Histone H1
L'histone H1 est l'une des cinq principales protéines histone qui forment la chromatine chez les cellules eucaryotes. Bien que hautement conservée , il s'agit néanmoins de l'histone présentant la plus haute variabilité de séquence inter-espèces.

Structure
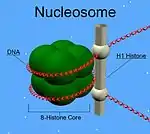
Chez les métazoaires, les protéines H1 présentent un domaine d'"hélice ailée" globulaire central et de longues queues C et N-terminales. H1 est impliquée dans le regroupement des sous-structures dites en "collier de perles" en une structure de niveau supérieur, dont les détails n'ont pas encore été résolus[1]. Les histones H1 présentes chez les protistes et les bactéries sont dépourvues du domaine central et de la queue N-terminale[2].
Fonction
Contrairement aux autres histones, H1 ne constitue pas la particule nucléosomale. Elle repose sur la structure nucléosomale, en maintenant en place l'ADN enroulé autour du nucléosome. Contrairement aux autres histones représentées par deux molécules par type d'histone par nucléosome, il n'y a qu'une molécule H1 par nucléosome. Au-delà de sa liaison au nucléosome, la protéine H1 se lie à l'ADN de liaison (ou encore ADN linker, d'environ 20 à 80 nucléotides de long) entre les nucléosomes, aidant ainsi à stabiliser la fibre chromatinienne de 30 nm[3]. La plupart des connaissances sur H1 découle d'études sur des fibres de chromatine purifiées. L'extraction ionique d'histones de liaison à partir de chromatine native ou reconstituée favorise son dépliement de fibres de 30 nm de longueur vers une conformation nucléosomale en collier de perles dans des conditions hypotoniques[4],[5],[6].
Il n’est pas certain qu'H1 favorise la formation d'une fibre chromatinienne en forme de solénoïde, dans laquelle l’ADN de liaison exposé est raccourci, ou si elle favorise simplement un changement de l’angle des nucléosomes adjacents, sans affecter la longueur de l'ADN linker[7]. Cependant, il fut montré que les histones linker favorisent la compaction des fibres de chromatine reconstituées in vitro à l’aide d’ensembles d'ADNs synthétiques reconstituant l'élément de positionnement nucléosomal fort «601»[8]. Les expériences de digestion par nucléase et de détections d'empreintes génomique suggèrent que le domaine globulaire de l'histone H1 est localisé près de la dyade du nucléosome, où il protège environ 15 à 30 paires de bases d'ADN additionnel[9],[10],[11],[12]. De plus, des expériences sur la chromatine reconstituée ont révélé un motif caractéristique dans la dyade en présence de H1[13]. Malgré des lacunes dans la compréhension du rôle d'H1, un modèle général est apparu selon lequel le domaine globulaire de H1 ferme le nucléosome en liant l'ADN entrant et sortant, tandis que la queue se lie à l'ADN de liaison et neutralise sa charge négative[7],[11].
De nombreuses expériences portant sur la fonction d'H1 ont été réalisées sur de la chromatine traitée et purifiée dans des conditions de faible teneur en sel, mais le rôle d'H1 in vivo est moins certain. Des études cellulaires ont montré que la surexpression d'H1 peut provoquer une morphologie nucléaire et une structure chromatinienne aberrantes, et qu'H1 peut servir à la fois de régulateur positif et négatif de la transcription, en fonction des gènes[14],[15],[16]. Dans les extraits d'œufs de Xenopus , l'appauvrissement en histones linker provoque une extension longitudinale d'environ 2 fois des chromosomes mitotiques, tandis qu'une surexpression provoque une hypercompaction des chromosomes en une masse inséparable[17],[18]. L'inactivation complète d'H1 in vivo n'a pas été atteinte chez les organismes multicellulaires en raison de la présence de multiples isoformes pouvant être présentes dans plusieurs clusters de gènes, mais diverses isoformes de l'histone de liaison ont été déplétés à divers degrés chez Tetrahymena, C. elegans, Arabidopsis, la mouche et la souris, entraînant divers défauts organisme-spécifiques dans la morphologie nucléaire, la structure de la chromatine, la méthylation de l’ADN et / ou l’expression de gènes spécifiques[19],[20],[21].
Dynamique
Alors que la plupart des histones H1 du noyau sont liées à la chromatine, les molécules H1 effectuent une navette entre les régions de la chromatine à une vitesse assez élevée[22],[23].
Il est difficile de comprendre en quoi protéine si dynamique pourrait être un composant structurel de la chromatine, mais il a été suggéré qu'à l'équilibre nucléaire, l'association entre H1 et la chromatine est encore fortement favorisée, ce qui signifie que malgré sa dynamique, la grande majorité d'H1 à tout moment donné est liée à la chromatine[24]. H1 compacte et stabilise l'ADN soumis à des forces et pendant l'assemblage de la chromatine, ce qui suggère que la liaison dynamique d'H1 pourrait conférer une protection à l'ADN dans les cas où des nucléosomes doivent être éliminés[25].
Des facteurs cytoplasmiques semblent être nécessaires à l'échange dynamique de l'histone H1 sur la chromatine, mais ils n'ont pas encore été spécifiquement identifiés[26]. La dynamique d'H1 peut être influencée dans une certaine mesure par la O-glycosylation et la phosphorylation. La O-glycosylation d'H1 peut favoriser la condensation et le compactage de la chromatine. Il a été démontré que la phosphorylation au cours de l’interphase diminue l’affinité d'H1 pour la chromatine et peut favoriser la décondensation de la chromatine et la transcription active. Cependant, pendant la mitose, il a été démontré que la phosphorylation augmentait l'affinité d'H1 envers les chromosomes et favorisait donc la condensation des chromosomes mitotiques[18].
Isoformes
Chez les animaux, la famille H1 comprend de multiples isoformes H1 qui peuvent être exprimés au sein d’un même organisme dans des tissus et stades de développement différents ou se chevauchant . La raison de l'existence ces multiples isoformes reste incertaine, mais leur conservation évolutive de l'oursin à l'homme ainsi que des différences significatives dans leurs séquences d'acides aminés suggèrent qu'elles ne sont pas fonctionnellement équivalentes[27],[28],[29]. L’une des isoformes est l’histone H5 , que l’on ne trouve que dans les érythrocytes aviaires, qui ne ressemblent pas aux érythrocytes de mammifères en ce sens qu’ils présentent un noyau. Un autre isoforme est l'isoforme H1M zygotique / zygotique (également appelée B4 ou H1foo), trouvée chez les oursins, les grenouilles, les souris et les humains, qui est remplacée dans l'embryon par les isoformes somatiques H1A-E et H10 qui ressemble à H5[29],[30],[31],[32]. Bien qu’elle ait plus de charges négatives que les isoformes somatiques, H1M se lie avec une affinité plus élevée pour les chromosomes mitotiques dans les extraits d’œufs de Xenopus[18].
Modifications post-traductionnelles
Comme les autres histones, les histones de la famille H1 sont largement modifiées après traduction. Cela comprend la phosphorylation de résidus sérine et thréonine, l'acétylation et méthylation de résidus lysine, et l'ubiquitination[33]. Ces modifications post-traductionnelles remplissent diverses fonctions, mais sont moins bien étudiés que celles d'autres histones.
Voir également
- Nucléosome
- Histone
- La chromatine
- Linker histone H1 variants
- Autres protéines histones impliquées dans la chromatine :
Références
- « Crystal structure of globular domain of histone H5 and its implications for nucleosome binding », Nature, vol. 362, no 6417, , p. 219–23 (PMID 8384699, DOI 10.1038/362219a0)
- KASINSKY, LEWIS, DACKS et AUSIÓ, « Origin of H1 linker histones », The FASEB Journal, vol. 15, no 1, , p. 34–42 (PMID 11149891, DOI 10.1096/fj.00-0237rev)
- (en) Jeon, Kwang W. et Berezney, Ronald, Structural and functional organization of the nuclear matrix, Boston, Academic Press, , 214–7 p. (ISBN 978-0-12-364565-4)
- « Solenoidal model for superstructure in chromatin », Proc. Natl. Acad. Sci. U.S.A., vol. 73, no 6, , p. 1897–901 (PMID 1064861, PMCID 430414, DOI 10.1073/pnas.73.6.1897)
- « Influence of histone H1 on chromatin structure », Cell, vol. 12, no 1, , p. 101–7 (PMID 561660, DOI 10.1016/0092-8674(77)90188-X)
- « Involvement of histone H1 in the organization of the nucleosome and of the salt-dependent superstructures of chromatin », J. Cell Biol., vol. 83, no 2 Pt 1, , p. 403–27 (PMID 387806, PMCID 2111545, DOI 10.1083/jcb.83.2.403)
- « What determines the folding of the chromatin fiber? », Proc. Natl. Acad. Sci. U.S.A., vol. 93, no 20, , p. 10548–55 (PMID 8855215, PMCID 38190, DOI 10.1073/pnas.93.20.10548)
- Routh, Sandin et Rhodes, « Nucleosome repeat length and linker histone stoichiometry determine chromatin fiber structure. », Proceedings of the National Academy of Sciences of the United States of America, vol. 105, no 26, , p. 8872–7 (PMID 18583476, PMCID 2440727, DOI 10.1073/pnas.0802336105)
- « Heterogeneity of chromatin subunits in vitro and location of histone H1 », Nucleic Acids Res., vol. 3, no 2, , p. 477–92 (PMID 1257057, PMCID 342917, DOI 10.1093/nar/3.2.477)
- « Removal of histone H1 exposes a fifty base pair DNA segment between nucleosomes », Biochemistry, vol. 15, no 15, , p. 3307–14 (PMID 952859, DOI 10.1021/bi00660a022)
- « The structure of histone H1 and its location in chromatin », Nature, vol. 288, no 5792, , p. 675–9 (PMID 7453800, DOI 10.1038/288675a0)
- « Footprinting of linker histones H5 and H1 on the nucleosome », EMBO J., vol. 7, no 12, , p. 3685–91 (PMID 3208745, PMCID 454941, DOI 10.1002/j.1460-2075.1988.tb03250.x)
- « Nucleosomes, linker DNA, and linker histone form a unique structural motif that directs the higher-order folding and compaction of chromatin », Proc. Natl. Acad. Sci. U.S.A., vol. 95, no 24, , p. 14173–8 (PMID 9826673, PMCID 24346, DOI 10.1073/pnas.95.24.14173)
- « The maternal histone H1 variant, H1M (B4 protein), is the predominant H1 histone in Xenopus pregastrula embryos », Dev. Biol., vol. 161, no 2, , p. 425–39 (PMID 8313993, DOI 10.1006/dbio.1994.1042)
- « Differential effect of H1 variant overexpression on cell cycle progression and gene expression », Nucleic Acids Res., vol. 24, no 3, , p. 486–93 (PMID 8602362, PMCID 145659, DOI 10.1093/nar/24.3.486)
- « Effects of H1 histone variant overexpression on chromatin structure », J. Biol. Chem., vol. 274, no 53, , p. 37950–6 (PMID 10608862, DOI 10.1074/jbc.274.53.37950)
- « Histone H1 is essential for mitotic chromosome architecture and segregation in Xenopus laevis egg extracts », J. Cell Biol., vol. 169, no 6, , p. 859–69 (PMID 15967810, PMCID 2171634, DOI 10.1083/jcb.200503031)
- « Functional Comparison of Linker Histones in Xenopus Reveals Isoform-Specific Regulation by Cdk1 and RanGTP », Curr. Biol., vol. 20, no 11, , p. 1048–52 (PMID 20471264, PMCID 2902237, DOI 10.1016/j.cub.2010.04.025)
- « Linker histones are not essential and affect chromatin condensation in vivo », Cell, vol. 82, no 1, , p. 47–56 (PMID 7606784, DOI 10.1016/0092-8674(95)90051-9)
- « A single histone H1 isoform (H1.1) is essential for chromatin silencing and germline development in Caenorhabditis elegans », Development, vol. 128, no 7, , p. 1069–80 (PMID 11245572)
- « Linker histone H1 is essential for Drosophila development, the establishment of pericentric heterochromatin, and a normal polytene chromosome structure », Genes Dev., vol. 23, no 4, , p. 452–65 (PMID 19196654, PMCID 2648648, DOI 10.1101/gad.1749309)
- « Dynamic binding of histone H1 to chromatin in living cells », Nature, vol. 408, no 6814, , p. 877–81 (PMID 11130729, DOI 10.1038/35048610)
- « Condensed mitotic chromatin is accessible to transcription factors and chromatin structural proteins », J. Cell Biol., vol. 168, no 1, , p. 41–54 (PMID 15623580, PMCID 2171683, DOI 10.1083/jcb.200407182)
- « The dynamics of histone H1 function in chromatin », Mol. Cell, vol. 17, no 5, , p. 617–20 (PMID 15749012, DOI 10.1016/j.molcel.2005.02.019)
- Xiao, Freedman, Miller et Heald, « Histone H1 compacts DNA under force and during chromatin assembly », Molecular Biology of the Cell, vol. 23, no 24, , p. 4864–4871 (PMID 23097493, PMCID 3521692, DOI 10.1091/mbc.E12-07-0518)
- « Xenopus Egg Extracts Increase Dynamics of Histone H1 on Sperm Chromatin », PLoS ONE, vol. 5, no 9, , e13111 (PMID 20927327, PMCID 2947519, DOI 10.1371/journal.pone.0013111)
- « Somatic linker histones cause loss of mesodermal competence in Xenopus », Nature, vol. 389, no 6649, , p. 395–9 (PMID 9311783, DOI 10.1038/38755)
- « Histone H1 variants differentially inhibit DNA replication through an affinity for chromatin mediated by their carboxyl-terminal domains », Gene, vol. 292, nos 1–2, , p. 173–81 (PMID 12119111, DOI 10.1016/S0378-1119(02)00675-3)
- « The histone H1 family: specific members, specific functions? », Biol. Chem., vol. 389, no 4, , p. 333–43 (PMID 18208346, DOI 10.1515/BC.2008.037)
- Khochbin S, « Histone H1 diversity: bridging regulatory signals to linker histone function », Gene, vol. 271, no 1, , p. 1–12 (PMID 11410360, DOI 10.1016/S0378-1119(01)00495-4)
- « Cracking the enigmatic linker histone code », J. Biochem., vol. 143, no 3, , p. 287–93 (PMID 18234717, DOI 10.1093/jb/mvn013)
- « Histone H1 and its isoforms: contribution to chromatin structure and function », Gene, vol. 431, nos 1–2, , p. 1–12 (PMID 19059319, DOI 10.1016/j.gene.2008.11.003)
- « H1 histones: current perspectives and challenges », Nucleic Acids Res., vol. 21, no 41, , p. 9593–609 (PMID 23945933, PMCID 3834806, DOI 10.1093/nar/gkt700)
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire