques, cette période correspond à l'époque estimée de l'hybridation des Néandertaliens avec les hommes modernes, juste après leur sortie d'Afrique. Cette découverte tend à appuyer la théorie d'une dernière sortie d'Afrique par le Levant plutôt que par la mer Rouge[19].
Hybridation avec Néandertal
Les études génétiques publiées depuis 2010 montrent une hybridation des hommes modernes non africains avec l'Homme de Néandertal, qui aurait eu lieu au Proche-Orient juste après leur sortie d’Afrique.
Le séquençage de l'ADN nucléaire néandertalien est effectué depuis 2010 par les équipes de l'Institut Max-Planck de Leipzig dirigées par Svante Pääbo[20]. La comparaison avec l'ADN de l'humanité actuelle montre que 1,8 % en moyenne du génome d'un non-africain actuel sont d'origine néandertalienne. Les populations non-africaines ayant sans aucune exception une part de gènes néandertaliens, et ce dans des proportions très proches, on en conclut que l'hybridation avec Néandertal s'est produite juste après la dernière sortie d'Afrique.
En 2014, l'étude du génome d'un fémur d'Homo sapiens, découvert à Ust-Ishim, en Sibérie occidentale, et daté d'environ 45 000 ans avant le présent, confirme l'hybridation avec Néandertal et permet d'estimer qu'elle aurait eu lieu environ 10 000 ans plus tôt, c'est-à-dire il y a environ 55 000 ans[6].
Diffusion australe
Il y a environ 55 000 ans, l'Homme moderne se répand du Moyen-Orient vers l'Asie du Sud en suivant plus ou moins la route côtière. Un peu plus tard, d'autres poursuivent vers l'Australie, qu'Homo sapiens a trouvée vierge de toute population humaine.
Le groupe qui a traversé la Mer Rouge aurait voyagé le long de la côte de l'Arabie et de la Perse jusqu'à atteindre l'Inde. Celle-ci semble être le premier lieu majeur d'expansion démographique. L'haplogroupe M est trouvé fréquemment le long des régions des côtes sud du Pakistan et de l'Inde et il possède sa plus grande diversité en Inde, montrant que c'est ici que les mutations se sont passées[21],[22]. 60 % de la population indienne appartient à l'haplogroupe M. Le peuple indigène des Îles Andaman appartient également au lignage M. On pense que les Andamanais possèdent les gènes des premiers habitants de l'Asie en raison de leur long isolement de l'Asie continentale. On trouve les traces de la route côtière des premiers colons d'Asie, de l'Inde jusqu'en Papouasie-Nouvelle-Guinée.
Puisque l'haplogroupe M est également trouvé fréquemment dans les hautes terres de la Nouvelle-Guinée, et que les Andamanais et les habitants de la Nouvelle-Guinée ont une peau sombre et des cheveux de type africain, certains scientifiques pensent que ceux-ci font tous partie de la même vague migratoire ayant franchi la Mer Rouge il y a environ 55 000 ans. Les recherches de Harding et al. (2000, p. 1355) montrent que, au moins en raison de la couleur de la peau sombre, le fond de l'haplotype MC1R des Papous-Néo Guinéens (un des nombreux gènes impliqués dans la production de mélanine) est le même que celui des Africains (à l'exception d'une seule mutation silencieuse). Ainsi, bien que ces groupes asiatiques soient génétiquement distincts des Africains, la sélection de la couleur de peau sombre a sûrement continué (au moins pour le MC1R) après l'exode. Les migrants originels venus d'Afrique ressemblaient probablement aux Africains actuels (au moins s'agissant de la couleur de la peau), ce qui expliquerait que les restes de ce phénotype soient trouvés chez les Andamanais et les Néo-Guinéens. D'autres suggèrent que leur ressemblance physique pourrait résulter d'une convergence évolutive[23],[24].
Hybridation avec Denisova
Plusieurs études publiées depuis 2010, basées sur le séquençage de l'ADN nucléaire extrait d'une phalange et de dents dénisoviennes trouvées dans la grotte de Denisova, en Sibérie, montrent que l'Homme de Denisova a contribué à hauteur de 2 à 4 % au génome des Mélanésiens et Aborigènes d'Australie actuels. Le génome des Asiatiques d'Extrême-Orient montre également des gènes dénisoviens, mais dans une proportion beaucoup plus faible d'environ 0,2 %. L'Homme de Denisova aurait été relativement répandu en Asie à la fin du Pléistocène[25].
Peuplement de l'Australie
Les premiers Homo sapiens sont peut-être arrivés en Australie il y a environ 65 000 ans. Durant la dernière glaciation, le niveau de la mer était beaucoup plus bas qu'aujourd'hui. La côte australienne se situait à environ 100 km de Timor. L'Australie et la Nouvelle-Guinée formaient un seul continent, appelé Sahul, et étaient reliées par un isthme situé au niveau de la mer d'Arafura, du golfe de Carpentarie et du détroit de Torrès. Des hommes auraient navigué sur de moyennes distances à partir des iles de l'actuelle Indonésie pour atteindre le Sahul. Puis, par voie terrestre, ils se seraient dispersés sur l'ensemble du continent.
Les fouilles archéologiques montrent des traces d'occupation humaine en amont de la Swan River, en Australie-Occidentale, il y a environ 40 000 ans. Il semble que la Tasmanie, elle aussi accessible par voie terrestre à l'époque, ait été atteinte vers 30 000 ans avant le présent.

La route du Nord
La différenciation entre groupes humains est due à leur séparation géographique, favorisant une dérive génétique propre à chaque groupe. Les populations d'Homo sapiens ayant atteint le centre de l'Eurasie il y a 50 000 ans se sont retrouvées isolées par les masses glaciaires de l'Himalaya, de l'Hindou Kouch, ainsi que par les déserts arides, lors du stade isotopique 3 de la glaciation de Würm (57 000 à 30 000 ans AP). Avec une intense pression climatique, ils ont dû s'adapter, physiquement et culturellement, à des climats froids, tout comme l'avaient fait avant eux les hommes de Denisova en Asie. Ces migrations vers des environnements moins ensoleillés expliquent la dépigmentation des populations européennes et nord-asiatiques, tandis que les populations restant dans les climats tropicaux d'Asie du Sud ont conservé leur pigmentation les protégeant du soleil, comme les Négritos des îles Andaman, les Papous ou les Aborigènes d'Australie.
Le Japon est atteint il y a 30 à 40 000 ans.
Peuplement de l'Europe
Homo sapiens arrive en Europe orientale il y a environ 48 000 ans[26]. Il lui faudra 20 000 ans pour remplacer complètement l'Homme de Néandertal, établi en Europe depuis 400 000 ans et particulièrement bien adapté au climat glaciaire. Certains des premiers hommes modernes arrivés en Europe se croisent avec les Néandertaliens d'Europe, mais leur descendance s'est probablement éteinte, car ces épisodes d'hybridation n'ont pas laissé de traces dans le génome des Européens actuels[27].
Les sites du début du Paléolithique supérieur en Europe centrale montrent différentes industries lithiques dites « de transition », plus ou moins intermédiaires entre le Moustérien des Néandertaliens et l'Aurignacien des hommes modernes, et que les chercheurs hésitent à attribuer à l'un ou l'autre des deux groupes. Il faut attendre 43 000 ans avant le présent (AP) pour que l'Aurignacien apparaisse en Allemagne, et 42 000 ans AP en France[26]. Vers 30 000 ans AP, Néandertal n'occupe plus en Europe de l'Ouest que la péninsule Ibérique.
Le territoire des Néandertaliens se serait lentement réduit, la compétition entre les deux groupes n'étant pas toujours à l'avantage de sapiens. Les Néandertaliens étaient en effet plus lourds et plus robustes, et donc physiquement plus forts que les hommes modernes, tout en ayant aussi des capacités cognitives avancées. Probablement avantagés par de meilleures facultés de coopération, les hommes modernes ont remplacé progressivement les Néandertaliens[9], dont les derniers refuges en Europe furent le sud de la péninsule Ibérique et la Crimée.
Peuplement de l'Amérique
L'époque de l'arrivée des premiers hommes en Amérique reste controversée, entre 40 000 et 20 000 ans avant le présent. Les actuels Amérindiens descendent de populations du Nord-est sibérien qui ont gagné l'Alaska en traversant la Béringie, et se sont ensuite diffusées jusqu'à l'extrême Sud de l'Amérique du Sud.
Peuplement du Pacifique
Vers 1 500 av. J.-C., des groupes austronésiens se déplacent vers les iles orientales de l'Indonésie, puis, devenus les Polynésiens, ils gagnent l'une après l'autre les iles du Pacifique, atteignant finalement la Nouvelle-Zélande il y a seulement 800 ans.
Références
- Jean-Jacques Hublin, « Aux origines d'Homo sapiens », sur Colloque international INRAP : Être humain ? Archéologie des origines, Muséum de Toulouse, 9-10 novembre 2018
- Jean-Jacques Hublin et Bernard Seytre, Quand d'autres hommes peuplaient la terre : nouveaux regards sur nos origines, Flammarion, (ISBN 9782081260436)
- (en) Israel Hershkovitz, Gerhard W. Weber, Rolf Quam, Mathieu Duval, Rainer Grün et al., « The earliest modern humans outside Africa », Science, vol. 359, no 6374, , p. 456-459 (DOI 10.1126/science.aap8369)
- (en) Eric Delson, « An early dispersal of modern humans from Africa to Greece », Nature, (DOI 10.1038/d41586-019-02075-9)
- (en) Katerina Harvati, Carolin Röding, Abel M. Bosman, Fotios A. Karakostis, Rainer Grün et al., « Apidima Cave fossils provide earliest evidence of Homo sapiens in Eurasia », Nature, (DOI 10.1038/s41586-019-1376-z)
- (en) Qiaomei Fu, Heng Li, Svante Pääbo et al., « The genome sequence of a 45,000-year-old modern human from western Siberia », Nature, vol. 514, no 7523, , p. 445–449 (DOI 10.1038/nature13810, lire en ligne)
- Laure Cailloce, « De tout temps, l’homme s’est déplacé », CNRS Le journal, (lire en ligne)
- Dominique Garcia et Hervé Le Bras, Archéologie des migrations, La Découverte, , 363 p. (présentation en ligne), p. 6
- (en) Nick Longrich, « Were other humans the first victims of the sixth mass extinction ? », The Conversation, (lire en ligne)
- (en) Max Ingman et Ulf Gyllensten, « Mitochondrial Genome Variation and Evolutionary History of Australian and New Guinean Aborigines », Genome Research, vol. 13, no 7, (lire en ligne)
- (en) Coop G, Pickrell JK, Novembre J, Kudaravalli S, Li J, et al, « The Role of Geography in Human Adaptation », PLoS Genet, vol. 5, no 6, , p. 1–16 (DOI 10.1371/journal.pgen.1000500, lire en ligne)
- « Among Many Peoples, Little Genomic Variety », The Washington Post, (consulté le )
- « Geography And History Shape Genetic Differences In Humans », Science Daily, (consulté le )
- (en) Christopher J. Bae, Katerina Douka et Michael D. Petraglia, « On the origin of modern humans : Asian perspectives », Science, vol. 358, no 6368, (DOI 10.1126/science.aai9067)
- (en) P Soares, F Alshamali, J. B Pereira, V Fernandes, N. M Silva, C Afonso, M. D Costa, E Musilova, V MacAulay, M. B Richards, V Cerny et L Pereira, « The Expansion of mtDNA Haplogroup L3 within and out of Africa », Molecular Biology and Evolution, vol. 29, no 3, , p. 915–927 (PMID 22096215, DOI 10.1093/molbev/msr245)
- (en) P. Marrero, K.K. Abu-Amero, J.M. Larruga et al., « Carriers of human mitochondrial DNA macrohaplogroup M colonized India from southeastern Asia », BMC Evolutionary Biology, vol. 16, no 246, (DOI 10.1186/s12862-016-0816-8)
- (en) Nicole Maca-Meyer, Ana M. González, José M. Larruga, Carlos Flores et Vicente M. Cabrera, « Major genomic mitochondrial lineages delineate early human expansions », BMC Genetics, vol. 2, no 13, (DOI 10.1186/1471-2156-2-13)
- (en) Marta Mirazón Lahr, « Searching for traces of the Southern Dispersal » (version du 4 août 2004 sur l'Internet Archive), University of Cambridge, sur www.human-evol.cam.ac.uk
- (en) Israel Hershkovitz, Ofer Marder et al., « Levantine cranium from Manot Cave (Israel) foreshadows the first European modern humans », Nature, vol. 520, no 7546, , p. 216–219 (PMID 25629628, DOI 10.1038/nature14134)
- (en) Richard E. Green et al., « A Draft Sequence of the Neandertal Genome », Science, vol. 328, no 5979, , p. 710-722 (DOI 10.1126/science.1188021, lire en ligne)
- (en) Vincent Macaulay, « Single, Rapid Coastal Settlement of Asia Revealed by Analysis of Complete Mitochondrial Genomes », Science Magazine, vol. 308, no 5724, , p. 1034–1036 (lire en ligne)
- Most of the extant mtDNA boundaries in South and Southwest Asia were likely shaped during the initial settlement of Eurasia by anatomically modern humans
- Genetic origins of the Andaman Islanders
- « Genetic affinities of the Andaman Islanders »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?) (consulté le )
- (en) Reich et al., « Genetic history of an archaic hominin group from Denisova Cave in Siberia », Nature, vol. 468, p. 1053-1060, 23 décembre 2010
- (en) Jean-Jacques Hublin, « The modern human colonization of western Eurasia : when and where ? », Quaternary Science Reviews, vol. 118, , p. 194-210 (DOI 10.1016/j.quascirev.2014.08.011, lire en ligne)
- François Savatier, « Un métissage tardif entre Sapiens et Néandertal en Europe », Pour la Science, (lire en ligne)
Bibliographie
- (en) Jean-Jacques Hublin, « The modern human colonization of western Eurasia : when and where ? », Quaternary Science Reviews, vol. 118, , p. 194-210 (DOI 10.1016/j.quascirev.2014.08.011, lire en ligne)
- (en) Stephen Oppenheimer, The Real Eve : Modern Man's Journey Out of Africa, (ISBN 0-7867-1192-2) (Hardcover), 2003.
- (en) Nicholas Wade, Before the Dawn : Recovering the Lost History of Our Ancestors, 2006 (ISBN 1-59420-079-3)
- (en) Wells, Spencer, The Journey of Man : A Genetic Odyssey, 2003 (ISBN 0-691-11532-X)
- (en) Wells, Spencer, Deep Ancestry : Inside the Genographic Project, 2006 (ISBN 0-7922-6215-8)
- (en) DNA studies trace human migration from Africa, 21 février 2008
Classiques
- Luigi Luca Cavalli-Sforza & Francesco Cavalli-Sforza, Qui sommes-nous ? Une histoire de la diversité humaine, éd. Albin Michel, 1994 (titre original : Chi Siamo : La Storia della Diversità Umana, 1993), (ISBN 0-201-44231-0).
- Bryan Sykes, Les Sept Filles d'Ève : Génétique et histoire de nos origines, trad. Pierre-Emmanuel Dauzat, Éditions Albin Michel, 2001, 363 p. (ISBN 2-226-12617-1) ; rééd. Librairie générale française, collection « Le Livre de poche » (no 15588), 2003, 344 p. (ISBN 2-253-15588-8)
Voir aussi
Articles connexes
Liens externes
 Portail de la Préhistoire
Portail de la Préhistoire  Portail de l’anthropologie
Portail de l’anthropologie