Interleukine 13
L' interleukine 13 ( IL-13 ) est une protéine codée par le gène IL13 chez l'homme. C'est une cytokine sécrétée principalement par des cellules T activées. Elle est impliquée dans plusieurs étapes de la maturation et de la différentiation des cellules B. Capable d'augmenter l'expression de CD23 et du CMH de type II et de promouvoir le changement d'isotype des IgE sur les cellules B. L'Interleukine 13 est capable de réduire l'activité des macrophages et donc d'inhiber la production de certaines cytokines pro-inflammatoires. C'est un médiateur de l'inflammation allergique et de différentes maladies dont l'asthme .
| Interleukine 13 | ||
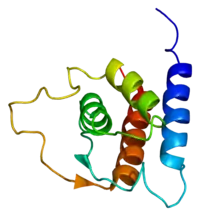 Structure de la protéine IL13. Basé sur la structure PDB 1ga3 analysée par PyMOL | ||
| Caractéristiques générales | ||
|---|---|---|
| Nom approuvé | Interleukine 13 | |
| Symbole | IL13 | |
| Homo sapiens | ||
| Locus | 5q31.1 | |
| Masse moléculaire | 15 816 Da[1] | |
| Nombre de résidus | 146 acides aminés[1] | |
| Entrez | 3596 | |
| HUGO | 5973 | |
| OMIM | 147683 | |
| UniProt | P35225 | |
| RefSeq (ARNm) | NM_002188.3 | |
| RefSeq (protéine) | NP_002179.2 | |
| Ensembl | ENSG00000169194 | |
| PDB | 1GA3, 1IJZ, 1IK0, 3BPO, 3G6D, 3L5W, 3L5X, 3LB6, 4I77, 4PS4, 5E4E | |
|
GENATLAS • GeneTests • GoPubmed • HCOP • H-InvDB • Treefam • Vega | ||
| Liens accessibles depuis GeneCards et HUGO. | ||
Génomique
L'interleukine 13 est une protéine de la famille des interleukines[2],[3],[4]. La séquence nucléique de l'ARNm de l'IL-13 d'une longueur de 1,4 kba été clonée pour la première fois en 1993[2]. Elle est codée par le gène IL13 et ce dernier forme un cluster de gènes de cytokines sur le chromosome 5q31 avec les gènes IL3, IL5, IL4, et CSF2. Le gène IL13 est particulièrement positionné à proximité de celui de IL4[2],[4].
La protéine
La forme non glycolsylée de l'Interleukine 13 a une masse de 12.4 kDa et se replie en 4 faisceaux alpha hélicoïdaux[2],[5]. Les caractéristiques structurelles secondaires de l'IL-13 sont similaires à celles de l' interleukine 4 (IL-4); cependant, il n'a que 25 % d'homologie de séquence protéique avec l'IL-4 mais les zones correspondantes aux hélices alpha 1 et 4 critiques pour l'activité de l'interleukine 4 sont les plus conservées entre les deux protéines[2]. L'homologie de la forme humaine d'IL13 avec celle de souris environ de 58%[3].
Production d'IL13
L'IL-13 est une cytokine sécrétée par les cellules T helper de type 2 (Th2), les cellules CD4, les cellules T tueuses naturelles, les mastocytes, les basophiles, les éosinophiles et les nuocytes[5] . L'interleukine-13 est un régulateur central de la synthèse des IgE, de l'hyperplasie des cellules caliciformes, de l'hypersécrétion de mucus, de l'hyperréactivité des voies respiratoires, de la fibrose et de la régulation positive de la chitinase[5]. C'est un médiateur de l' inflammation allergique et de différentes maladies dont l' asthme[5].
Les fonctions
L'IL-13 a des effets sur les cellules immunitaires similaires à ceux de la cytokine étroitement apparentée IL-4[2]. In vitro, elle a montré un niveau d'inhibition de la synthèse d'IL6 comparable à celle provoquée par IL4[2]. IL13 est capable d'une signalisation indépendante de l'IL-4 ne se fixant pas sur le récepteur de l'IL4[5],[6]. Cependant, l'IL-13 est suspectée d'être le médiateur central des changements physiologiques induits par l'inflammation allergique dans de nombreux tissus[5].
Bien que l'IL-13 soit principalement associée à l'induction de maladies des voies respiratoires, elle possède également des propriétés anti-inflammatoires[2]. L'IL-13 induit une classe d'enzymes dégradant les protéines, appelées métalloprotéinases matricielles (MMP) et notamment la MMP2 endogène dans les fibroblastes des voies respiratoires[7]. Ces enzymes sont nécessaires pour induire l'agression des cellules inflammatoires du parenchyme dans la lumière des voies respiratoires, où elles sont ensuite éliminées. Entre autres facteurs, l'IL-13 induit ces MMP dans le cadre d'un mécanisme de protection contre l'inflammation allergique excessive qui prédispose à l'asphyxie et la production de MMP2 induite par IL13 est beaucoup plus importante et significative dans les fibroblastes de patients asthmatiques que dans les populations témoins[7].
L'IL-13 est connue pour induire des changements dans les cellules hématopoïétiques, mais ces effets sont probablement moins importants que ceux de l'IL-4[2],[3]. De plus, l'IL-13 peut induire la sécrétion d'immunoglobulines E (IgE) à partir de cellules B humaines activées[3],[5]. L'interleukine-13 est un régulateur central de la synthèse des IgE mais est aussi capable est également impliquée dans l'induction de la production d'IgM et d'IgG en présence de cellules T CD4 positives[3]. Elle est capable d'induire le changement des isotypes des immunoglobulines[3]. La suppression de l'IL-13 chez la souris n'affecte pas de manière marquée le développement des cellules Th2 ou les réponses IgE spécifiques à l'antigène induites par des allergènes puissants[2]. En comparaison, la suppression de l'IL-4 désactive ces réponses. Ainsi, plutôt qu'une cytokine lymphoïde, l'IL-13 agit davantage comme un pont moléculaire reliant les cellules inflammatoires allergiques aux cellules non immunitaires en contact avec elles, altérant ainsi la fonction physiologique[2].
La signalisation de l'IL-13 s'effectue grâce à un complexe de récepteurs hétérodimères constitué du récepteur alpha IL-4 (IL-4Rα) et du récepteur alpha de l'interleukine-13 (IL13RA1)[5]. L'Interleukine 13 se fixe d'abord sur son récepteur IL13RA1. La forte affinité de l'IL-13 pour l'IL-13RA1 conduit à leur formation de liaisons qui est suivie d'une formation d'un hétérodimère avec l'IL4R. L'IL4R s'associe avec JAK1 tandis que l'IL13RA1 s'associe avec la protéine JAK2 ou TYK2 mais pas JAK3[5]. L'hétérodimérisation active JAK1 permettant la phosphorylation de tyrosine intracellulaire de l'IL4R . La protéine STAT6 est alors capable de s'arrimer au complexe[5]. STAT6 va être phosphorylée et va se dimériser, puis se transloquer dans le noyau pour réguler la transcription de gènes dépendant de l'IL13[5]. La signalisation STAT6 est importante dans l'initiation de la réponse allergique[5].
La plupart des effets biologiques de l'IL-13, comme ceux de l'IL-4, sont liés à un seul facteur de transcription, transducteur de signal et activateur de la transcription 6 ( STAT6 )[5]. L'interleukine-13 et ses récepteurs associés avec la sous-unité du récepteur IL-4 (IL-4Rα) permettent l'activation en aval de STAT6[8]. Les protéines JAK Janus kinase arrimées à l'extrémité cytoplasmique des récepteurs permettent la phosphorylation de STAT6, qui forme alors un homodimère activé et est transporté vers le noyau. [8] Une fois, dans le noyau, la molécule d'homodimère STAT6 régule l'expression génique des types cellulaires essentiels à l'équilibre entre la défense immunitaire de l'hôte et les réponses inflammatoires allergiques telles que le développement de Th2[8]. Cela peut être le résultat d'une réaction allergique provoquée face à un gène Ala.
L'IL-13 se lie également à un autre récepteur appelé IL-13Rα2[9]. L'IL-13Rα2 (qui est marqué comme un récepteur leurre) est dérivé des cellules Th2 et est une cytokine régulatrice immunitaire pléotrope[9]. L'IL-13 a une plus grande affinité (50 fois) pour l'IL-13Rα2 que pour l'IL-13Ra1[9]. La sous-unité IL-13Rα2 se lie uniquement à l'IL-13 et existe à la fois sous des formes liées à la membrane et solubles chez la souris[9]. Aucune forme soluble d'IL-13Rα2 n'a été détectée chez les sujets humains[9].
Ce récepteur IL-13Rα2 est aussi très fortement surexprimé dans des gliomes et impliqué dans le croissance, la migration et l'invasion de ces cellules cancéreuses[9]. Les mécanismes d'actions sont liés à l'activation de la protéine Scr pour l'invasion et probablement par l'intermédiaire de'Akt, de PI3K et mTOR pour la croissance et la migration métastatique[9].
Des études sur des poumons de souris transgéniques IL-13 avec des loci nuls d'IL-13Rα2 ont indiqué que la carence en IL-13Rα2 augmentait significativement l'inflammation et le remodelage pulmonaires induits par l'IL-13 ou l'ovalbumine[9]. La plupart des cellules normales, telles que les cellules immunitaires ou les cellules endothéliales, expriment des niveaux très faibles ou indétectables de récepteurs de l'IL-13[9]. La recherche a montré que l'expression à la surface cellulaire de l'IL-13Rα2 sur les fibroblastes des voies respiratoires asthmatiques humaines était réduite par rapport à l'expression sur les fibroblastes des voies respiratoires témoins normaux[9]. Cela a soutenu l'hypothèse selon laquelle l'IL-13Rα2 est un régulateur négatif de la réponse induite par l'IL-13 et a illustré une production significativement réduite de TGF-β1 et le dépôt de collagène dans les poumons des souris[9].
L'interleukine-13 joue un rôle essentiel dans la métaplasie des cellules caliciformes[10]. Les cellules caliciformes sont remplies de mucine (MUC)[10]. La mucine 5AC (MUC5AC) est le gène dominant de mucine produit dans les cellules caliciformes[10]. Dans quasiment tous les cas d'asthmes (faible, modéré ou sévère), les cellules caliciformes subissent des changements notamment dans le stockage et la sécrétion de mucine[10]. L'interleukine-13 induit la différenciation des cellules caliciformes et permet la production de MUC5AC dans l'épithélium trachéal[11]. La 15-Lipoxygénase-1 (ALOX15), une enzyme du métabolisme des acides gras et ses métabolites dont le 15-HETE, le 15-HETE-PE et le 13-HODE sont fortement exprimés dans l'asthme (qui conduisent à la surexpression de MUC5AC) et sont induits par l'IL-13 chez cellules épithéliales des voies respiratoires humaines[7]. C'est surtout le 15-HETE-PE qui médie les effets de l'IL13 dans des cellules épithéliales pulmonaires humaines[7]. Avec l'augmentation du nombre de cellules caliciformes, il y a une production excessive de mucus dans les bronches[10]. Les conséquences fonctionnelles des changements de stockage et de sécrétion de MUC contribuent aux mécanismes physiopathologiques de diverses anomalies cliniques chez les patients asthmatiques, notamment la production d'expectorations, le rétrécissement des voies respiratoires, l'exacerbation et la perte accélérée de la fonction pulmonaire[10].
De plus, il a été démontré que l'IL-13 induit un puissant programme fibrogène au cours de diverses maladies marquées par des cytokines de type 2 élevées telles que la schistosomiase chronique et la dermatite atopique, entre autres[7]. Lors de l'infection par Schistosoma mansoni, l'IL13 est capable d'induire de manière indépendante à la fois une fibrose hépatique et une prolifération compensatoire de cellules biliaires et d'hépatocytes[7]. Il a été suggéré que ce programme fibrogène dépend de manière critique de la signalisation directe de l'IL-13 par l'IL-4Rα sur les fibroblastes PDGFRβ+[7].
Implications cliniques
L'IL-13 induit des mécanismes de défense de l'organisme par des modifications physiologiques des organes parasités rendant l'environnement local hostile au parasite ou isolant ses produits. Par exemple, la sécrétion d'IL-13 par les cellules Th2 permet l'expulsion de l'intestin d'une variété d' helminthes de souris. L'IL-13 induit plusieurs changements dans les cellules intestinales qui créent un environnement hostile au parasite, notamment en permettant des contractions accrues et une hypersécrétion de glycoprotéines par les cellules épithéliales intestinales, qui conduisent finalement au détachement de l'organisme de la paroi intestinale et à son élimination[12].
Le parasite Schistosoma mansoni peut déposer ses œufs dans les organes tels que l'intestin (surtout dans la paroi intestinale), le foie, les poumons et même le système nerveux central[7]. Le processus normal de translocation de ces œufs dans la paroi intestinale s'accompagne de la formation de granulomes inflammatoires sous le contrôle de l'IL-13 et de la réponse Th2[7]. Le résultat final peut parfois être pathologique avec la production de dommages aux organes et souvent une maladie chronique voire mortelle sans la résolution de l'infection. Un concept émergent est que l'IL-13 peut s'opposer aux réponses Th1 qui sont nécessaires pour résoudre les infections intracellulaires[7]. Dans ce contexte de dérégulation immunitaire, marqué par le recrutement d'un nombre aberrant de cellules Th2, l'IL-13 inhibe la capacité des cellules immunitaires de l'hôte à détruire les pathogènes intracellulaires.
Il a été démontré que l'expression de l'IL-13 est augmentée dans le liquide de lavage broncho-alvéolaire (BAL) et les cellules chez les patients atteints d'asthme léger atopique après provocation allergénique[13]. Le rôle de médiateur central de l'asthme chez l'homme est appuyé par des études d'association à l'échelle du génome qui ont identifié de multiples polymorphismes de l'IL-13 et du récepteur IL4R associés à la susceptibilité à l'asthme, à l'hyperréactivité bronchique et à l'augmentation des taux d'IgE[13]. Malgré son rôle central, vu la diversité des formes d'asthmes, il est probable que d'autres facteurs soient importants[13]. La surexpression de l'IL-13 induit les caractéristiques pathophysiologiques des maladies pulmonaires allergiques, notamment l'hyperréactivité des voies respiratoires, la métaplasie des cellules caliciformes, l'hypersécrétion de mucus et le remodelage des voies respiratoires, qui contribuent toutes à l'obstruction des voies respiratoires[14]. L'IL-13 est capable d'induire ces caractéristiques de manière indépendante des éosinophiles et des immunoglobulines E[14]. Les modèles murins ont permis d'améliorer les connaissances sur les mécanismes d'actions de l'IL-13. Ils ont démontré qu'il était à la fois nécessaire et suffisante pour générer des réponses Th2 de type asthmatique dans le poumon de souris[5]. L'IL-13 est principalement surexprimée dans les expectorations, la sous-muqueuse bronchique, le sang périphérique et les mastocytes du faisceau musculaire lisse des voies respiratoires[5]. L'IL-4 contribue à ces changements physiologiques, mais est moins importante que l'IL-13. L'IL-13 induit également la sécrétion de chimiokines nécessaires au recrutement des cellules effectrices allergiques dans le poumon. Des études sur des souris transgéniques STAT6 suggèrent la possibilité que la signalisation de l'IL-13 se produisant uniquement à travers l'épithélium des voies respiratoires soit nécessaire pour la plupart de ces effets. Bien qu'aucune étude n'ait encore directement impliqué l'IL-13 dans le contrôle des maladies humaines, il a été démontré que de nombreux polymorphismes du gène de l'IL-13 confèrent un risque accru de maladies respiratoires atopiques telles que l' asthme[12]. Dans une étude menée sur des souris knock-out modèle pour l'asthme, la résistance à l'air, la production de mucus et l'induction de médiateurs profibrogéniques se sont révélées uniquement dépendantes de la présence d'IL-13R1 et non d'IL-13Rα2[5]. La surexpression pulmonaire d'IL-13, dans des souris transgéniques, induit une fibrose des voies aériennes sous-épithéliales[5]. Dans des souris transgéniques, l'expression d'IL-13 localisée au niveau pulmonaire était responsable d'une réponse inflammatoire de mononuléaires et d'éosinophiles, une métaplasie de mucus cellulaire, une obstruction des alvéoles et leur fibrose[15]. L'IL-13 est aussi l'effecteur dominant dans les modèles de fibrose par toxine, infection, allergique et post-transplantation de bronchiolite oblitérante[5].
L'interkeukine 13 serait aussi impliquée dans la promotion de la survie et de la migration des cellules épithéliales, de la production d'oxyde nitrique synthase inductible par les cellules épithéliales des voies respiratoires, de l'activation des macrophages, de la perméabilité des cellules épithéliales et de la transformation des fibroblastes des voies respiratoires en myofibroblastes, soit des mécanismes conduisant à un dépôt de collagène et donc impliqués dans la fibrose[13]. Le dépôt de collagène influence alors le remodelage des voies aériennes chez les patients asthmatiques[13].
Le dupilumab est un anticorps monoclonal modulateur de l'IL-13 et de l'IL-4 qui cible le récepteur commun de l'IL-4 et de l'IL-13, l' IL4Rα[16]. Étant donné que l'IL-4 et l'IL-13 ont des activités biologiques similaires, l'antagoniste du récepteur IL4Rα qu'est le dupilumab semble être très prometteur en améliorant la santé des patients asthmatiques à forme non contrôlée lié à une réponse Th2[16].
Voir également
- Récepteur de l'interleukine-13, le récepteur IL-13
Références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Interleukin 13 » (voir la liste des auteurs).
- Les valeurs de la masse et du nombre de résidus indiquées ici sont celles du précurseur protéique issu de la traduction du gène, avant modifications post-traductionnelles, et peuvent différer significativement des valeurs correspondantes pour la protéine fonctionnelle.
- (en) A. Minty, P. Chalon, J.-M. Derocq, X. Dumont, J.-C. Guillemot, M. Kaghad, C. Labit, P. Leplatois, P. Liauzun, B. Miloux, C. Minty, P. Casellas, G. Loison, J. Lupker, D. Shire, P. Ferrara et D. Caput, « Interleukin-13 is a new human lymphokine regulating inflammatory and immune responses », Nature, vol. 362, no 6417, , p. 248–50 (PMID 8096327, DOI 10.1038/362248a0, Bibcode 1993Natur.362..248M)
- (en) A.N. McKenzie, J.A. Culpepper, R. de Waal Malefyt, J. Punnonen, G. Aversa, A. Sato, W. Dang, B.G. Cocks et S. Menon, « Interleukin 13, a T-cell-derived cytokine that regulates human monocyte and B-cell function », Proceedings of the National Academy of Sciences of the United States of America, vol. 90, no 8, , p. 3735–9 (PMID 8097324, PMCID 46376, DOI 10.1073/pnas.90.8.3735, Bibcode 1993PNAS...90.3735M)
- (en) J.G. Morgan, G.M. Dolganov, S.E. Robbins, L.M. Hinton et M. Lovett, « The selective isolation of novel cDNAs encoded by the regions surrounding the human interleukin 4 and 5 genes », Nucleic Acids Res., vol. 20, no 19, , p. 5173–9 (PMID 1408833, PMCID 334302, DOI 10.1093/nar/20.19.5173)
- (en) Efren L. Rael et Richard F. Lockey, « Interleukin-13 signaling and its role in asthma », The World Allergy Organization Journal, vol. 4, no 3, , p. 54–64 (PMID 23283176, PMCID 3651056, DOI 10.1097/WOX.0b013e31821188e0)
- (en) Gerard Zurawski et Jan E. de Vries, « Interleukin 13, an interleukin 4-like cytokine that acts on monocytes and B cells, but not on T cells », Immunol. Today, vol. 15, no 1, , p. 19–26 (PMID 7907877, DOI 10.1016/0167-5699(94)90021-3)
- (en) Rafael Firszt, Dave Francisco, Tony D Church, Joseph M Thomas, Jennifer L Ingram et Monica Kraft, « Interleukin-13 induces collagen type-1 expression through matrix metalloproteinase-2 and transforming growth factor-β1 in airway fibroblasts in asthma », Eur. Respir. J., vol. 43, no 2, , p. 464-473 (PMID 23682108, PMCID PMC6747688, DOI 10.1183/09031936.00068712, lire en ligne)
- (en) Hannah H. Walford et Taylor A. Doherty, « STAT6 and lung inflammation », JAK-STAT, vol. 2, no 4, , e25301 (PMID 24416647, PMCID 3876430, DOI 10.4161/jkst.25301)
- (en) Ming Tu, Wei Wange, Lin Cai, Penglei Zhu, Zhichao Gao et Weiming Zheng, « IL-13 receptor α2 stimulates human glioma cell growth and metastasis through the Src/PI3K/Akt/mTOR signaling pathway », Tumour Biology, vol. 37, no 11, , p. 14701–14709 (PMID 27623944, DOI 10.1007/s13277-016-5346-x)
- (en) John V. Fahy, « Goblet cell and mucin gene abnormalities in asthma », Chest, vol. 122, no 6 Suppl, , p. 320S–326S (PMID 12475809, DOI 10.1378/chest.122.6_suppl.320S, lire en ligne)
- (en) Mitsuko Kondo, Jun Tamaoki, Kiyoshi Takeyama, Junko Nakata et Atsushi Nagai, « Interleukin-13 induces goblet cell differentiation in primary cell culture from Guinea pig tracheal epithelium », Am. J. Respir. Cell Mol. Biol., , p. 536-541 (lire en ligne)
- Narges Seyfizadeh, Nayer Seyfizadeh, Tohid Gharibi et Zohreh Babaloo, « Interleukin-13 as an important cytokine : A review on its roles in some human diseases », Iranian Journal of Allergy, Asthma and Immunology, vol. 62, , p. 341-378 (lire en ligne)
- (en) Jennifer L Ingram et Monica Kraft, « IL-13 in asthma and allergic disease: asthma phenotypes and targeted therapies », The Journal of Allergy and Clinical Immunology, vol. 130, no 4, , p. 829–42; quiz 843–4 (PMID 22951057, DOI 10.1016/j.jaci.2012.06.034, lire en ligne)
- (en) M Wills-Karp, J Luyimbazi, X Xu, B Schofield, T Y Neben, C L Karp et D D Donaldson, « Interleukin-13: central mediator of allergic asthma », Science, vol. 282, no 5397, , p. 2258–61 (PMID 9856949, DOI 10.1126/science.282.5397.2258, Bibcode 1998Sci...282.2258W)
- (en) Z. Zhu, R.J. Homer, Z. Wang, Q. Chen, G. P. Geba, J. Wang, Y Zhang et J.A. Elias, « Pulmonary expression of interleukin-13 causes inflammation, mucus hypersecretion, subepithelial fibrosis, physiologic abnormalities, and eotaxin production », J. Clin. Invest., vol. 103, no 6, , p. 779-788 (PMID 10079098, PMCID PMC408149, DOI 10.1172/JCI5909, lire en ligne)
- (en) Alessandro Vatrella, Immacolata Fabozzi, Cecilia Calabrese, Rosario Maselli et Girolamo Pelaia, « Dupilumab: a novel treatment for asthma », Journal of Asthma and Allergy, vol. 7, , p. 123–30 (PMID 25214796, PMCID 4159398, DOI 10.2147/JAA.S52387, lire en ligne)
Liens externes
- Overview of all the structural information available in the PDB for UniProt: P35225 (Interleukin-13) at the PDBe-KB.
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire